
Attenuation of a Virulent Porcine Deltacoronavirus Strain DHeB1 via Serial Passage in LLC-PK1 Cells

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cell Culture and Virus
2.3. Immunofluorescence Assay (IFA)
2.4. Growth Kinetics
2.5. Complete Genome Sequencing and Phylogenetic Analysis
2.6. Pathogenicity of PDCoV DHeB1 Serially Passaged Strains in 5-Day-Old Piglets
2.7. Histopathology and Immunohistochemistry
2.8. Immunogenicity of PDCoV DHeB1 Serially Passaged Strains in Weaned Piglets
2.9. Serum Neutralization Test
2.10. Statistical Analysis
3. Results
3.1. Biological Characteristics of PDCoV DHeB1 Serially Passaged Strains
3.2. Complete Genome Sequencing and Phylogenetic Analysis of PDCoV DHeB1 Serially Passaged Strains
3.3. Clinical Evaluation of Piglets Challenged with PDCoV DHeB1 Serially Passaged Strains
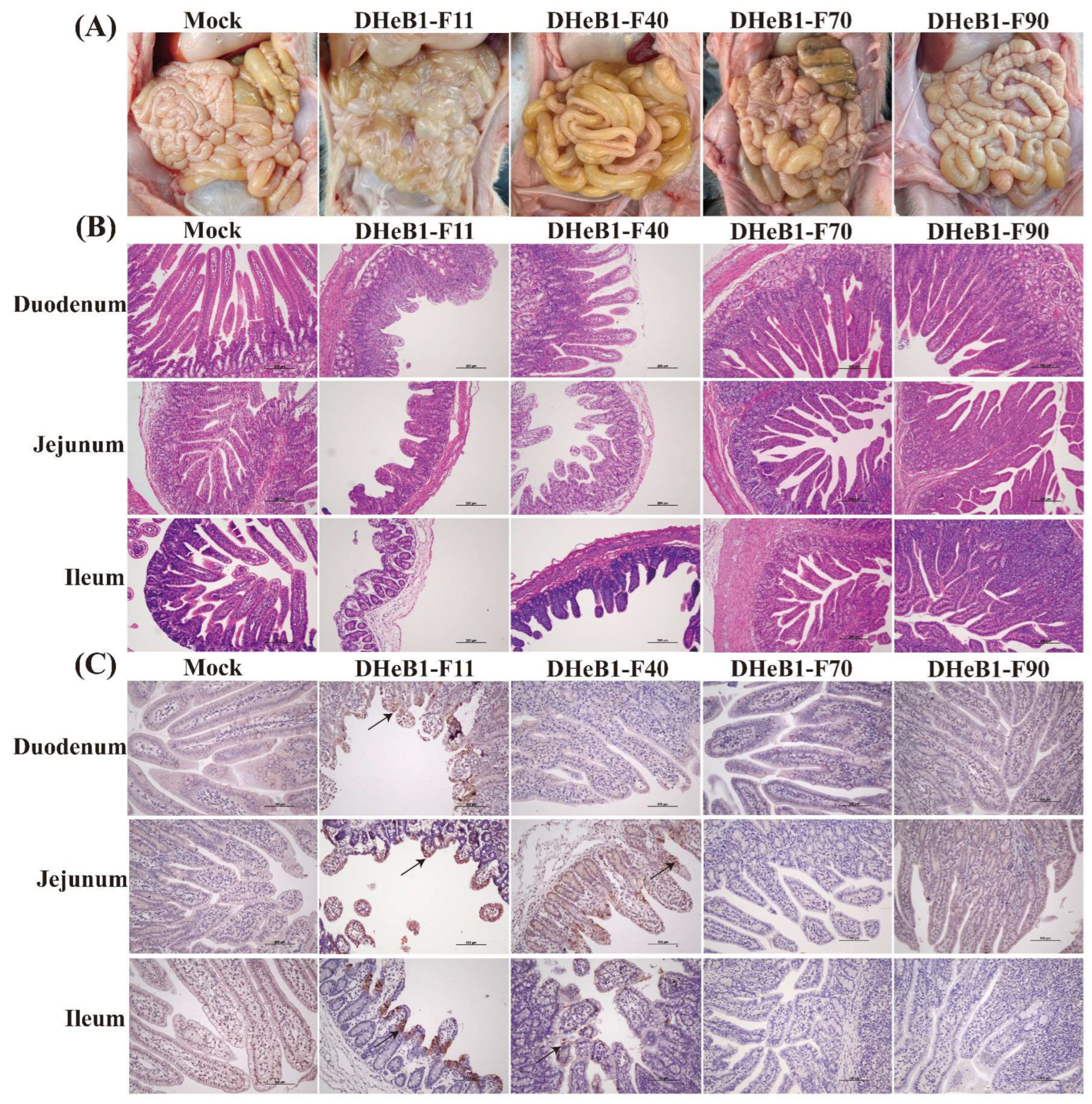
3.4. Histopathological Observations
3.5. Immunogenicity Evaluation of PDCoV DHeB1 Serially Passaged Strains in Weaned Piglets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, Y.M.; Zhang, Y.; Liang, X.Y.; Lou, F.F.; Oglesbee, M.; Krakowka, S.; Li, J.R. Origin, Evolution, and Virulence of Porcine Deltacoronaviruses in the United States. mBio 2015, 6, e00064. [Google Scholar] [CrossRef] [PubMed]
- Fang, P.X.; Fang, L.R.; Hong, Y.Y.; Liu, X.R.; Dong, N.; Ma, P.P.; Bi, J.; Wang, D.; Xiao, S.B. Discovery of a novel accessory protein NS7a encoded by porcine deltacoronavirus. J. Gen. Virol. 2017, 98, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.C.Y.; Huang, Y.; Lau, S.K.P.; Yuen, K.Y. Coronavirus Genomics and Bioinformatics Analysis. Viruses 2010, 2, 1804–1820. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Gauger, P.; Stafne, M.; Thomas, J.; Arruda, P.; Burrough, E.; Madson, D.; Brodie, J.; Magstadt, D.; Derscheid, R.; et al. Pathogenicity and pathogenesis of a United States porcine deltacoronavirus cell culture isolate in 5-day-old neonatal piglets. Virology 2015, 482, 51–59. [Google Scholar] [CrossRef]
- Jung, K.; Hu, H.; Eyerly, B.; Lu, Z.Y.; Chepngeno, J.; Saif, L.J. Pathogenicity of 2 Porcine Deltacoronavirus Strains in Gnotobiotic Pigs. Emerg. Infect. Dis. 2015, 21, 650–654. [Google Scholar] [CrossRef]
- Zhao, Y.J.; Qu, H.; Hu, J.F.; Fu, J.Y.; Chen, R.; Li, C.; Cao, S.J.; Wen, Y.P.; Wu, R.; Zhao, Q.; et al. Characterization and Pathogenicity of the Porcine Deltacoronavirus Isolated in Southwest China. Viruses 2019, 11, 1074. [Google Scholar] [CrossRef]
- Zhang, H.L.; Han, F.F.; Shu, X.L.; Li, Q.Q.; Ding, Q.W.; Hao, C.L.; Yan, X.G.; Xu, M.L.; Hu, H. Co-infection of porcine epidemic diarrhoea virus and porcine deltacoronavirus enhances the disease severity in piglets. Transbound. Emerg. Dis. 2021, 69, 1715–1726. [Google Scholar] [CrossRef]
- Duan, C. An Updated Review of Porcine Deltacoronavirus in Terms of Prevalence, Pathogenicity, Pathogenesis and Antiviral Strategy. Front. Vet. Sci. 2022, 8, 811187. [Google Scholar] [CrossRef]
- Lednicky, J.A.; Tagliamonte, M.S.; White, S.K.; Elbadry, M.A.; Alam, M.M.; Stephenson, C.J.; Bonny, T.S.; Loeb, J.C.; Telisma, T.; Chavannes, S.; et al. Independent infections of porcine deltacoronavirus among Haitian children. Nature 2021, 600, 133–137. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Lau, S.K.P.; Lam, C.S.F.; Lau, C.C.Y.; Tsang, A.K.L.; Lau, J.H.N.; Bai, R.; Teng, J.L.L.; Tsang, C.C.C.; Wang, M.; et al. Discovery of seven novel Mammalian and avian coronaviruses in the genus deltacoronavirus supports bat coronaviruses as the gene source of alphacoronavirus and betacoronavirus and avian coronaviruses as the gene source of gammacoronavirus and deltacoronavirus. J. Virol. 2012, 86, 3995–4008. [Google Scholar] [CrossRef]
- Zhang, F.F.; Luo, S.X.; Gu, J.; Li, Z.Q.; Li, K.; Yuan, W.F.; Ye, Y.; Li, H.; Ding, Z.; Song, D.P.; et al. Prevalence and phylogenetic analysis of porcine diarrhea associated viruses in southern China from 2012 to 2018. BMC Vet. Res. 2019, 15, 470. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Shibahara, T.; Imai, N.; Yamamoto, T.; Ohashi, S. Genetic characterization and pathogenicity of Japanese porcine deltacoronavirus. Infect. Genet. Evol. 2018, 61, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Ajayi, T.; Dara, R.; Misener, M.; Pasma, T.; Moser, L.; Poljak, Z. Herd-level prevalence and incidence of porcine epidemic diarrhoea virus (PEDV) and porcine deltacoronavirus (PDCoV) in swine herds in Ontario, Canada. Transbound. Emerg. Dis. 2018, 65, 1197–1207. [Google Scholar] [CrossRef] [PubMed]
- Saeng-chuto, K.; Stott, C.J.; Wegner, M.; Senasuthum, R.; Tantituvanont, A.; Nilubol, D. Retrospective investigation and evolutionary analysis of a novel porcine deltacoronavirus strain detected in Thailand from 2008 to 2015. Arch. Virol. 2017, 162, 2103–2108. [Google Scholar] [CrossRef]
- Lee, S.; Lee, C. Complete Genome Characterization of Korean Porcine Deltacoronavirus Strain KOR/KNU14-04/2014. Genome Announc. 2014, 2, e01191-14. [Google Scholar] [CrossRef]
- Turlewicz-Podbielska, H.; Pomorska-Mól, M. Porcine Coronaviruses: Overview of the State of the Art. Virol. Sin. 2021, 36, 833–851. [Google Scholar] [CrossRef]
- Mai, K.J.; Li, D.; Wu, J.L.; Wu, Z.X.; Cheng, J.; He, L.L.; Tang, X.Y.; Zhou, Z.H.; Sun, Y.; Ma, J.Y. Complete Genome Sequences of Two Porcine Deltacoronavirus Strains, CHN-GD16-03 and CHN-GD16-05, Isolated in Southern China, 2016. Genome Announc. 2018, 6, e01545-17. [Google Scholar] [CrossRef]
- He, W.T.; Ji, X.; He, W.; Dellicour, S.; Wang, S.L.; Li, G.R.; Zhang, L.T.; Gilbert, M.; Zhu, H.N.; Xing, G.; et al. Genomic Epidemiology, Evolution, and Transmission Dynamics of Porcine Deltacoronavirus. Mol. Biol. Evol. 2020, 37, 2641–2654. [Google Scholar] [CrossRef]
- Dong, N.; Fang, L.R.; Zeng, S.L.; Sun, Q.Q.; Chen, H.C.; Xiao, S.B. Porcine Deltacoronavirus in Mainland China. Emerg. Infect. Dis. 2015, 21, 2254–2255. [Google Scholar] [CrossRef]
- Kong, F.Z.; Wang, Q.H.; Kenney, S.P.; Jung, K.N.; Vlasova, A.N.; Saif, L.J. Porcine Deltacoronaviruses: Origin, Evolution, Cross-Species Transmission and Zoonotic Potential. Pathogens 2022, 11, 79. [Google Scholar] [CrossRef]
- Wang, H.J.; Qin, Y.B.; Zhao, W.; Yuan, T.T.; Yang, C.J.; Mi, X.; Zhao, P.; Lu, Y.; Lu, B.X.; Chen, Z.W.; et al. Genetic Characteristics and Pathogenicity of a Novel Porcine Deltacoronavirus Southeast Asia-Like Strain Found in China. Front. Vet. Sci. 2021, 8, 701612. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.X.; Yin, Y.W.; Wang, W.; Cao, L.; Sun, W.C.; Shi, K.C.; Lu, H.J.; Jin, N.Y. Emergence of Thailand-like strains of porcine deltacoronavirus in Guangxi Province, China. Vet. Med. Sci. 2020, 6, 854–859. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.L.; Chen, J.F.; Liu, Y.; Da, S.; Shi, H.Y.; Zhang, X.; Liu, J.B.; Cao, L.Y.; Zhu, X.D.; Wang, X.B.; et al. Pathogenicity of porcine deltacoronavirus (PDCoV) strain NH and immunization of pregnant sows with an inactivated PDCoV vaccine protects 5-day-old neonatal piglets from virulent challenge. Transbound. Emerg. Dis. 2019, 67, 572–583. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Z.; Zhao, S.Q.; Zhang, B.T.; Huang, J.; Peng, Q.; Xiao, L.; Yuan, X.S.; Guo, R.L.; Zhou, J.Z.; Fan, B.C.; et al. A novel recombinant S-based subunit vaccine induces protective immunity against porcine deltacoronavirus challenge in piglets. J. Virol. 2023, 97, e0095823. [Google Scholar] [CrossRef]
- Huang, Y.; Xu, Z.W.; Gu, S.R.; Nie, M.C.; Wang, Y.L.; Zhao, J.; Li, F.Q.; Deng, H.D.; Huang, J.B.; Sun, X.G.; et al. The recombinant pseudorabies virus expressing porcine deltacoronavirus spike protein is safe and effective for mice. BMC Vet. Res. 2022, 18, 16. [Google Scholar] [CrossRef]
- Chen, P.F.; Zhao, X.W.; Zhou, S.T.; Zhou, T.X.; Tan, X.M.; Wu, X.; Tong, W.; Gao, F.; Yu, L.X.; Jiang, Y.F.; et al. A Virulent PEDV Strain FJzz1 with Genomic Mutations and Deletions at the High Passage Level Was Attenuated in Piglets via Serial Passage In Vitro. Virol. Sin. 2021, 36, 1052–1065. [Google Scholar] [CrossRef]
- Chen, Y.; He, S.Y.; Sun, L.; Luo, Y.F.; Sun, Y.K.; Xie, J.X.; Zhou, P.; Su, S.; Zhang, G.H. Genetic variation, pathogenicity, and immunogenicity of highly pathogenic porcine reproductive and respiratory syndrome virus strain XH-GD at different passage levels. Arch. Virol. 2016, 161, 77–86. [Google Scholar] [CrossRef]
- Li, X.F.; Cui, Z.; Fan, H.; Chen, Q.; Cao, L.; Qiu, H.Y.; Zhang, N.N.; Xu, Y.P.; Zhang, R.R.; Zhou, C.; et al. A highly immunogenic live-attenuated vaccine candidate prevents SARS-CoV-2 infection and transmission in hamsters. Innovation 2022, 3, 100221. [Google Scholar] [CrossRef]
- Dong, N.; Fang, L.R.; Yang, H.; Liu, H.; Du, T.; Fang, P.X.; Wang, D.; Chen, H.C.; Xiao, S.B. Isolation, genomic characterization, and pathogenicity of a Chinese porcine deltacoronavirus strain CHN-HN-2014. Vet. Microbiol. 2016, 196, 98–106. [Google Scholar] [CrossRef]
- Luo, J.Y.; Fang, L.R.; Dong, N.; Fang, P.X.; Ding, Z.; Wang, D.; Chen, H.C.; Xiao, S.B. Porcine deltacoronavirus (PDCoV) infection suppresses RIG-I-mediated interferon-β production. Virology 2016, 495, 10–17. [Google Scholar] [CrossRef]
- Lee, J.H.; Chung, H.C.; Nguyen, V.G.; Moon, H.J.; Kim, H.K.; Park, S.J.; Lee, C.H.; Lee, G.E.; Park, B.K. Detection and Phylogenetic Analysis of Porcine Deltacoronavirus in Korean Swine Farms, 2015. Transbound. Emerg. Dis. 2016, 63, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Le, V.P.; Song, S.; An, B.H.; Park, G.N.; Pham, N.T.; Le, D.Q.; Nguyen, V.T.; Vu, T.T.H.; Kim, K.S.; Choe, S.; et al. A novel strain of porcine deltacoronavirus in Vietnam. Arch. Virol. 2018, 163, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.Q.; Zhang, H.L.; Li, B.X.; Ding, Q.W.; Wang, Y.B.; Gao, W.M.; Guo, D.H.; Wei, Z.Y.; Hu, H. Susceptibility of Chickens to Porcine Deltacoronavirus Infection. Viruses 2019, 11, 573. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.L.; Ding, Q.W.; Yuan, J.; Han, F.F.; Wei, Z.Y.; Hu, H. Susceptibility to mice and potential evolutionary characteristics of porcine deltacoronavirus. J. Med. Virol. 2022, 94, 5723–5738. [Google Scholar] [CrossRef]
- Yang, J.P.; Chen, R.; Sun, M.K.; Yuan, R.; Xiao, Y.F.; Sun, Y.; Zhou, G.P.; Wen, Y.P.; Wang, Y.P.; Wu, R.; et al. Development and immunogenicity evaluation of attenuated Salmonella typhimurium delivering porcine Deltacoronavirus S1 gene. Int. J. Biol. Macromol. 2025, 288, 138615. [Google Scholar] [CrossRef]
- Li, J.Z.; Xiao, L.; Chen, Z.Q.; Fan, L.Y.; Wang, W.; Guo, R.L.; He, Z.M.; Hu, H.P.; Jiang, J.H.; Zhao, L.X.; et al. A spike-based mRNA vaccine that induces durable and broad protection against porcine deltacoronavirus in piglets. J. Virol. 2024, 98, e0053524. [Google Scholar] [CrossRef]
- Woods, R.D. Efficacy of a transmissible gastroenteritis coronavirus with an altered ORF-3 gene. Can. J. Vet. Res. 2001, 65, 28–32. [Google Scholar]
- Song, D.S.; Oh, J.S.; Kang, B.K.; Yang, J.S.; Moon, H.J.; Yoo, H.S.; Jang, Y.S.; Park, B.K. Oral efficacy of Vero cell attenuated porcine epidemic diarrhea virus DR13 strain. Res. Vet. Sci. 2007, 82, 134–140. [Google Scholar] [CrossRef]
- Kweon, C.H.; Kwon, B.J.; Lee, J.G.; Kwon, G.O.; Kang, Y.B. Derivation of attenuated porcine epidemic diarrhea virus (PEDV) as vaccine candidate. Vaccine 1999, 17, 2546–2553. [Google Scholar] [CrossRef]
- Liu, Q.G. Genomic Characteristics and Pathogenicity Analysis of Different Passages of Porcine Deltacoronavirus NH Strain. Master’s Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2020. [Google Scholar]
- Xiong, X.L.; Tortorici, M.A.; Snijder, J.; Yoshioka, C.; Walls, A.C.; Li, W.T.; McGuire, A.T.; Rey, F.A.; Bosch, B.J.; Veesler, D. Glycan Shield and Fusion Activation of a Deltacoronavirus Spike Glycoprotein Fine-Tuned for Enteric Infections. J. Virol. 2018, 92, e01628-17. [Google Scholar] [CrossRef]
- Shang, J.; Zheng, Y.; Yang, Y.; Liu, C.; Geng, Q.B.; Tai, W.B.; Du, L.Y.; Zhou, Y.S.; Zhang, W.; Li, F. Cryo-Electron Microscopy Structure of Porcine Deltacoronavirus Spike Protein in the Prefusion State. J. Virol. 2018, 92, e01556-17. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Fu, J.Y.; Hu, J.F.; Li, C.; Zhao, Y.J.; Qu, H.; Wen, X.T.; Cao, S.J.; Wen, Y.P.; Wu, R.; et al. Identification of the immunodominant neutralizing regions in the spike glycoprotein of porcine deltacoronavirus. Virus Res. 2020, 276, 197834. [Google Scholar] [CrossRef] [PubMed]
- DeDiego, M.L.; Nieto-Torres, J.L.; Jimenez-Guardeño, J.M.; Regla-Nava, J.A.; Castaño-Rodriguez, C.; Fernandez-Delgado, R.; Usera, F.; Enjuanes, L. Coronavirus virulence genes with main focus on SARS-CoV envelope gene. Virus Res. 2014, 194, 124–137. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Jung, K.; Vlasova, A.N.; Saif, L.J. Experimental infection of gnotobiotic pigs with the cell-culture-adapted porcine deltacoronavirus strain OH-FD22. Arch. Virol. 2016, 161, 3421–3434. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Amino Acid Position | PDCoV-DHeB1 Passage | ||||
|---|---|---|---|---|---|---|
| F11 | F40 | F70 | F90 | F110 | ||
| Nsp2 | 19 | CTA (L) | CCA (P) * | CCA (P) | CCA (P) | CCA (P) |
| Nsp3 | 831 | GGG (G) | GGG (G) | TGG (W) | TGG (W) | TGG (W) |
| Nsp4 | 2129 | AGG (R) | AGG (R) | ATG (M) | ATG (M) | ATG (M) |
| Nsp4 | 2190 | CCT (P) | TCT (S) | CCT (P) | CCT (P) | CCT (P) |
| Nsp4 | 2486 | GGT (G) | GGT (G) | GGT (G) | AGT (S) | GGT (G) |
| Nsp5 | 2555 | GAC (D) | GAC (D) | GAC (D) | CAC (H) | CAC (H) |
| Nsp9 | 3452 | GGA (G) | AGA (R) | AGA (R) | GGA (G) | GGA (G) |
| Nsp12 | 163 | CCA (P) | CCA (P) | CCA (P) | CCA (P) | CAA (Q) |
| S | 29 | CCG (P) | CCG (P) | TCG (S) | TCG (S) | TCG (S) |
| 140 | ATC (I) | ATC (I) | ATC (I) | ATC (I) | ATG (M) | |
| 151 | AAT (N) | AAT (N) | AAT (N) | AAT (N) | AAG (K) | |
| 160 | TTT (F) | TCT (S) | TCT (S) | TCT (S) | TCT (S) | |
| 162 | GAC (D) | TAC (Y) | TAC (Y) | TAC (Y) | TAC (Y) | |
| 169 | TCT (S) | TCT (S) | CCT (P) | CCT (P) | CCT (P) | |
| 396 | AAT (N) | AAG (K) | AAG (K) | AAG (K) | AAG (K) | |
| 488 | CTA (L) | CAA (Q) | CAA (Q) | CAA (Q) | CAA (Q) | |
| 552 | ACA (T) | ATA (I) | ATA (I) | ACA (T) | ACA (T) | |
| 641 | AAA (K) | AAA (K) | AAA (K) | AAA (K) | CAA (Q) | |
| 705 | GCT (A) | ACT (T) | ACT (T) | ACT (T) | ACT (T) | |
| 926 | AAT (N) | AAT (N) | AAT (N) | TAT (Y) | AAT (N) | |
| 946 | TCT (S) | TTT (F) | TTT (F) | TTT (F) | TTT (F) | |
| 947 | GCC (A) | GCC (A) | GCC (A) | GCC (A) | ACC (T) | |
| 1067 | GCA (A) | GTA (V) | GTA (V) | GTA (V) | GTA (V) | |
| 1094 | CTT (L) | CTT (L) | ATT (I) | ATT (I) | ATT (I) | |
| E | 19 | CTA (L) | CCA (P) | CTA (L) | CTA (L) | CTA (L) |
| 61 | AAG (K) | AAG (K) | GAG (E) | GAG (E) | GAG (E) | |
| N | 101 | CCG (P) | CCG (P) | TCG (S) | TCG (S) | TCG (S) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Liu, K.; Chen, L.; Yuan, M.; Lu, H.; Xiao, S.; Fang, L. Attenuation of a Virulent Porcine Deltacoronavirus Strain DHeB1 via Serial Passage in LLC-PK1 Cells. Viruses 2025, 17, 695. https://doi.org/10.3390/v17050695
Zhang Y, Liu K, Chen L, Yuan M, Lu H, Xiao S, Fang L. Attenuation of a Virulent Porcine Deltacoronavirus Strain DHeB1 via Serial Passage in LLC-PK1 Cells. Viruses. 2025; 17(5):695. https://doi.org/10.3390/v17050695
Chicago/Turabian StyleZhang, Yuhan, Kang Liu, Longfei Chen, Meng Yuan, Hongyu Lu, Shaobo Xiao, and Liurong Fang. 2025. "Attenuation of a Virulent Porcine Deltacoronavirus Strain DHeB1 via Serial Passage in LLC-PK1 Cells" Viruses 17, no. 5: 695. https://doi.org/10.3390/v17050695
APA StyleZhang, Y., Liu, K., Chen, L., Yuan, M., Lu, H., Xiao, S., & Fang, L. (2025). Attenuation of a Virulent Porcine Deltacoronavirus Strain DHeB1 via Serial Passage in LLC-PK1 Cells. Viruses, 17(5), 695. https://doi.org/10.3390/v17050695