

Protective Efficacy of a Chimeric Pestivirus KD26_E2LOM Vaccine Against Classical Swine Fever Virus Infection of Pigs

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Efficacy of the KD26_E2 LOM Strain in Animals
2.2. Real-Time PCR (qRT-PCR) to Detect CSFV and BVDV
2.3. Anti-CSFV E2 and Anti-CSFV Erns Antibodies
2.4. Serum Neutralization Assay
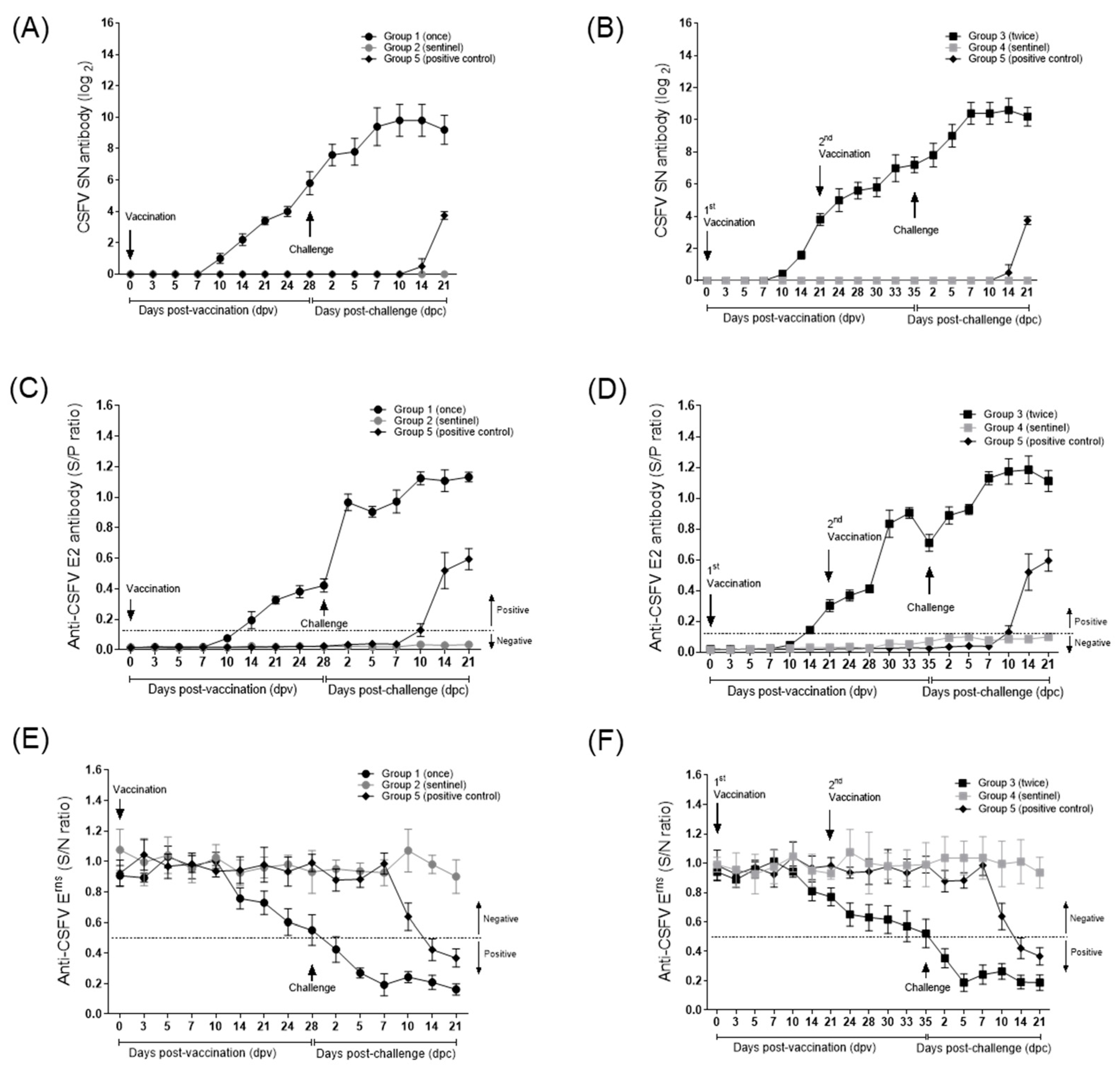
3. Results
3.1. Body Temperature, Leukocyte Counts, and CSFV Antigen Levels in Challenged Pigs
3.2. Anti-CSFV and Anti-BVDV Antibody Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yadav, S.; Weng, H.Y. Estimating the Scale of Adverse Animal Welfare Consequences of Movement Restriction and Mitigation Strategies in a Classical Swine Fever Outbreak. BMC Vet. Res. 2017, 13, 83. [Google Scholar] [CrossRef]
- Ganges, L.; Crooke, H.R.; Bohórquez, J.A.; Postel, A.; Sakoda, Y.; Becher, P.; Ruggli, N. Classical Swine Fever Virus: The Past, Present and Future. Virus Res. 2020, 289, 198151. [Google Scholar] [CrossRef]
- Meyers, G.; Thiel, H.J. Molecular Characterization of Pestiviruses. In Advances in Virus Research; Maramorosch, K., Murphy, F.A., Shatkin, A.J., Eds.; Academic Press: San Diego, CA, USA, 1996; Volume 47, pp. 53–118. [Google Scholar]
- Tautz, N.; Tews, B.A.; Meyers, G. Chapter Two—The Molecular Biology of Pestiviruses. In Advances in Virus Research; Kielian, M., Maramorosch, K., Mettenleiter, T.C., Eds.; Academic Press: San Diego, CA, USA, 2015; Volume 93, pp. 47–160. [Google Scholar]
- Postel, A.; Austermann-Busch, S.; Petrov, A.; Moennig, V.; Becher, P. Epidemiology, Diagnosis and Control of Classical Swine Fever: Recent Developments and Future Challenges. Transbound. Emerg. Dis. 2018, 65, 248–261. [Google Scholar] [CrossRef]
- Kaashoek, M.J.; Moerman, A.; Madic, J.; Rijsewijk, F.A.; Quak, J.; Gielkens, A.L.; van Oirschot, J.T. A conventionally attenuated glycoprotein E-negative strain of bovine herpesvirus type 1 is an efficacious and safe vaccine. Vaccine 1994, 12, 439–444. [Google Scholar] [CrossRef]
- van Oirschot, J.T.; Gielkens, A.L.; Moormann, R.J.; Berns, A.J. Marker vaccines, virus protein-specific antibody assays and the control of Aujeszky’s disease. Vet. Microbiol. 1990, 23, 85–101. [Google Scholar] [CrossRef]
- Reimann, I.; Depner, K.; Trapp, S.; Beer, M. An avirulent chimeric Pestivirus with altered cell tropism protects pigs against lethal infection with classical swine fever virus. Virology 2004, 322, 143–157. [Google Scholar] [CrossRef]
- Wei, Q.; Liu, Y.; Zhang, G. Research Progress and Challenges in Vaccine Development against Classical Swine Fever Virus. Viruses 2021, 13, 445. [Google Scholar] [CrossRef]
- Postel, A.; Becher, P. Genetically Distinct Pestiviruses Pave the Way to Improved Classical Swine Fever Marker Vaccine Candidates Based on the Chimeric Pestivirus Concept. Emerg. Microbes Infect. 2020, 9, 2180–2189. [Google Scholar] [CrossRef]
- Huynh, L.T.; Isoda, N.; Hew, L.Y.; Ogino, S.; Mimura, Y.; Kobayashi, M.; Kim, T.; Nishi, T.; Fukai, K.; Hiono, T.; et al. Generation and Efficacy of Two Chimeric Viruses Derived from GPE− Vaccine Strain as Classical Swine Fever Vaccine Candidates. Viruses 2023, 15, 1587. [Google Scholar] [CrossRef]
- Huynh, L.T.; Otsuka, M.; Kobayashi, M.; Ngo, H.D.; Hew, L.Y.; Hiono, T.; Isoda, N.; Sakoda, Y. Assessment of the Safety Profile of Chimeric Marker Vaccine against Classical Swine Fever: Reversion to Virulence Study. Viruses 2024, 16, 1120. [Google Scholar] [CrossRef]
- Park, G.N.; Shin, J.; Choe, S.; Kim, K.S.; Kim, J.J.; Lim, S.I.; An, B.H.; Hyun, B.H.; An, D.J. Safety and Immunogenicity of Chimeric Pestivirus KD26_E2LOM in Piglets and Calves. Vaccines 2023, 11, 1622. [Google Scholar] [CrossRef]
- Park, G.S.; Lim, S.I.; Hong, S.H.; Song, J.Y. Establishment and characterization of an infectious cDNA clone of a classical swine fever virus LOM strain. J. Vet. Sci. 2012, 13, 81–91. [Google Scholar]
- Lim, S.I.; Choe, S.; Kim, K.S.; Jeoung, H.Y.; Cha, R.M.; Park, G.S.; Shin, J.; Park, G.N.; Cho, I.S.; Song, J.Y.; et al. Assessment of the efficacy of an attenuated live marker classical swine fever vaccine (Flc-LOM-BE(rns)) in pregnant sows. Vaccine 2019, 37, 3598–3604. [Google Scholar] [CrossRef]
- Meyers, T.; Becher, T. Recovery of cytopathogenic and noncytopathogenic bovine viral diarrhea viruses from cDNA constructs. J. Virol. 1996, 70, 8606–8613. [Google Scholar]
- Moormann, R.J.; van Gennip, H.G.; Miedema, G.K.; Hulst, M.M.; van Rijn, P.A. Infectious RNA transcribed from an engineered full-length cDNA template of the genome of a pestivirus. J. Virol. 1996, 70, 763–770. [Google Scholar]
- Ruggli, N.; Tratschin, J.D.; Mittelholzer, C.; Hofmann, M.A. Nucleotide sequence of classical swine fever virus strain Alfort/187 and transcription of infectious RNA from stably cloned full-length cDNA. J. Virol. 1996, 70, 3478–3487. [Google Scholar]
- Vassilev, V.B.; Collett, M.S.; Donis, R.O. Authentic and chimeric full-length genomic cDNA clones of bovine viral diarrhea virus that yield infectious transcripts. J. Virol. 1997, 71, 471. [Google Scholar] [CrossRef]
- Corapi, W.V.; Donis, R.O.; Dubovi, E.J. Monoclonal antibody analyses of cytopathic and noncytopathic viruses from fatal bovine viral diarrhea virus infections. J. Virol. 1988, 62, 2823–2827. [Google Scholar]
- Koenig, P.; Lange, E.; Reimann, I.; Beer, M. CP7_E2alf: A safe and efficient marker vaccine strain for oral immunisation of wild boar against classical swine fever virus (CSFV). Vaccine 2007, 25, 3391–3399. [Google Scholar]
- Renson, P.; LeDimna, M.; Keranflech, A.; Cariolet, R.; Koenen, F.; Le Potier, M.F. CP7_E2alf oral vaccination confers partial protection against early classical swine fever virus challenge and interferes with pathogeny-related cytokine responses. Vet. Res. 2013, 44, 9. [Google Scholar] [CrossRef]
- Blome, S.; Wernike, K.; Reimann, I.; Konig, P.; Moss, C.; Beer, M. A decade of research into classical swine fever marker vaccine CP7_E2alf (Suvaxyn® CSF Marker): A review of vaccine properties. Vet. Res. 2017, 48, 51. [Google Scholar] [PubMed]
- Meyer, D.; Fritsche, S.; Luo, Y.; Engemann, C.; Blome, S.; Beyerbach, M.; Chang, C.Y.; Qiu, H.J.; Becher, P.; Postel, A. The double-antigen ELISA concept for early detection of Erns-specific classical swine fever virus antibodies and application as an accompanying test for differentiation of infected from marker vaccinated animals. Transbound. Emerg. Dis. 2017, 64, 2013–2022. [Google Scholar] [PubMed]
- Choe, S.; Shin, J.; Kim, K.S.; Song, S.; Cha, R.M.; Jung, B.I.; Hyun, B.H.; Park, B.K.; An, D.J. Protection of Piglets with Maternally Derived Antibodies from Sows Inoculated with an Attenuated Live Marker Classical Swine Fever Vaccine (Flc-LOM-BErns). Pathogens 2020, 9, 608. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Group | Vaccine (no. of Injections) | No. Pigs | Days after CSFV Inoculation | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 2 | 5 | 7 | 10 | 14 | 21 | |||
| 1 | KD26_E2 LOM strain (one) | 5 | 0/0/0 | 0/0/0 | 0/0/0 | 0/0/0 | 0/0/0 | 0/0/0 | 0/0/0 |
| 2 | Sentinel with group 1 | 2 | 0/0/0 | 0/0/0 | 0/0/0 | 0/0/0 | 0/0/0 | 0/0/0 | 0/0/0 |
| 3 | KD26_E2 LOM strain (two) | 5 | 0/0/0 | 0/0/0 | 0/0/0 | 0/0/0 | 0/0/0 | 0/0/0 | 0/0/0 |
| 4 | Sentinel with group 3 | 2 | 0/0/0 | 0/0/0 | 0/0/0 | 0/0/0 | 0/0/0 | 0/0/0 | 0/0/0 |
| 5 | Positive control | 4 | 0/0/0 | 0/0/0 | 4/0/4 * | 4/4/4 | 4/4/4 | 4/4/4 | 4/4/4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.-H.; Jung, B.-K.; Kim, S.-Y.; Kim, D.; Jang, M.-K.; Choe, S.; An, B.-H.; Kim, J.-J.; Cho, Y.S.; An, D.-J. Protective Efficacy of a Chimeric Pestivirus KD26_E2LOM Vaccine Against Classical Swine Fever Virus Infection of Pigs. Viruses 2025, 17, 529. https://doi.org/10.3390/v17040529
Lee Y-H, Jung B-K, Kim S-Y, Kim D, Jang M-K, Choe S, An B-H, Kim J-J, Cho YS, An D-J. Protective Efficacy of a Chimeric Pestivirus KD26_E2LOM Vaccine Against Classical Swine Fever Virus Infection of Pigs. Viruses. 2025; 17(4):529. https://doi.org/10.3390/v17040529
Chicago/Turabian StyleLee, Young-Hyeon, Bo-Kyoung Jung, Song-Yi Kim, Dohyun Kim, Min-Kyung Jang, SeEun Choe, Byung-Hyun An, Jae-Jo Kim, Yun Sang Cho, and Dong-Jun An. 2025. "Protective Efficacy of a Chimeric Pestivirus KD26_E2LOM Vaccine Against Classical Swine Fever Virus Infection of Pigs" Viruses 17, no. 4: 529. https://doi.org/10.3390/v17040529
APA StyleLee, Y.-H., Jung, B.-K., Kim, S.-Y., Kim, D., Jang, M.-K., Choe, S., An, B.-H., Kim, J.-J., Cho, Y. S., & An, D.-J. (2025). Protective Efficacy of a Chimeric Pestivirus KD26_E2LOM Vaccine Against Classical Swine Fever Virus Infection of Pigs. Viruses, 17(4), 529. https://doi.org/10.3390/v17040529