
In Vitro System for Studying Ilhéus Virus, a Neglected Arbovirus: Ultrastructural Characterization of Cytopathology, Morphology, and Morphogenesis

 , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. ILHV Strain
2.2. Cell Lineage
2.3. Clarification and Viral Titer
2.4. Statistical Analysis
2.5. Infection Kinetics
2.6. Analysis of Cytopathic Effects by Bright-Field Microscopy
2.7. Processing of Cell Monolayers for TEM Analysis
2.8. Quantitative Analysis of Viral Particle Size
2.9. Sequencing and Representative Phylogenetic Tree of ILHV
3. Results
3.1. Sequencing of ILHV
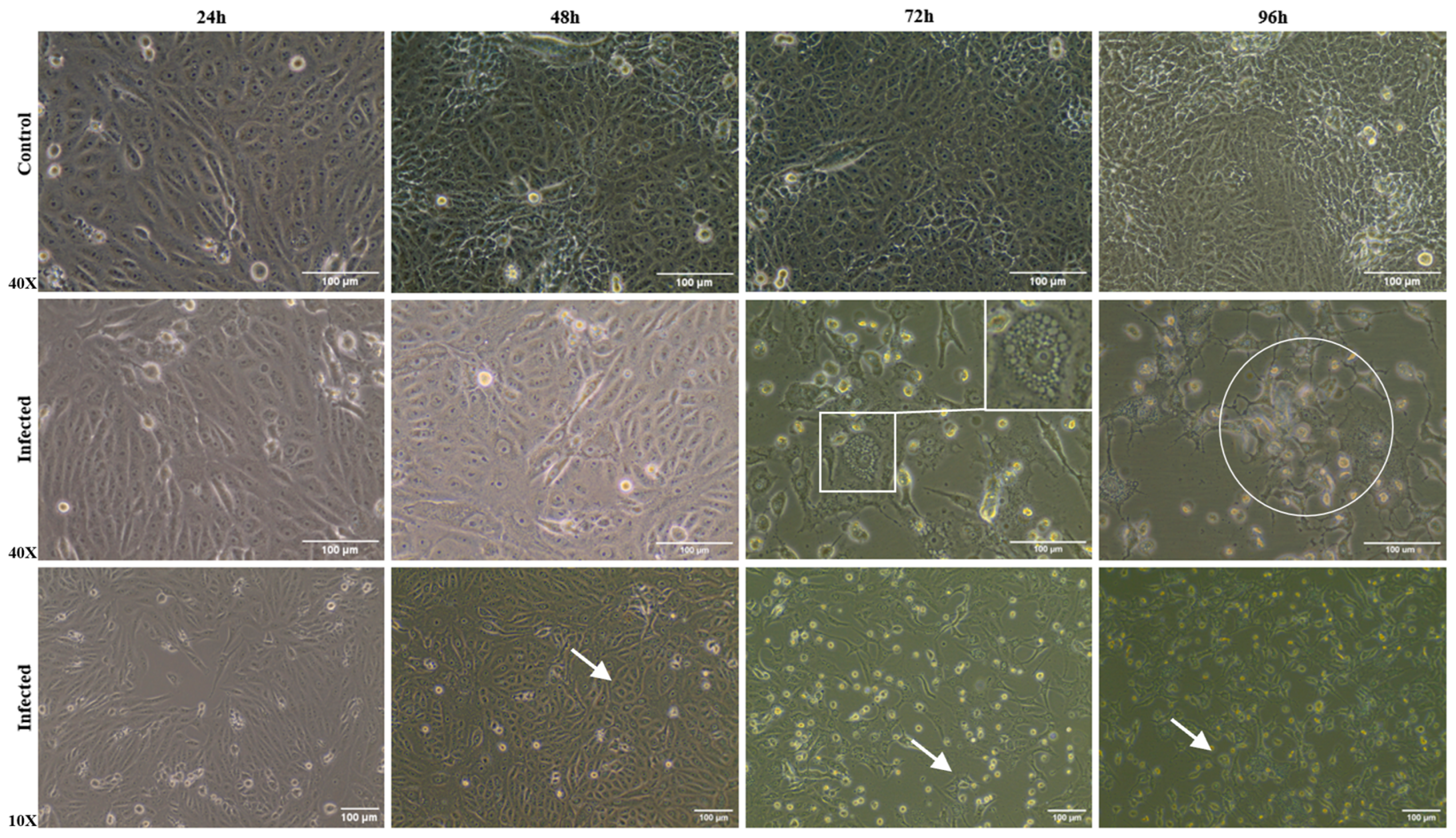
3.2. Morphological Alterations of Infected Cells
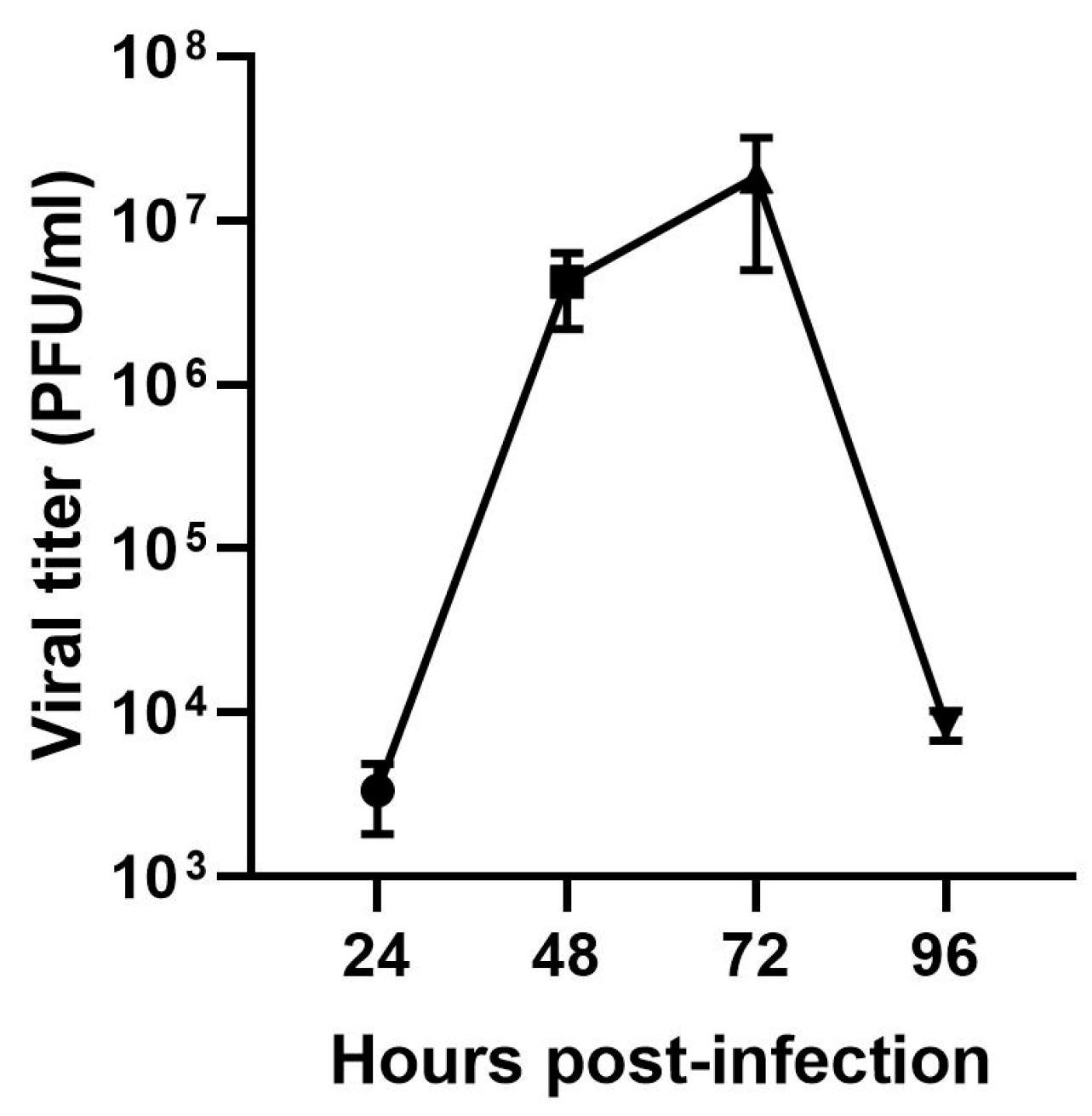
3.3. Infectious Viral Particles in the Supernatant of Vero Cells
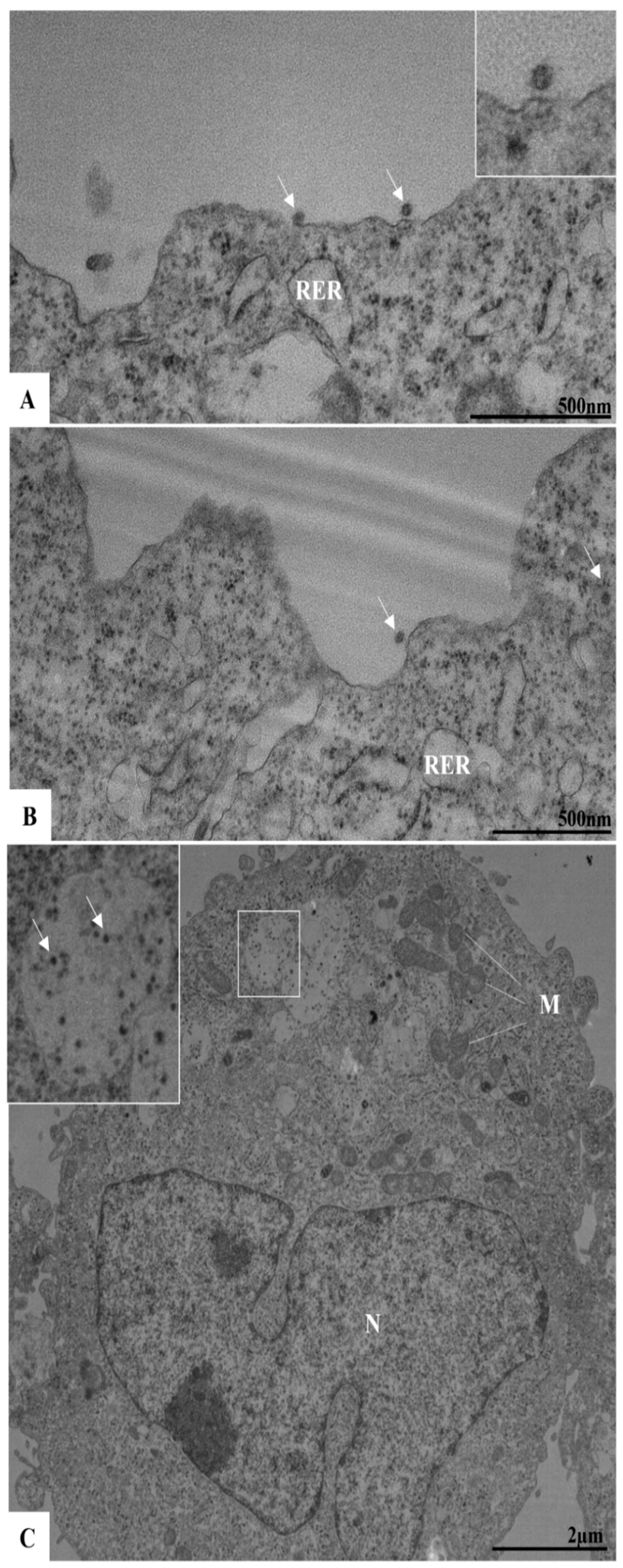
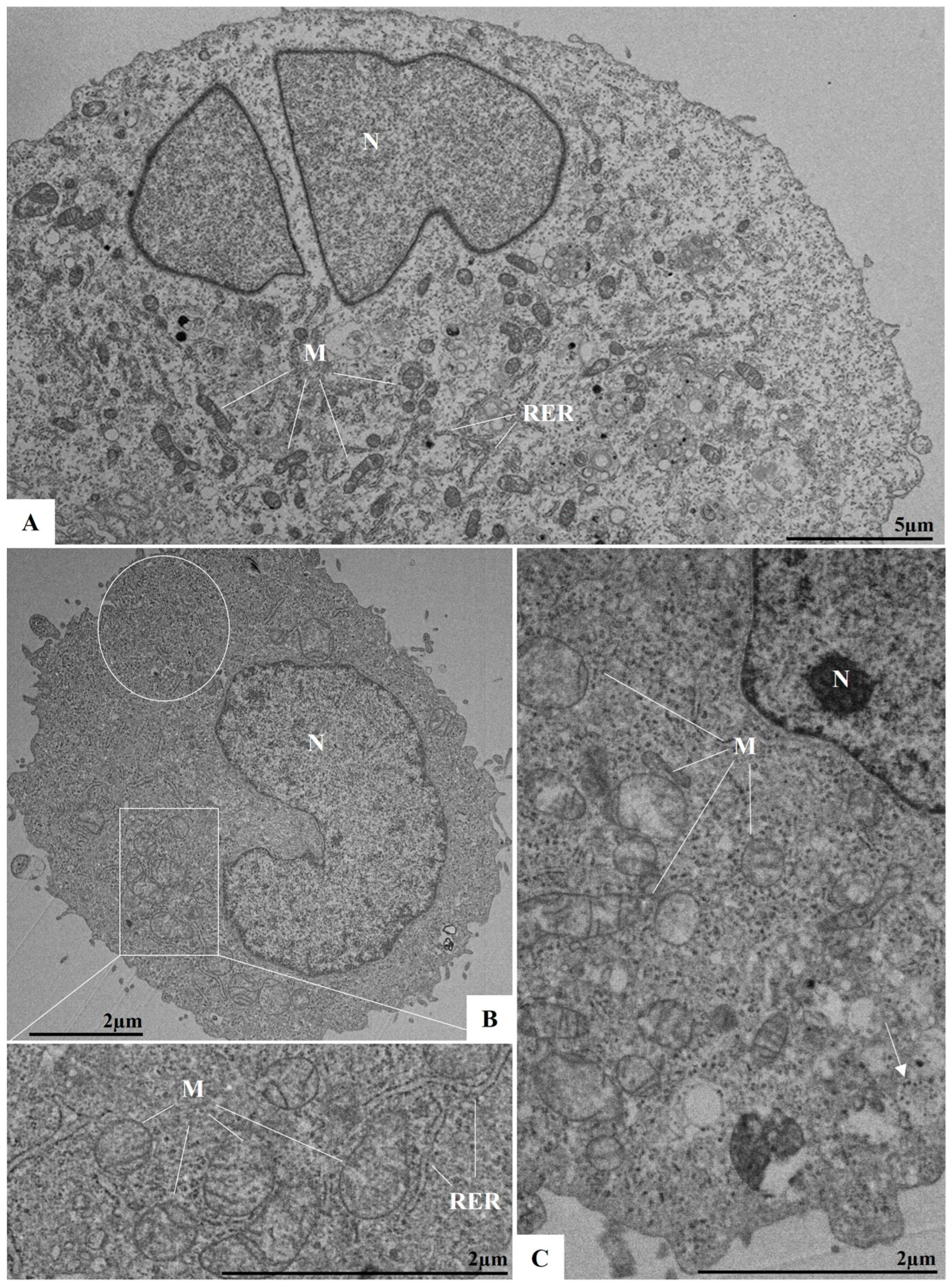
3.4. Ultrastructural Analyses by TEM of Infected Cells
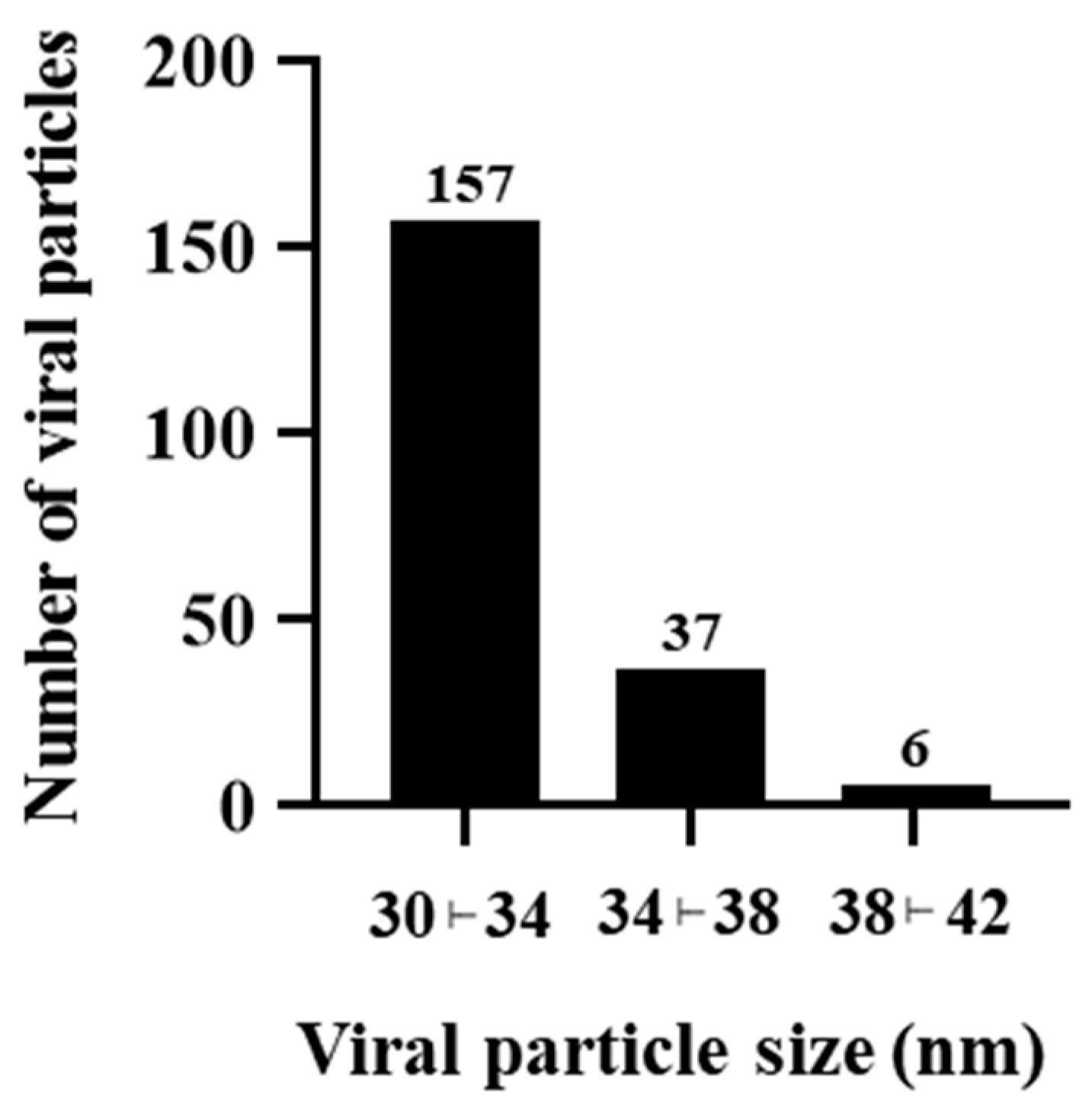
3.5. ILHV Particle Diameter
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laemmert, H.W.; Hughes, T.P. The Virus of Ilhéus Encephalitis. J. Immunol. 1947, 55, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Pauvolid-Corrêa, A.; Kenney, J.L.; Couto-Lima, D.; Campos, Z.M.S.; Schatzmayr, H.G.; Nogueira, R.M.R.; Brault, A.C.; Komar, N. Ilheus Virus Isolation in the Pantanal, West-Central Brazil. PLoS Negl. Trop. Dis. 2013, 7, e2318. [Google Scholar] [CrossRef] [PubMed]
- Vieira, C.J.d.S.P.; de Andrade, C.D.; Kubiszeski, J.R.; da Silva, D.J.F.; Barreto, E.S.; Massey, A.L.; Canale, G.R.; Bernardo, C.S.S.; Levi, T.; Peres, C.A.; et al. Detection of Ilheus Virus in Mosquitoes from Southeast Amazon, Brazil. Trans. R. Soc. Trop. Med. Hyg. 2019, 113, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Iversson, L.B.; Silva, R.A.M.; da Rosa, A.P.T.; Barros, V.L.R. Circulation of Eastern Equine Encephalitis, Western Equine Encephalitis, Ilhéus, Maguari and Tacaiuma Viruses in Equines of the Brazilian Pantanal, South America. Rev. Inst. Med. Trop. São Paulo 1993, 35, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, I.B.; Pereira, L.E.; Rocco, I.M.; Marti, A.T.; de Souza, L.T.M.; Iversson, L.B. Surveillance of Arbovirus Infections in the Atlantic Forest Region, State of São Paulo, Brazil: I. Detection of Hemagglutination-Inhibition Antibodies in Wild Birds between 1978 and 1990. Rev. Inst. Med. Trop. São Paulo 1994, 36, 265–274. [Google Scholar] [CrossRef]
- Pereira, L.E.; Suzuki, A.; Coimbra, T.L.M.; de Souza, R.P.; Chamelet, E.L.B. Arbovírus Ilheus em aves silvestres (Sporophila caerulescens e Molothrus bonariensis). Rev. Saúde Pública 2001, 35, 119–123. [Google Scholar] [CrossRef]
- Casseb, A.R.; Cruz, A.V.; Jesus, I.S.; Chiang, J.O.; Martins, L.C.; Silva, S.P.; Henriques, D.F.; Casseb, L.M.; Vasconcelos, P.F.C. Seroprevalence of Flaviviruses Antibodies in Water Buffaloes (Bubalus bubalis) in Brazilian Amazon. J. Venom. Anim. Toxins Incl. Trop. Dis. 2014, 20, 9. [Google Scholar] [CrossRef]
- Morales, M.A.; Fabbri, C.M.; Zunino, G.E.; Kowalewski, M.M.; Luppo, V.C.; Enría, D.A.; Levis, S.C.; Calderón, G.E. Detection of the Mosquito-Borne Flaviviruses, West Nile, Dengue, Saint Louis Encephalitis, Ilheus, Bussuquara, and Yellow Fever in Free-Ranging Black Howlers (Alouatta caraya) of Northeastern Argentina. PLoS Neglected Trop. Dis. 2017, 11, e0005351. [Google Scholar] [CrossRef]
- de Almeida, M.A.B.; dos Santos, E.; Cardoso, J.d.C.; Noll, C.A.; Lima, M.d.M.; Silva, F.d.A.e.; Ferreira, M.S.; Martins, L.C.; Vasconcelos, P.F.d.C.; Bicca-Marques, J.C. Detection of Antibodies against Icoaraci, Ilhéus, and Saint Louis Encephalitis Arboviruses during Yellow Fever Monitoring Surveillance in Non-Human Primates (Alouatta caraya) in Southern Brazil. J. Med. Primatol. 2019, 48, 211–217. [Google Scholar] [CrossRef]
- Causey, O.R.; Theiler, M. Virus Antibody Survey on Sera of Residents of the Amazon Valley in Brazil. Am. J. Trop. Med. Hyg. 1958, 7, 36–41. [Google Scholar] [CrossRef]
- Nassar, E.; Coimbra, T.; Rocco, I.; Pereira, L.; Ferreira, I.; Desouza, L.; de Souza, D.; Ueda-Ito, M.; Moura, J.; Bergo, R. Human Disease Caused by an Arbovirus Closely Related to Ilheus Virus: Report of Five Cases. Intervirology 1997, 40, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Milhim, B.H.G.A.; da Rocha, L.C.; Terzian, A.C.B.; Mazaro, C.C.P.; Augusto, M.T.; Luchs, A.; Zini, N.; Sacchetto, L.; dos Santos, B.F.; Garcia, P.H.C.; et al. Arboviral Infections in Neurological Disorders in Hospitalized Patients in São José Do Rio Preto, São Paulo, Brazil. Viruses 2022, 14, 1488. [Google Scholar] [CrossRef] [PubMed]
- Venegas, E.A.; Aguilar, P.V.; Cruz, C.; Guevara, C.; Kochel, T.J.; Vargas, J.; Halsey, E.S. Ilheus Virus Infection in Human, Bolivia. Emerg. Infect. Dis. J. CDC 2012, 18, 516–518. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.W.; Cruz, C.; Felices, V.; Espinoza, W.R.; Manock, S.R.; Guevara, C.; Olson, J.G.; Kochel, T.J. Ilheus Virus Isolate from a Human, Ecuador. Emerg. Infect. Dis. 2007, 13, 956–958. [Google Scholar] [CrossRef]
- Spence, L.; Anderson, C.R.; Downs, W.G. Isolation of Ilhéus Virus from Human Beings in Trinidad, West Indies. Trans. R. Soc. Trop. Med. Hyg. 1962, 56, 504–509. [Google Scholar] [CrossRef]
- Salgado, B.B.; de Jesus Maués, F.C.; Pereira, R.L.; Chiang, J.O.; de Oliveira Freitas, M.N.; Ferreira, M.S.; Martins, L.C.; da Costa Vasconcelos, P.F.; Ganoza, C.; Lalwani, P. Prevalence of Arbovirus Antibodies in Young Healthy Adult Population in Brazil. Parasites Vectors 2021, 14, 403. [Google Scholar] [CrossRef]
- da Costa, V.G.; Saivish, M.V.; Lino, N.A.B.; Bittar, C.; de Freitas Calmon, M.; Nogueira, M.L.; Rahal, P. Clinical Landscape and Rate of Exposure to Ilheus Virus: Insights from Systematic Review and Meta-Analysis. Viruses 2022, 15, 92. [Google Scholar] [CrossRef]
- Postler, T.S.; Beer, M.; Blitvich, B.J.; Bukh, J.; De Lamballerie, X.; Drexler, J.F.; Imrie, A.; Kapoor, A.; Karganova, G.G.; Lemey, P.; et al. Renaming of the Genus Flavivirus to Orthoflavivirus and Extension of Binomial Species Names within the Family Flaviviridae. Arch. Virol. 2023, 168, 224. [Google Scholar] [CrossRef]
- Plante, J.A.; Plante, K.S.; Popov, V.L.; Shinde, D.P.; Widen, S.G.; Buenemann, M.; Nogueira, M.L.; Vasilakis, N. Morphologic and Genetic Characterization of Ilheus Virus, a Potential Emergent Flavivirus in the Americas. Viruses 2023, 15, 195. [Google Scholar] [CrossRef]
- Klema, V.; Padmanabhan, R.; Choi, K. Flaviviral Replication Complex: Coordination between RNA Synthesis and 5′-RNA Capping. Viruses 2015, 7, 4640–4656. [Google Scholar] [CrossRef]
- Mackenzie, J.M.; Jones, M.K.; Young, P.R. Immunolocalization of the Dengue Virus Nonstructural Glycoprotein NS1 Suggests a Role in Viral RNA Replication. Virology 1996, 220, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Junjhon, J.; Pennington, J.G.; Edwards, T.J.; Perera, R.; Lanman, J.; Kuhn, R.J. Ultrastructural Characterization and Three-Dimensional Architecture of Replication Sites in Dengue Virus-Infected Mosquito Cells. J. Virol. 2014, 88, 4687–4697. [Google Scholar] [CrossRef]
- Cortese, M.; Goellner, S.; Acosta, E.G.; Neufeldt, C.J.; Oleksiuk, O.; Lampe, M.; Haselmann, U.; Funaya, C.; Schieber, N.; Ronchi, P.; et al. Ultrastructural Characterization of Zika Virus Replication Factories. Cell Rep. 2017, 18, 2113–2123. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, E.D.; Peters, K.N.; Connor, J.H.; Bullitt, E. Zika Virus Induced Cellular Remodelling. Cell. Microbiol. 2017, 19, e12740. [Google Scholar] [CrossRef] [PubMed]
- Barreto-Vieira, D.F.; Jácome, F.C.; Da Silva, M.A.N.; Caldas, G.C.; De Filippis, A.M.B.; De Sequeira, P.C.; De Souza, E.M.; Andrade, A.A.; Manso, P.P.D.A.; Trindade, G.F.; et al. Structural Investigation of C6/36 and Vero Cell Cultures Infected with a Brazilian Zika Virus. PLoS ONE 2017, 12, e0184397. [Google Scholar] [CrossRef] [PubMed]
- Rivera, J.A.; Rengifo, A.C.; Sarmiento, L.; Díaz, T.; Laiton-Donato, K.; Gracia, M.; Camacho, S.; Velandia-Romero, M.; Castellanos, J.; Caldas, M.L. Nuclei Ultrastructural Changes of C6/36 Cells Infected with Virus Dengue Type 2. Biomedica 2018, 38, 135–143. [Google Scholar] [CrossRef]
- Plante, K.S.; Plante, J.A.; Azar, S.R.; Shinde, D.P.; Scharton, D.; Versiani, A.F.; Oliveira Da Silva, N.I.; Strange, T.; Sacchetto, L.; Fokam, E.B.; et al. Potential of Ilhéus Virus to Emerge. Heliyon 2024, 10, e27934. [Google Scholar] [CrossRef]
- Barreto-Vieira, D.F. Modelo Animal Experimental Para o Estudo da Patogênese dos Vírus Dengue Sorotipos 1 e 2; Interciência: Rio de Janeiro, Brazil, 2010. [Google Scholar]
- Barth, O.M.; Silva, M.A.N.D.; Barreto-Vieira, D.F. Low Impact to Fixed Cell Processing Aiming Transmission Electron Microscopy. Mem. Inst. Oswaldo Cruz 2016, 111, 411–413. [Google Scholar] [CrossRef]
- Reynolds, E.S. The Use of Lead Citrate at High pH as An Electron-Opaque Stain In Electron Microscopy. J. Cell Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef]
- Vilsker, M.; Moosa, Y.; Nooij, S.; Fonseca, V.; Ghysens, Y.; Dumon, K.; Pauwels, R.; Alcantara, L.C.; Vanden Eynden, E.; Vandamme, A.-M.; et al. Genome Detective: An Automated System for Virus Identification from High-Throughput Sequencing Data. Bioinformatics 2019, 35, 871–873. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000; ISBN 978-0-19-535051-7. [Google Scholar]
- Chatel-Chaix, L.; Bartenschlager, R. Dengue Virus- and Hepatitis C Virus-Induced Replication and Assembly Compartments: The Enemy Inside—Caught in the Web. J. Virol. 2014, 88, 5907–5911. [Google Scholar] [CrossRef] [PubMed]
- McCormick, K.D.; Liu, S.; Jacobs, J.L.; Marques, E.T.A.; Sluis-Cremer, N.; Wang, T. Development of a Robust Cytopathic Effect-Based High-Throughput Screening Assay To Identify Novel Inhibitors of Dengue Virus. Antimicrob. Agents Chemother. 2012, 56, 3399–3401. [Google Scholar] [CrossRef] [PubMed]
- Offerdahl, D.K.; Dorward, D.W.; Hansen, B.T.; Bloom, M.E. Cytoarchitecture of Zika Virus Infection in Human Neuroblastoma and Aedes Albopictus Cell Lines. Virology 2017, 501, 54–62. [Google Scholar] [CrossRef]
- Kuhn, R.J.; Zhang, W.; Rossmann, M.G.; Pletnev, S.V.; Corver, J.; Lenches, E.; Jones, C.T.; Mukhopadhyay, S.; Chipman, P.R.; Strauss, E.G.; et al. Structure of Dengue Virus: Implications for Flavivirus Organization, Maturation, and Fusion. Struct. Dengue Virus Implic. Flavivirus Organ. Matur. Fusion 2002, 108, 717–725. [Google Scholar]
- Mukhopadhyay, S.; Kim, B.-S.; Chipman, P.R.; Rossmann, M.G.; Kuhn, R.J. Structure of West Nile Virus. Science 2003, 302, 248. [Google Scholar] [CrossRef]
- Morita, E.; Suzuki, Y. Membrane-Associated Flavivirus Replication Complex—Its Organization and Regulation. Viruses 2021, 13, 1060. [Google Scholar] [CrossRef]
- Bílý, T.; Palus, M.; Eyer, L.; Elsterová, J.; Vancová, M.; Růžek, D. Electron Tomography Analysis of Tick-Borne Encephalitis Virus Infection in Human Neurons. Sci. Rep. 2015, 5, 10745. [Google Scholar] [CrossRef]
- Arakawa, M.; Morita, E. Flavivirus Replication Organelle Biogenesis in the Endoplasmic Reticulum: Comparison with Other Single-Stranded Positive-Sense RNA Viruses. Int. J. Mol. Sci. 2019, 20, 2336. [Google Scholar] [CrossRef]
- McLauchlan, J. Lipid Droplets and Hepatitis C Virus Infection. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2009, 1791, 552–559. [Google Scholar] [CrossRef]
- Dias, S.S.G.; Soares, V.C.; Ferreira, A.C.; Sacramento, C.Q.; Fintelman-Rodrigues, N.; Temerozo, J.R.; Teixeira, L.; da Silva, M.A.N.; Barreto, E.; Mattos, M.; et al. Lipid Droplets Fuel SARS-CoV-2 Replication and Production of Inflammatory Mediators. PLoS Pathogens 2020, 16, e1009127. [Google Scholar] [CrossRef]
- Dias, S.S.G.; Cunha-Fernandes, T.; Souza-Moreira, L.; Soares, V.C.; Lima, G.B.; Azevedo-Quintanilha, I.G.; Santos, J.; Pereira-Dutra, F.; Freitas, C.; Reis, P.A.; et al. Metabolic Reprogramming and Lipid Droplets Are Involved in Zika Virus Replication in Neural Cells. J. Neuroinflamm. 2023, 20, 61. [Google Scholar] [CrossRef]
- Miller, M.A.; Zachary, J.F. Mechanisms and Morphology of Cellular Injury, Adaptation, and Death. In Pathologic Basis of Veterinary Disease, 6th ed.; Zachary, J.F., Ed.; Mosby: Maryland Heights, MD, USA, 2017; pp. 2–43.e19. ISBN 978-0-323-35775-3. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia, M.D.d.N.; Da Costa, I.P.S.; da Silva, M.A.N.; Ferreira, V.N.d.S.; de Almeida, A.L.T.; Caldas, G.C.; de Almeida, A.S.; de Filippis, A.M.B.; Fintelman-Rodrigues, N.; Silva, A.d.P.D.d.; et al. In Vitro System for Studying Ilhéus Virus, a Neglected Arbovirus: Ultrastructural Characterization of Cytopathology, Morphology, and Morphogenesis. Viruses 2025, 17, 320. https://doi.org/10.3390/v17030320
Garcia MDdN, Da Costa IPS, da Silva MAN, Ferreira VNdS, de Almeida ALT, Caldas GC, de Almeida AS, de Filippis AMB, Fintelman-Rodrigues N, Silva AdPDd, et al. In Vitro System for Studying Ilhéus Virus, a Neglected Arbovirus: Ultrastructural Characterization of Cytopathology, Morphology, and Morphogenesis. Viruses. 2025; 17(3):320. https://doi.org/10.3390/v17030320
Chicago/Turabian StyleGarcia, Maycon Douglas do Nascimento, Igor Pinto Silva Da Costa, Marcos Alexandre Nunes da Silva, Vivian Neuza dos Santos Ferreira, Ana Luisa Teixeira de Almeida, Gabriela Cardoso Caldas, Andressa Santos de Almeida, Ana Maria Bispo de Filippis, Natalia Fintelman-Rodrigues, Aline de Paula Dias da Silva, and et al. 2025. "In Vitro System for Studying Ilhéus Virus, a Neglected Arbovirus: Ultrastructural Characterization of Cytopathology, Morphology, and Morphogenesis" Viruses 17, no. 3: 320. https://doi.org/10.3390/v17030320
APA StyleGarcia, M. D. d. N., Da Costa, I. P. S., da Silva, M. A. N., Ferreira, V. N. d. S., de Almeida, A. L. T., Caldas, G. C., de Almeida, A. S., de Filippis, A. M. B., Fintelman-Rodrigues, N., Silva, A. d. P. D. d., Alves Ferreira, M., Souza, T. M. L., Pauvolid-Corrêa, A., & Barreto-Vieira, D. F. (2025). In Vitro System for Studying Ilhéus Virus, a Neglected Arbovirus: Ultrastructural Characterization of Cytopathology, Morphology, and Morphogenesis. Viruses, 17(3), 320. https://doi.org/10.3390/v17030320