
Impact of BTV-3 Circulation in Belgium in 2024 and Current Knowledge Gaps Hindering an Evidence-Based Control Program

 , ,
, ,  , , ,
, , ,  , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Confirmatory BTV-3 Diagnostics in 2024
2.2. Additional Samples for Virological and Serological BTV Testing
2.3. RNA Extraction and RT-qPCRs
2.4. BTV ELISA
2.5. BTV VNT
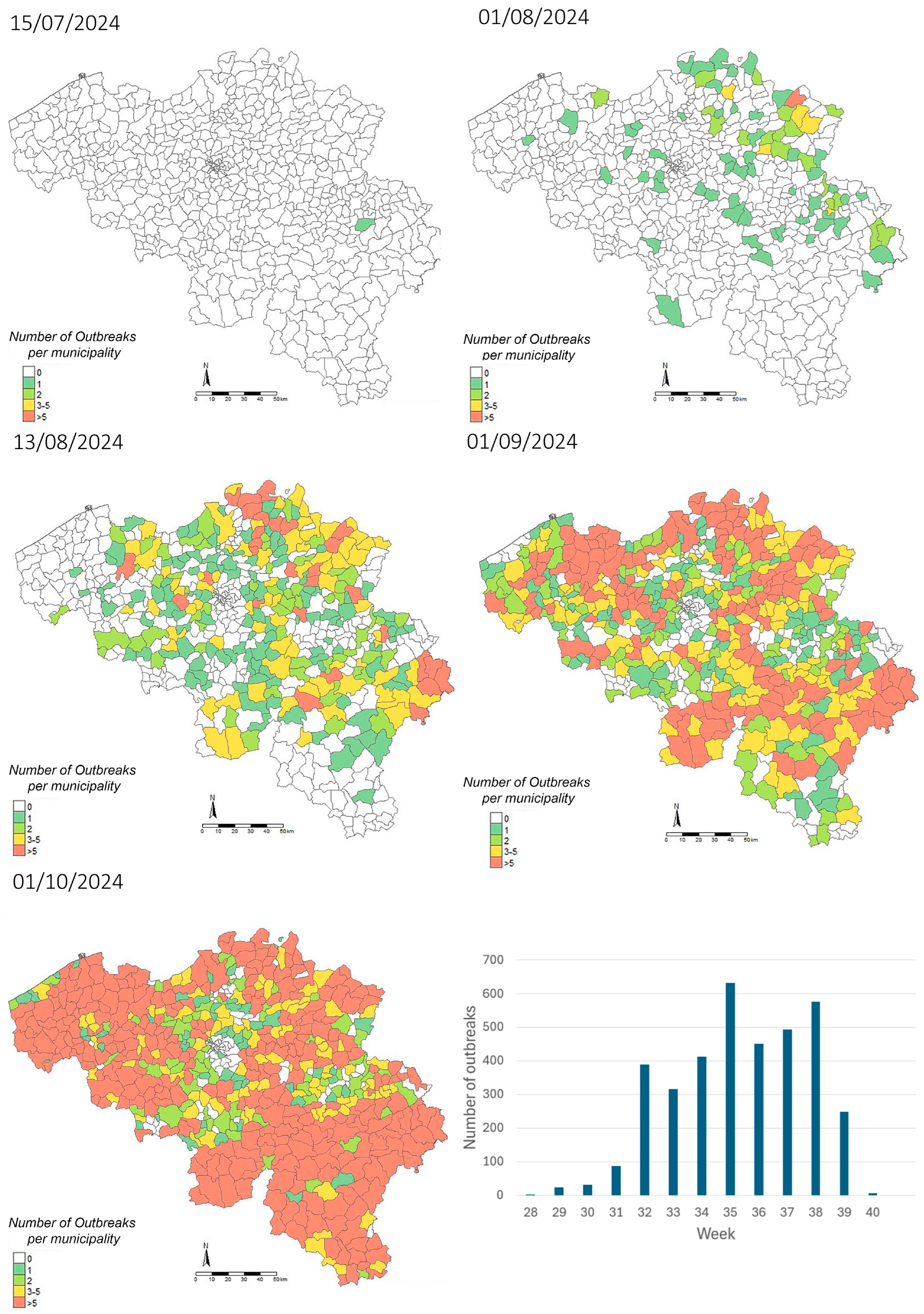
2.6. Geographical Mapping of Confirmed BTV Cases
2.7. Data on Ruminant Mortality, Births and Milk Production
2.8. Calculation of Mortality Rates
2.9. Statistical Analysis
3. Results and Discussion
3.1. BTV-3 Spread in Belgium
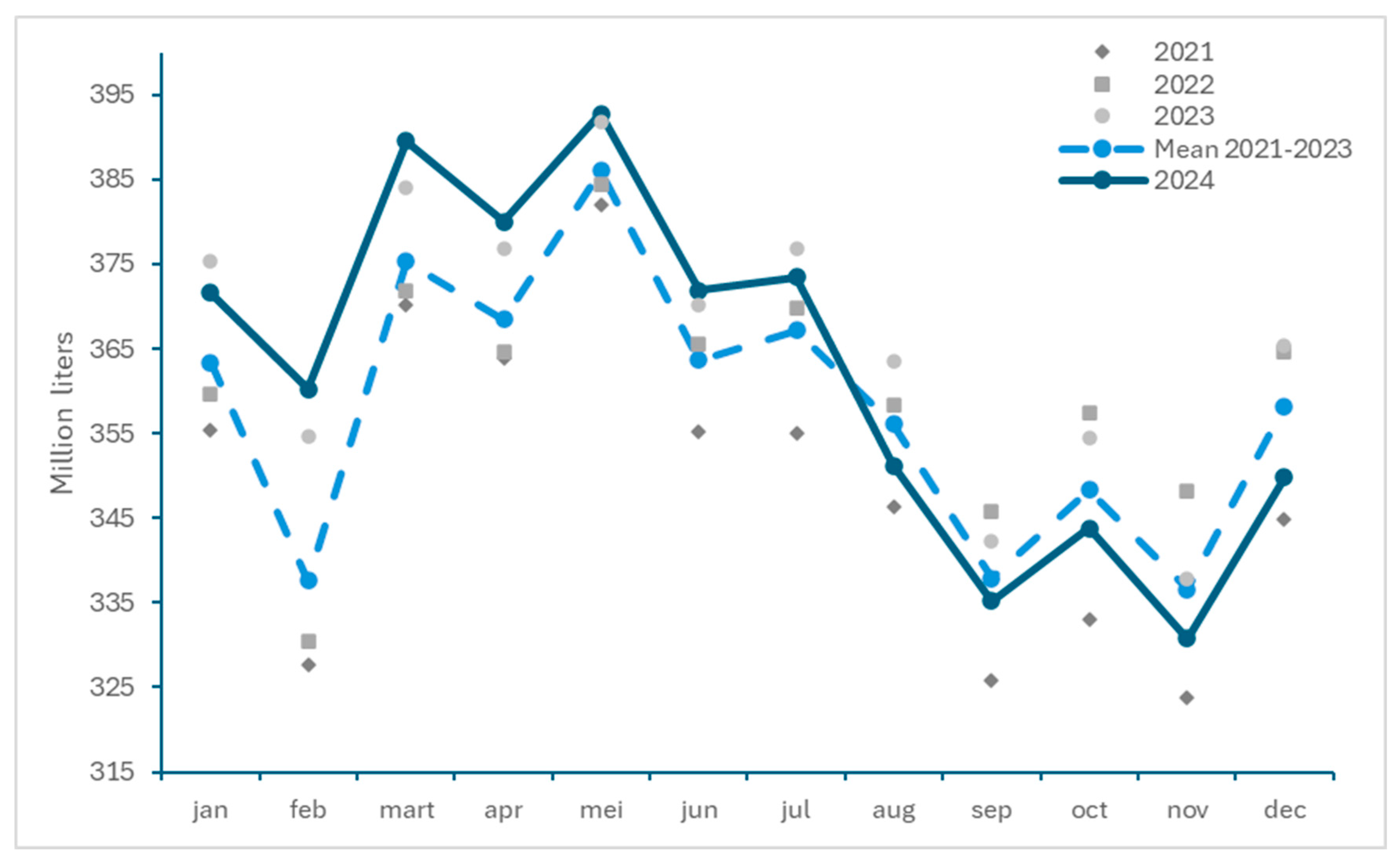
3.2. Impact of BTV-3 Circulation on Livestock Production
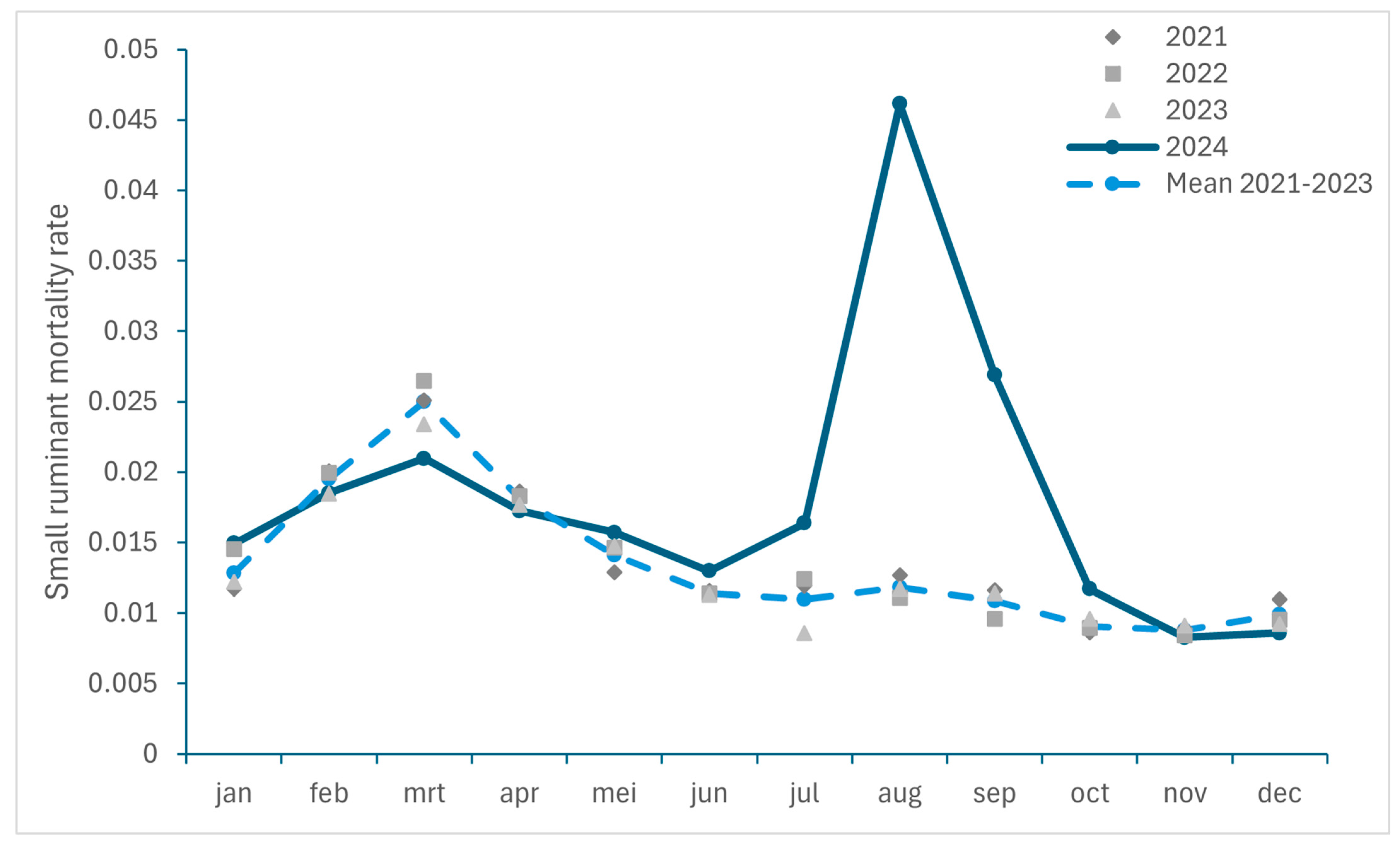
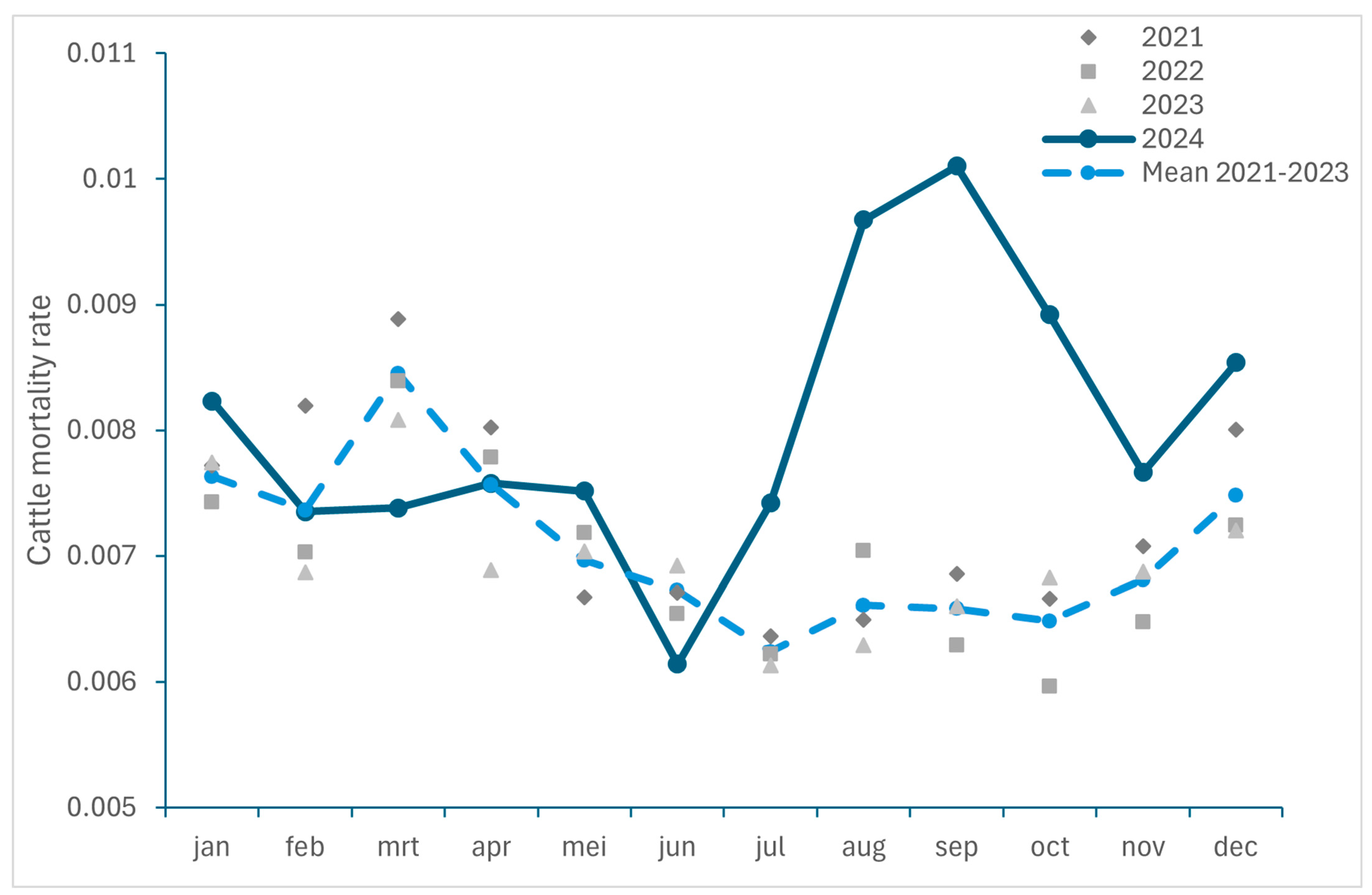
3.2.1. Excess Mortality in Cattle and Small Ruminants
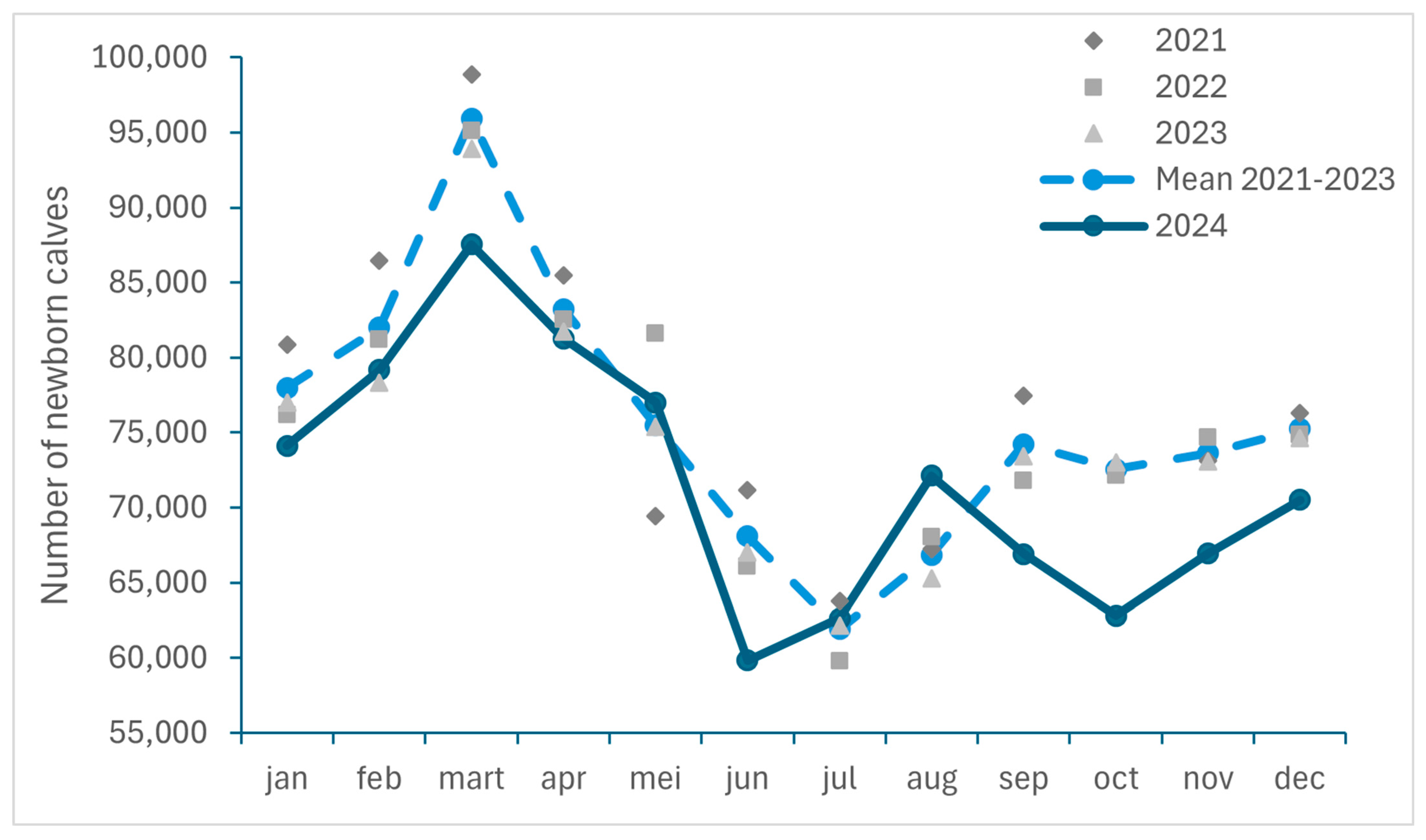
3.2.2. Decrease in Number of Newborn Calves
3.2.3. Impact on Milk Deliveries
3.2.4. Conclusions
3.3. Research Recommendations Based on Identified Knowledge Gaps
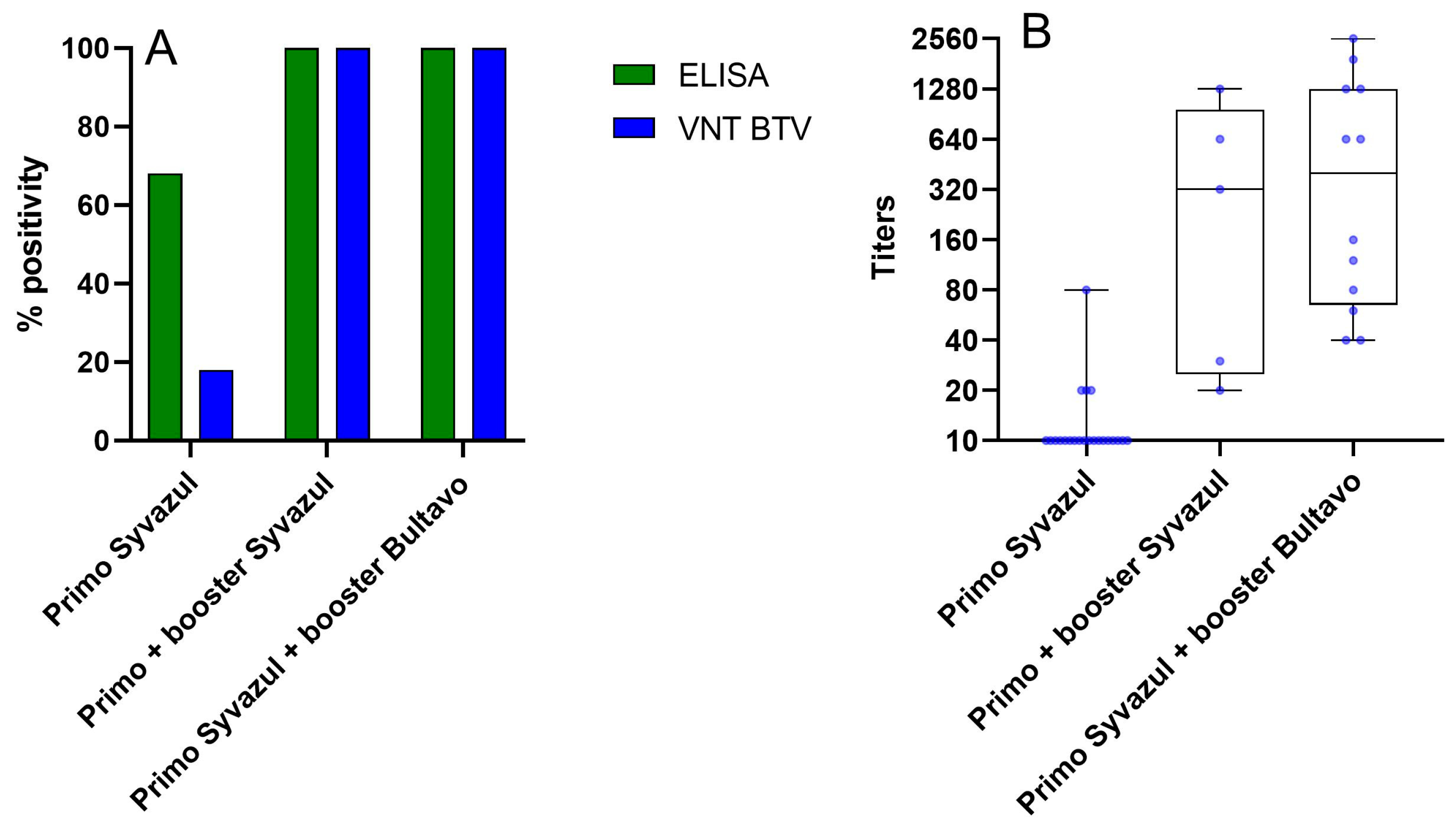
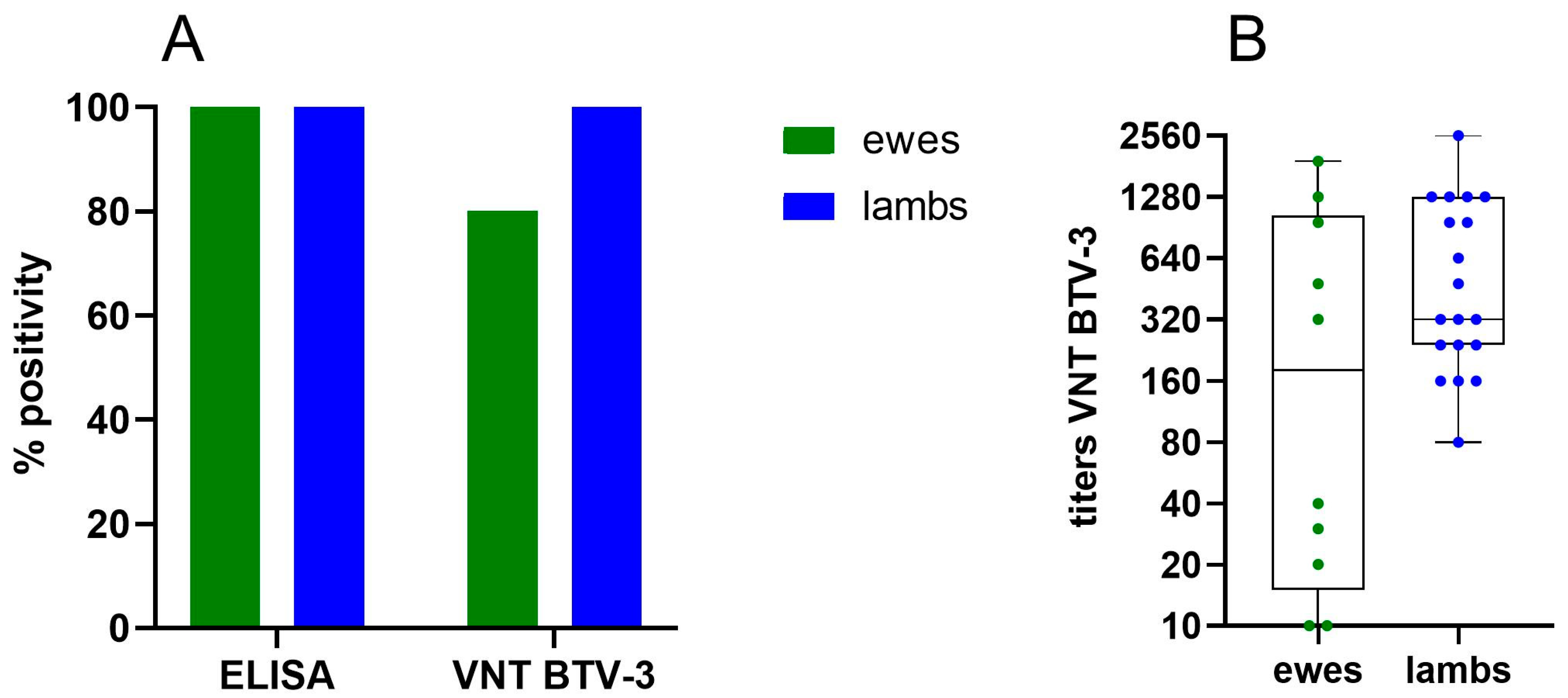
3.3.1. Additional Aspects on Vaccine Protection from BTV-3 Vaccination in the Field
3.3.2. BTV-3 Transmission Through Sperm and Embryos
3.3.3. Diagnostic Challenges
3.3.4. Role of Wildlife in BTV-3 Epidemiology
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BT | Bluetongue |
| BTV | Bluetongue Virus |
| EHDV | Epizootic Hemorrhagic Disease Virus |
| IgG | Immunoglobulin G |
| NRL | National Reference Laboratory |
| PCR | Polymerase Chain Reaction |
| VNT | Virus Neutralization test |
References
- Rodríguez-Martín, D.; Louloudes-Lázaro, A.; Avia, M.; Martín, V.; Rojas, J.M.; Sevilla, N. The Interplay between Bluetongue Virus Infections and Adaptive Immunity. Viruses 2021, 13, 1511. [Google Scholar] [CrossRef] [PubMed]
- WOAH. Terrestrial Manual 2021. Chapter 3.1.3. Bluetongue (Infection with Bluetongue Virus). Available online: https://www.woah.org/fileadmin/Home/fr/Health_standards/tahm/3.01.03_BLUETONGUE.pdf (accessed on 6 March 2025).
- Belbis, G.; Zientara, S.; Bréard, E.; Sailleau, C.; Caignard, G.; Vitour, D.; Attoui, H. Bluetongue Virus: From BTV-1 to BTV-27. Adv. Virus Res. 2017, 99, 161–197. [Google Scholar] [CrossRef] [PubMed]
- Ries, C.; Vögtlin, A.; Hüssy, D.; Jandt, T.; Gobet, H.; Hilbe, M.; Burgener, C.; Schweizer, L.; Häfliger-Speiser, S.; Beer, M.; et al. Putative Novel Atypical BTV Serotype ‘36’ Identified in Small Ruminants in Switzerland. Viruses 2021, 13, 721. [Google Scholar] [CrossRef]
- Spedicato, M.; Compagni, E.D.; Caporale, M.; Teodori, L.; Leone, A.; Ancora, M.; Mangone, I.; Perletta, F.; Portanti, O.; Di Giallonardo, F.; et al. Reemergence of an Atypical Bluetongue Virus Strain in Goats, Sardinia, Italy. Res. Vet. Sci. 2022, 151, 36–41. [Google Scholar] [CrossRef]
- Nomikou, K.; Hughes, J.; Wash, R.; Kellam, P.; Breard, E.; Zientara, S.; Palmarini, M.; Biek, R.; Mertens, P. Widespread Reassortment Shapes the Evolution and Epidemiology of Bluetongue Virus Following European Invasion. PLoS Pathog. 2015, 11, e1005056. [Google Scholar] [CrossRef]
- Saegerman, C.; Berkvens, D.; Mellor, P.S. Bluetongue Epidemiology in the European Union. Emerg. Infect. Dis. 2008, 14, 539–544. [Google Scholar] [CrossRef]
- Lorusso, A.; Guercio, A.; Purpari, G.; Cammà, C.; Calistri, P.; D’Alterio, N.; Hammami, S.; Sghaier, S.; Savini, G. Bluetongue virus serotype 3 in Western Sicily, November 2017. Vet. Ital. 2017, 53, 273–275. [Google Scholar] [CrossRef] [PubMed]
- Niedbalski, W. Bluetongue Virus in Europe: The Current Epidemiological Situation. Med. Weter. 2022, 78, 109–114. [Google Scholar] [CrossRef]
- Holwerda, M.; Santman-Berends, I.M.G.A.; Harders, F.; Engelsma, M.; Vloet, R.P.M.; Dijkstra, E.; van Gennip, R.G.P.; Mars, M.H.; Spierenburg, M.; Roos, L.; et al. Emergence of Bluetongue Virus Serotype 3 in the Netherlands in September 2023. Emerg. Infect. Dis. 2024, 30, 1552–1561. [Google Scholar] [CrossRef]
- Möhlmann, T.W.R.; Keeling, M.J.; Wennergren, U.; Favia, G.; Santman-Berends, I.; Takken, W.; Koenraadt, C.J.M.; Brand, S.P.C. Biting Midge Dynamics and Bluetongue Transmission: A Multiscale Model Linking Catch Data with Climate and Disease Outbreaks. Sci. Rep. 2021, 11, 1892. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control (ECDC) Biting Midge Maps. Available online: https://www.ecdc.europa.eu/en/disease-vectors/surveillance-and-disease-data/biting-midge-maps (accessed on 25 February 2025).
- Guyot, H.; Mauroy, A.; Kirschvink, N.; Rolin, F.; Saegerman, C. Clinical Aspects of Bluetongue in Ruminants. In Bluetongue in Northern Europe; Saegerman, C., Reviriego-Gorrdejo, F., Pastoret, P.P., Eds.; World Organisation for Animal Health: Paris, France, 2008; pp. 34–52. [Google Scholar]
- van den Brink, K.M.J.A.; Santman-Berends, I.M.G.A.; Harkema, L.; Scherpenzeel, C.G.M.; Dijkstra, E.; Bisschop, P.I.H.; Peterson, K.; van de Burgwal, N.S.; Waldeck, H.W.F.; Dijkstra, T.; et al. Bluetongue Virus Serotype 3 in Ruminants in the Netherlands: Clinical Signs, Seroprevalence and Pathological Findings. Vet. Rec. 2024, 195, e4533. [Google Scholar] [CrossRef] [PubMed]
- Saegerman, C.; Bolkaerts, B.; Baricalla, C.; Raes, M.; Wiggers, L.; de Leeuw, I.; Vandenbussche, F.; Zimmer, J.-Y.; Haubruge, E.; Cassart, D.; et al. The Impact of Naturally-Occurring, Trans-Placental Bluetongue Virus Serotype-8 Infection on Reproductive Performance in Sheep. Vet. J. 2011, 187, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Gethmann, J.; Probst, C.; Conraths, F.J. Economic Impact of a Bluetongue Serotype 8 Epidemic in Germany. Front. Vet. Sci. 2020, 7, 65. [Google Scholar] [CrossRef]
- Hanon, J.-B.; Uyttenhoef, A.; Fecher-Bourgeois, F.; Kirschvink, N.; Haubruge, E.; Duquesne, B.; Saegerman, C. Estimation quantitative des pertes économiques directes et indirectes subies par les éleveurs wallons dans le cadre de la fièvre catarrhale ovine (sérotype 8) durant la période 2006–2007. Epidémiologie Santé Anim. 2009, 56, 187–195. [Google Scholar]
- Santman-Berends, I.M.G.A.; van den Brink, K.M.J.A.; Dijkstra, E.; van Schaik, G.; Spierenburg, M.A.H.; van den Brom, R. The Impact of the Bluetongue Serotype 3 Outbreak on Sheep and Goat Mortality in the Netherlands in 2023. Prev. Vet. Med. 2024, 231, 106289. [Google Scholar] [CrossRef]
- van Den Brink, K.M.J.A.; Brouwer-Middelesch, H.; Van Schaik, G.; Lam, T.J.G.M.; Stegeman, A.; Van Den Brom, R.; Spierenburg, M.A.H.; Santman-Berends, I.M.G.A. The Impact of Bluetongue Serotype 3 on Cattle Mortality, Abortions and Premature Births in the Netherlands in the First Year of the Epidemic. Available online: https://ssrn.com/abstract=5056290 (accessed on 26 March 2025).
- Gondard, M.; Postic, L.; Garin, E.; Turpaud, M.; Vorimore, F.; Ngwa-Mbot, D.; Tran, M.-L.; Hoffmann, B.; Warembourg, C.; Savini, G.; et al. Exceptional Bluetongue Virus (BTV) and Epizootic Hemorrhagic Disease Virus (EHDV) Circulation in France in 2023. Virus Res. 2024, 350, 199489. [Google Scholar] [CrossRef]
- van den Brom, R.; Santman-Berends, I.; van der Heijden, M.G.; Harders, F.; Engelsma, M.; van Gennip, R.G.P.; Maris-Veldhuis, M.A.; Feddema, A.-J.; Peterson, K.; Golender, N.; et al. Bluetongue Virus Serotype 12 in Sheep and Cattle in the Netherlands in 2024–A BTV Serotype Reported in Europe for the First Time. Vet. Microbiol. 2025, 301, 110365. [Google Scholar] [CrossRef]
- Hofmann, M.; Griot, C.; Chaignat, V.; Perler, L.; Thür, B. Blauzungenkrankheit Erreicht Die Schweiz. Schweiz. Arch. Tierheilkd. 2008, 150, 49–56. [Google Scholar] [CrossRef]
- Lorusso, A.; Sghaier, S.; Di Domenico, M.; Barbria, M.E.; Zaccaria, G.; Megdich, A.; Portanti, O.; Ben Seliman, I.; Spedicato, M.; Pizzurro, F.; et al. Analysis of Bluetongue Serotype 3 Spread in Tunisia and Discovery of a Novel Strain Related to the Bluetongue Virus Isolated from a Commercial Sheep Pox Vaccine. Infect. Genet. Evol. 2018, 59, 63–71. [Google Scholar] [CrossRef]
- Viarouge, C.; Breard, E.; Zientara, S.; Vitour, D.; Sailleau, C. Duplex Real-Time RT-PCR Assays for the Detection and Typing of Epizootic Haemorrhagic Disease Virus. PLoS ONE 2015, 10, e0132540. [Google Scholar] [CrossRef]
- Pioz, M.; Guis, H.; Calavas, D.; Durand, B.; Abrial, D.; Ducrot, C. Estimating Front-Wave Velocity of Infectious Diseases: A Simple, Efficient Method Applied to Bluetongue. Vet. Res. 2011, 42, 60. [Google Scholar] [CrossRef] [PubMed]
- Martinelle, L.; Dal Pozzo, F.; Thys, C.; De Leeuw, I.; Van Campe, W.; De Clercq, K.; Thiry, E.; Saegerman, C. Assessment of Cross-Protection Induced by a Bluetongue Virus (BTV) Serotype 8 Vaccine towards Other BTV Serotypes in Experimental Conditions. Vet. Res. 2018, 49, 63. [Google Scholar] [CrossRef]
- Fay, P.C.; Jaafar, F.M.; Batten, C.; Attoui, H.; Saunders, K.; Lomonossoff, G.P.; Reid, E.; Horton, D.; Maan, S.; Haig, D.; et al. Serological Cross-Reactions between Expressed Vp2 Proteins from Different Bluetongue Virus Serotypes. Viruses 2021, 13, 1455. [Google Scholar] [CrossRef]
- Santman-Berends, I.M.G.A.; van Schaik, G.; Bartels, C.J.M.; Stegeman, J.A.; Vellema, P. Mortality Attributable to Bluetongue Virus Serotype 8 Infection in Dutch Dairy Cows. Vet. Microbiol. 2011, 148, 183–188. [Google Scholar] [CrossRef]
- Elbers, A.R.W.; Backx, A.; Mintiens, K.; Gerbier, G.; Staubach, C.; Hendrickx, G.; van der Spek, A. Field Observations during the Bluetongue Serotype 8 Epidemic in 2006. II. Morbidity and Mortality Rate, Case Fatality and Clinical Recovery in Sheep and Cattle in the Netherlands. Prev. Vet. Med. 2008, 87, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Saegerman, C.; Mellor, P.S.; Berkvens, D.; Pozzo, F.D. Epidémiologie de La Fièvre Catarrhale Ovine En Europe. In Fièvre Catarrhale Ovine; Éditions France Agricole: Paris, France, 2008; pp. 40–59. [Google Scholar]
- Conraths, F.J.; Gethmann, J.M.; Staubach, C.; Mettenleiter, T.C.; Beer, M.; Hoffmann, B. Epidemiology of Bluetongue Virus Serotype 8, Germany. Emerg. Infect. Dis. 2009, 15, 433–435. [Google Scholar] [CrossRef]
- Wouda, W.; Roumen, M.P.H.M.; Peperkamp, N.H.M.T.; Vos, J.H.; van Garderen, E.; Muskens, J. Hydranencephaly in Calves Following the Bluetongue Serotype 8 Epidemic in the Netherlands. Vet. Rec. 2008, 162, 422–423. [Google Scholar] [CrossRef]
- Vinomack, C.; Rivière, J.; Bréard, E.; Viarouge, C.; Postic, L.; Zientara, S.; Vitour, D.; Belbis, G.; Spony, V.; Pagneux, C.; et al. Clinical Cases of Bluetongue Serotype 8 in Calves in France in the 2018–2019 Winter. Transbound. Emerg. Dis. 2020, 67, 1401–1405. [Google Scholar] [CrossRef] [PubMed]
- Bulletin Épidémiologique N°35. Réseau Wallon D’Épidémiosurveillance Des Avortements Bovins. ARSIA. Available online: https://www.arsia.be/wp-content/uploads/PDF-Arsia-Infos/2025/AI-Janvier-2025-FR.pdf (accessed on 4 March 2025).
- Santman-Berends, I.M.G.A.; Hage, J.J.; van Rijn, P.A.; Stegeman, J.A.; van Schaik, G. Bluetongue Virus Serotype 8 (BTV-8) Infection Reduces Fertility of Dutch Dairy Cattle and Is Vertically Transmitted to Offspring. Theriogenology 2010, 74, 1377–1384. [Google Scholar] [CrossRef]
- Kirschvink, N.; Raes, M.; Saegerman, C. Impact of a Natural Bluetongue Serotype 8 Infection on Semen Quality of Belgian Rams in 2007. Vet. J. 2009, 182, 244–251. [Google Scholar] [CrossRef]
- Müller, U.; Kemmerling, K.; Straet, D.; Janowitz, U.; Sauerwein, H. Effects of Bluetongue Virus Infection on Sperm Quality in Bulls: A Preliminary Report. Vet. J. 2010, 186, 402–403. [Google Scholar] [CrossRef]
- Leemans, J.; Raes, M.; Vanbinst, T.; De Clercq, K.; Saegerman, C.; Kirschvink, N. Viral RNA Load in Semen from Bluetongue Serotype 8-Infected Rams: Relationship with Sperm Quality. Vet. J. 2012, 192, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Martinelle, L.; Dal Pozzo, F.; Thiry, E.; De Clercq, K.; Saegerman, C. Reliable and Standardized Animal Models to Study the Pathogenesis of Bluetongue and Schmallenberg Viruses in Ruminant Natural Host Species with Special Emphasis on Placental Crossing. Viruses 2019, 11, 753. [Google Scholar] [CrossRef] [PubMed]
- Nusinovici, S.; Seegers, H.; Joly, A.; Beaudeau, F.; Fourichon, C. Quantification and At-Risk Period of Decreased Fertility Associated with Exposure to Bluetongue Virus Serotype 8 in Naïve Dairy Herds. J. Dairy Sci. 2012, 95, 3008–3020. [Google Scholar] [CrossRef] [PubMed]
- Melkleveringen (BCZ CBL). Available online: https://bcz-cbl.be/nl/domeinen/melkaanvoer/melkleveringen (accessed on 4 March 2025).
- Melkaanvoer en Zuivelproductie Door Zuivelfabrieken (Central Bureau Voor de Statistiek). Available online: https://www.cbs.nl/nl-nl/cijfers/detail/7425zuiv (accessed on 4 March 2025).
- Cargnel, M.; Van der Stede, Y.; Haegeman, A.; De Leeuw, I.; De Clercq, K.; Méroc, E.; Welby, S. Effectiveness and Cost-Benefit Study to Encourage Herd Owners in a Cost Sharing Vaccination Programme against Bluetongue Serotype-8 in Belgium. Transbound. Emerg. Dis. 2019, 66, 400–411. [Google Scholar] [CrossRef]
- Velthuis, A.G.J.; Saatkamp, H.W.; Mourits, M.C.M.; de Koeijer, A.A.; Elbers, A.R.W. Financial Consequences of the Dutch Bluetongue Serotype 8 Epidemics of 2006 and 2007. Prev. Vet. Med. 2010, 93, 294–304. [Google Scholar] [CrossRef]
- Vitour, D.; Guillotin, J.; Sailleau, C.; Viarouge, C.; Desprat, A.; Wolff, F.; Belbis, G.; Durand, B.; Bakkali-Kassimi, L.; Breard, E.; et al. Colostral Antibody Induced Interference of Inactivated Bluetongue Serotype-8 Vaccines in Calves. Vet. Res. 2011, 42, 18. [Google Scholar] [CrossRef]
- van der Sluijs, M.T.W.; Schroer-Joosten, D.P.H.; Fid-Fourkour, A.; Vrijenhoek, M.P.; Debyser, I.; Gregg, D.A.; Dufe, D.M.; Moulin, V.; Moormann, R.J.M.; de Smit, A.J. Effect of Vaccination with an Inactivated Vaccine on Transplacental Transmission of BTV-8 in Mid Term Pregnant Ewes and Heifers. Vaccine 2012, 30, 647–655. [Google Scholar] [CrossRef]
- Leemans, J.; Hamers, C.; Chery, R.; Bibard, A.; Besancon, L.; Duboeuf, M.; Hudelet, P.; Goutebroze, S.; Kirschvink, N. Interference of Colostral Antibodies with Response to a Bluetongue Serotype 8 Inactivated Vaccine in Lambs Born from Hyperimmune Ewes. Vaccine 2013, 31, 1975–1980. [Google Scholar] [CrossRef]
- Farooq, U.; Ahmed, S.; Liu, G.; Jiang, X.; Yang, H.; Ding, J.; Ali, M. Biochemical Properties of Sheep Colostrum and Its Potential Benefits for Lamb Survival: A Review. Anim. Biotechnol. 2024, 35, 2320726. [Google Scholar] [CrossRef]
- EFSA AHAW Panel (EFSA Panel on Animal Health and Welfare). Scientific opinion on bluetongue: Control, Surveillance and Safe Movement of Animals. EFSA J. 2017, 15, e04698. [Google Scholar] [CrossRef]
- Kirkland, P.D.; Melville, L.F.; Hunt, N.T.; Williams, C.F.; Davis, R.J. Excretion of Bluetongue Virus in Cattle Semen: A Feature of Laboratory-Adapted Virus. Vet. Ital. 2004, 40, 497–501. [Google Scholar] [PubMed]
- EFSA AHAW Panel (EFSA Panel on Animal Health and Welfare). Scientific Opinion on Bluetongue Serotype 8. EFSA J. 2011, 9, 2189. [Google Scholar] [CrossRef]
- De Clercq, K.; Vandaele, L.; Vanbinst, T.; Riou, M.; Deblauwe, I.; Wesselingh, W.; Pinard, A.; Van Eetvelde, M.; Boulesteix, O.; Leemans, B.; et al. Transmission of Bluetongue Virus Serotype 8 by Artificial Insemination with Frozen–Thawed Semen from Naturally Infected Bulls. Viruses 2021, 13, 652. [Google Scholar] [CrossRef] [PubMed]
- Wrathall, A.E.; Simmons, H.A.; Van Soom, A. Evaluation of Risks of Viral Transmission to Recipients of Bovine Embryos Arising from Fertilisation with Virus-Infected Semen. Theriogenology 2006, 65, 247–274. [Google Scholar] [CrossRef]
- Haegeman, A.; Vandaele, L.; De Leeuw, I.; Oliveira, A.P.; Nauwynck, H.; Van Soom, A.; De Clercq, K. Failure to Remove Bluetongue Serotype 8 Virus (BTV-8) From in Vitro Produced and in Vivo Derived Bovine Embryos and Subsequent Transmission of BTV-8 to Recipient Cows After Embryo Transfer. Front. Vet. Sci. 2019, 6, 432. [Google Scholar] [CrossRef]
- Linden, A.; Grégoire, F.; Nahayo, A.; Hanrez, D.; Mousset, B.; Massart, L.; de Leeuw, I.; Vandemeulebroucke, E.; Vandenbussche, F.; de Clercq, K. Bluetongue Virus in Wild Deer, Belgium, 2005–2008. Emerg. Infect. Dis. 2010, 16, 833–836. [Google Scholar] [CrossRef]
- Mauroy, A.; Guyot, H.; De Clercq, K.; Cassart, D.; Thiry, E.; Saegerman, C. Bluetongue in Captive Yaks. Emerg. Infect. Dis. 2008, 14, 675–676. [Google Scholar] [CrossRef]
- Caballero-Gómez, J.; Cano Terriza, D.; Pujols, J.; Martínez-Nevado, E.; Carbonell, M.D.; Guerra, R.; Recuero, J.; Soriano, P.; Barbero, J.; García-Bocanegra, I. Monitoring of Bluetongue Virus in Zoo Animals in Spain, 2007–2019. Transbound. Emerg. Dis. 2022, 69, 1739–1747. [Google Scholar] [CrossRef]
- Martinelle, L.; Haegeman, A.; Lignereux, L.; Chaber, A.-L.; Dal Pozzo, F.; De Leeuw, I.; De Clercq, K.; Saegerman, C. Orbivirus Screening from Imported Captive Oryx in the United Arab Emirates Stresses the Importance of Pre-Import and Transit Measures. Pathogens 2022, 11, 697. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Leeuw, V.; De Leeuw, I.; Degives, N.; Depoorter, P.; Dewulf, J.; Hanon, J.-B.; Hooyberghs, J.; Linden, A.; Praet, L.; Raemaekers, M.; et al. Impact of BTV-3 Circulation in Belgium in 2024 and Current Knowledge Gaps Hindering an Evidence-Based Control Program. Viruses 2025, 17, 521. https://doi.org/10.3390/v17040521
Van Leeuw V, De Leeuw I, Degives N, Depoorter P, Dewulf J, Hanon J-B, Hooyberghs J, Linden A, Praet L, Raemaekers M, et al. Impact of BTV-3 Circulation in Belgium in 2024 and Current Knowledge Gaps Hindering an Evidence-Based Control Program. Viruses. 2025; 17(4):521. https://doi.org/10.3390/v17040521
Chicago/Turabian StyleVan Leeuw, Virginie, Ilse De Leeuw, Nicolas Degives, Pieter Depoorter, Jeroen Dewulf, Jean-Baptiste Hanon, Jozef Hooyberghs, Annick Linden, Laura Praet, Marc Raemaekers, and et al. 2025. "Impact of BTV-3 Circulation in Belgium in 2024 and Current Knowledge Gaps Hindering an Evidence-Based Control Program" Viruses 17, no. 4: 521. https://doi.org/10.3390/v17040521
APA StyleVan Leeuw, V., De Leeuw, I., Degives, N., Depoorter, P., Dewulf, J., Hanon, J.-B., Hooyberghs, J., Linden, A., Praet, L., Raemaekers, M., Saegerman, C., Simons, X., Sohier, C., Steurbaut, N., Sury, A., Thiry, E., Zientara, S., Mauroy, A., & De Regge, N. (2025). Impact of BTV-3 Circulation in Belgium in 2024 and Current Knowledge Gaps Hindering an Evidence-Based Control Program. Viruses, 17(4), 521. https://doi.org/10.3390/v17040521