
Structure and Antigenicity of the Porcine Astrovirus 4 Capsid Spike
 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
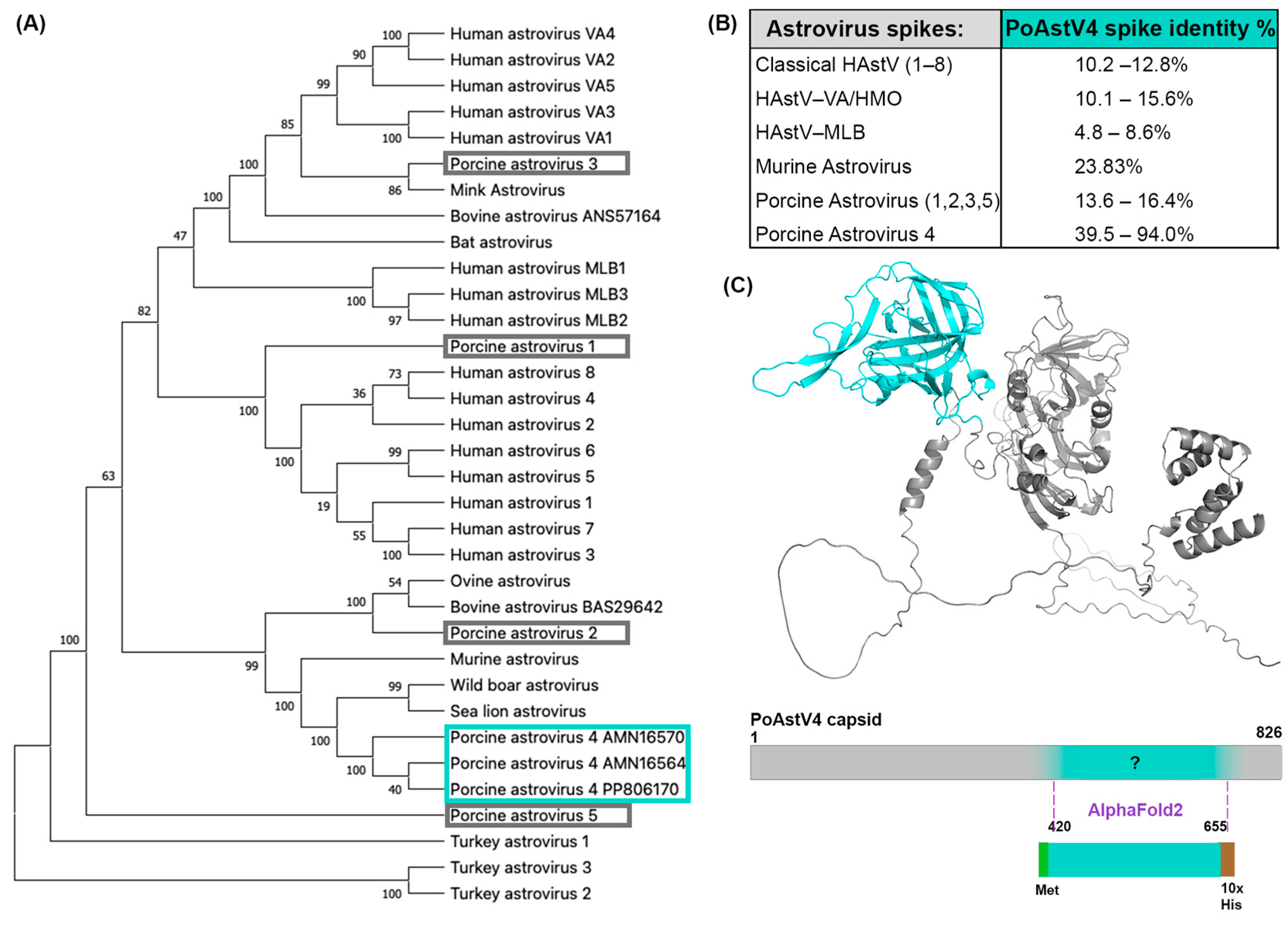
2.1. Phylogenetic Analysis of Astroviruses with MEGA X
2.2. Comparison of Pairwise Identity between PoAstV4 Spike and HAstV Spikes
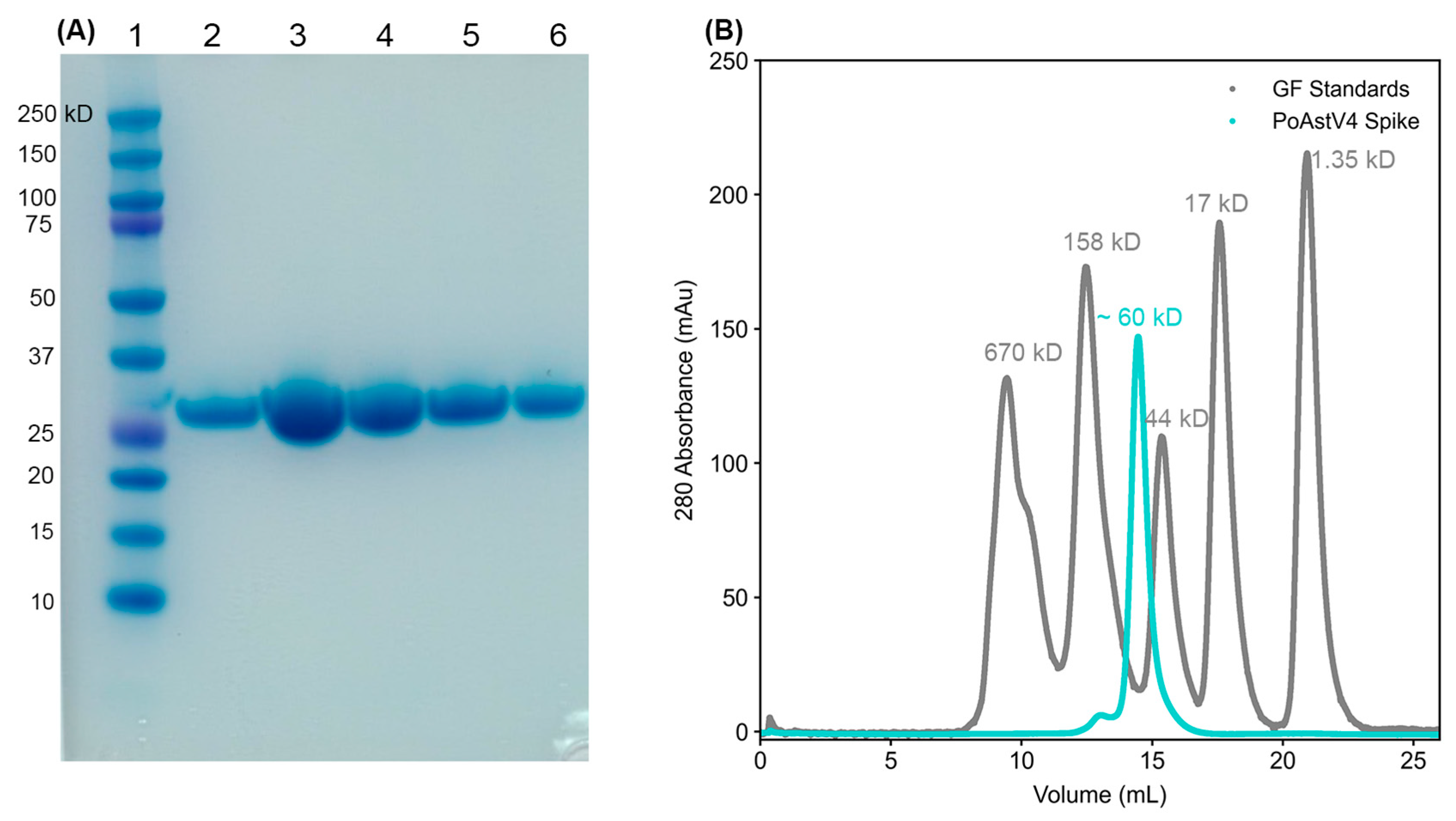
2.3. Design and Production of Recombinant PoAstV4 Capsid Spike Protein
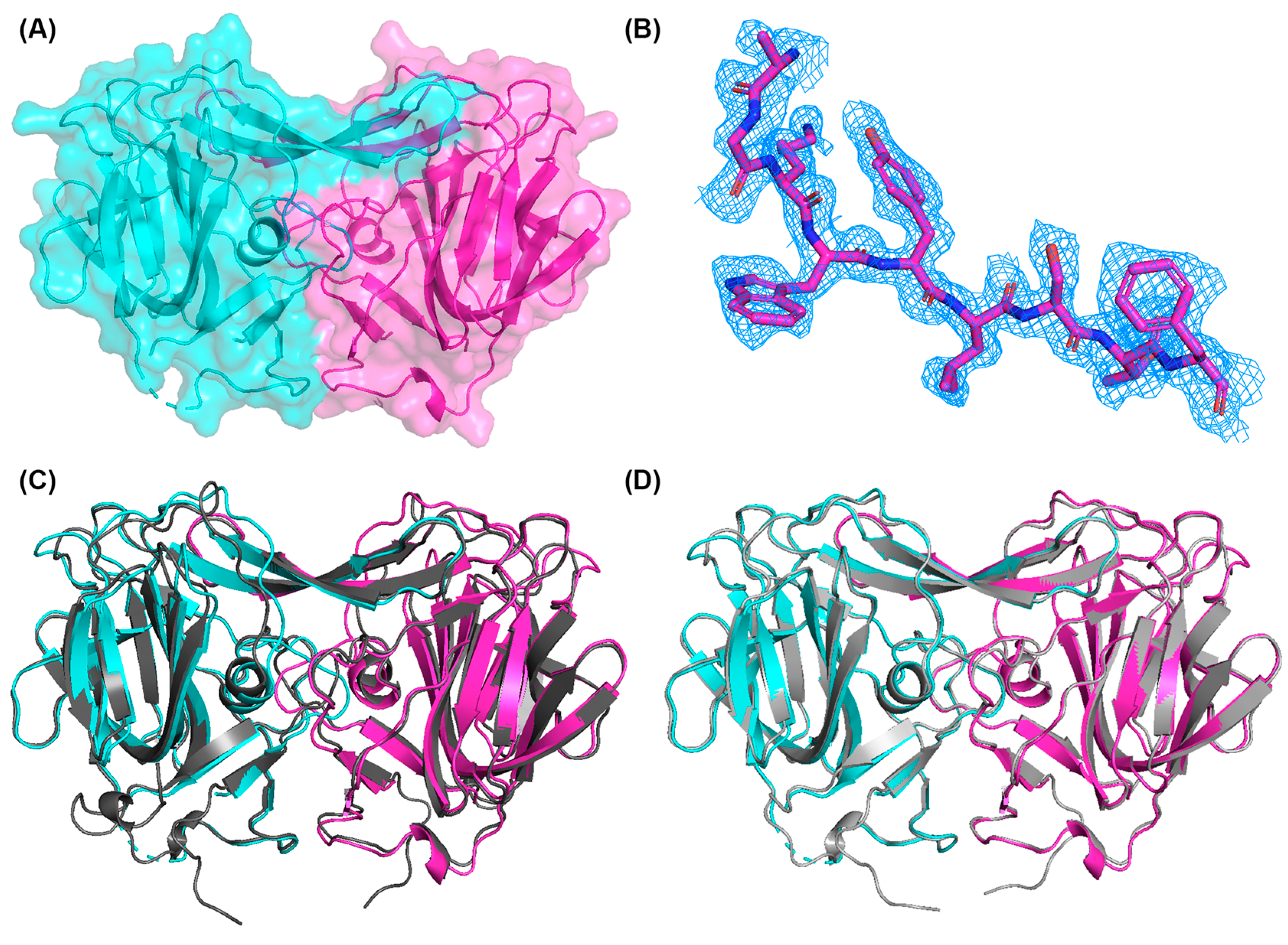
2.4. Structural Determination of PoAstV4 Spike
2.5. Development of an Enzyme-Linked Immunosorbent Assay to Evaluate PoAstV Antibodies in Pig Sera
3. Results
3.1. Delineation and Production of Recombinant PoAstV4 Capsid Spike Domain
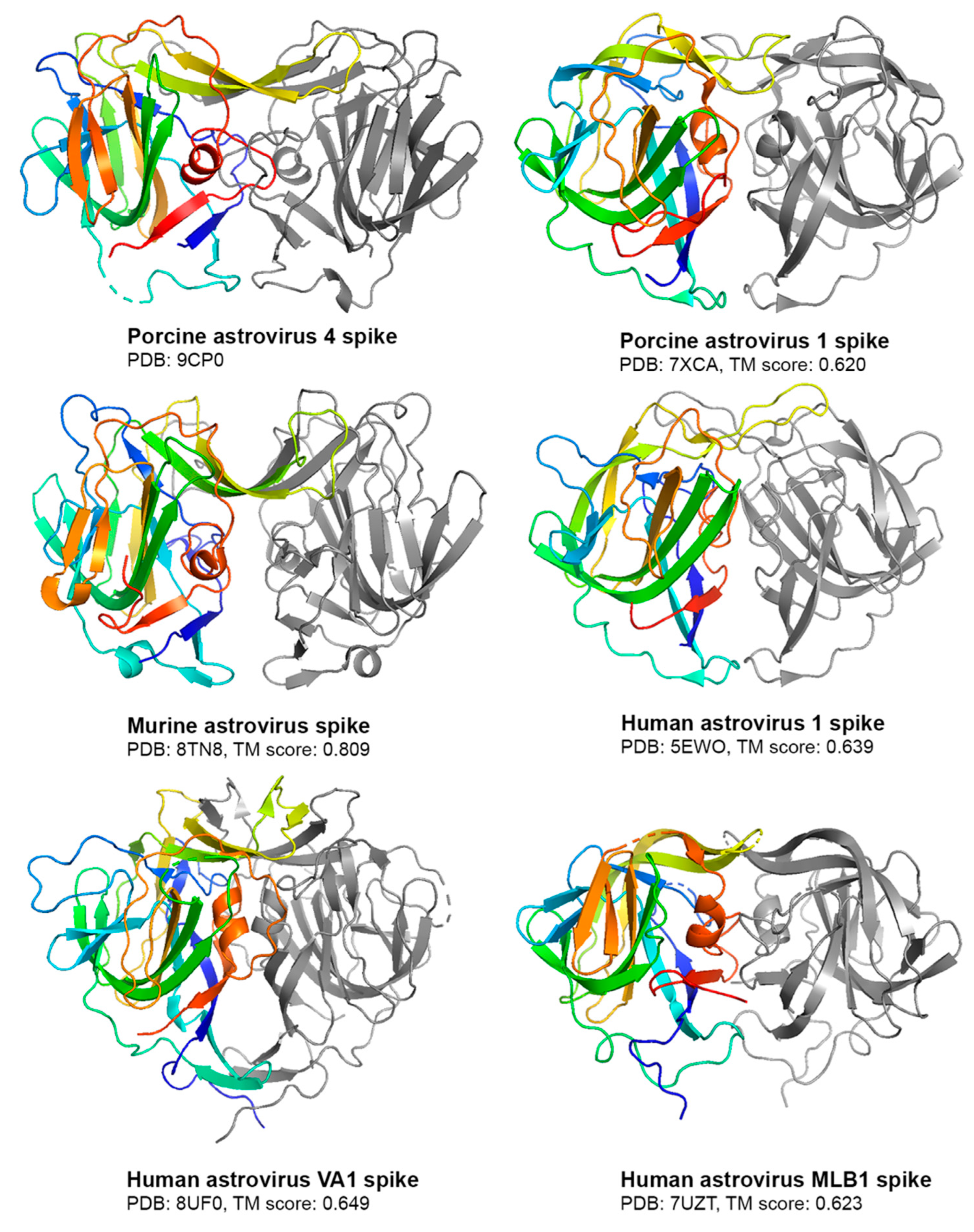
3.2. Crystal Structure of PoAstV4 Spike and Similarity to Other Astroviruses
3.3. Development of an ELISA to Detect PoAstV4 Antibodies in Pig Sera
4. Discussion
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bosch, A.; Pintó, R.M.; Guix, S. Human Astroviruses. Clin. Microbiol. Rev. 2014, 27, 1048–1074. [Google Scholar] [CrossRef] [PubMed]
- Olortegui, M.P.; Rouhani, S.; Yori, P.P.; Salas, M.S.; Trigoso, D.R.; Mondal, D.; Bodhidatta, L.; Platts-Mills, J.; Samie, A.; Kabir, F.; et al. Astrovirus Infection and Diarrhea in 8 Countries. Pediatrics 2018, 141, e20171326. [Google Scholar] [CrossRef] [PubMed]
- Vu, D.-L.; Cordey, S.; Brito, F.; Kaiser, L. Novel human astroviruses: Novel human diseases? J. Clin. Virol. 2016, 82, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Meyer, L.; Delgado-Cunningham, K.; Lorig-Roach, N.; Ford, J.; DuBois, R.M. Human Astrovirus 1–8 Seroprevalence Evaluation in a United States Adult Population. Viruses 2021, 13, 979. [Google Scholar] [CrossRef]
- Koopmans, M.P.G.; Bijen, M.H.L.; Monroe, S.S.; Vinjé, J. Age-Stratified Seroprevalence of Neutralizing Antibodies to Astrovirus Types 1 to 7 in Humans in The Netherlands. Clin. Diagn. Lab. Immunol. 1998, 5, 33–37. [Google Scholar] [CrossRef]
- Payne, S. Family Astroviridae. Viruses 2017, 125–128. [Google Scholar] [CrossRef]
- De Benedictis, P.; Schultz-Cherry, S.; Burnham, A.; Cattoli, G. Astrovirus infections in humans and animals—Molecular biology, genetic diversity, and interspecies transmissions. Infect. Genet. Evol. 2011, 11, 1529–1544. [Google Scholar] [CrossRef]
- Dryden, K.A.; Tihova, M.; Nowotny, N.; Matsui, S.M.; Mendez, E.; Yeager, M. Immature and Mature Human Astrovirus: Structure, Conformational Changes, and Similarities to Hepatitis E Virus. J. Mol. Biol. 2012, 422, 650–658. [Google Scholar] [CrossRef]
- Arias, C.; DuBois, R. The Astrovirus Capsid: A Review. Viruses 2017, 9, 15. [Google Scholar] [CrossRef]
- Ricemeyer, L.; Aguilar-Hernández, N.; López, T.; Espinosa, R.; Lanning, S.; Mukherjee, S.; Cuellar, C.; López, S.; Arias, C.F.; DuBois, R.M. Structures of Two Human Astrovirus Capsid/Neutralizing Antibody Complexes Reveal Distinct Epitopes and Inhibition of Virus Attachment to Cells. J. Virol. 2022, 96, e01415-21. [Google Scholar] [CrossRef]
- Bogdanoff, W.A.; Campos, J.; Perez, E.I.; Yin, L.; Alexander, D.L.; DuBois, R.M. Structure of a Human Astrovirus Capsid-Antibody Complex and Mechanistic Insights into Virus Neutralization. J. Virol. 2017, 91, e01859-16. [Google Scholar] [CrossRef] [PubMed]
- York, R.L.; Yousefi, P.A.; Bogdanoff, W.; Haile, S.; Tripathi, S.; DuBois, R.M. Structural, Mechanistic, and Antigenic Characterization of the Human Astrovirus Capsid. J. Virol. 2016, 90, 2254–2263. [Google Scholar] [CrossRef] [PubMed]
- Lanning, S.; Pedicino, N.; Haley, D.J.; Hernandez, S.; Cortez, V.; DuBois, R.M. Structure and immunogenicity of the murine astrovirus capsid spike. J. Gen. Virol. 2023, 104, 001913. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, R.; López, T.; Bogdanoff, W.A.; Espinoza, M.A.; López, S.; DuBois, R.M.; Arias, C.F. Isolation of Neutralizing Monoclonal Antibodies to Human Astrovirus and Characterization of Virus Variants That Escape Neutralization. J. Virol. 2019, 93, e01465-18. [Google Scholar] [CrossRef]
- Bass, D.M.; Upadhyayula, U. Characterization of human serotype 1 astrovirus-neutralizing epitopes. J. Virol. 1997, 71, 8666–8671. [Google Scholar] [CrossRef]
- Bridger, J.C. Detection by electron microscopy of caliciviruses, astroviruses and rotavirus-like particles in the faeces of piglets with diarrhoea. Vet. Rec. 1980, 107, 532–533. [Google Scholar]
- Padmanabhan, A.; Hause, B.M. Detection and characterization of a novel genotype of porcine astrovirus 4 from nasal swabs from pigs with acute respiratory disease. Arch. Virol. 2016, 161, 2575–2579. [Google Scholar] [CrossRef]
- Rawal, G.; Linhares, D.C.L. Scoping review on the epidemiology, diagnostics and clinical significance of porcine astroviruses. Transbounding Emerg. Dis. 2022, 69, 974–985. [Google Scholar] [CrossRef]
- Fang, Q.; Wang, C.; Liu, H.; Wu, Q.; Liang, S.; Cen, M.; Dong, Q.; Wei, Y.; Chen, Y.; Ouyang, K.; et al. Pathogenic Characteristics of a Porcine Astrovirus Strain Isolated in China. Viruses 2019, 11, 1156. [Google Scholar] [CrossRef]
- Xiao, C.-T.; Giménez-Lirola, L.G.; Gerber, P.F.; Jiang, Y.-H.; Halbur, P.G.; Opriessnig, T. Identification and characterization of novel porcine astroviruses (PAstVs) with high prevalence and frequent co-infection of individual pigs with multiple PAstV types. J. Gen. Virol. 2013, 94, 570–582. [Google Scholar] [CrossRef]
- Luo, Z.; Roi, S.; Dastor, M.; Gallice, E.; Laurin, M.-A.; L’Homme, Y. Multiple novel and prevalent astroviruses in pigs. Vet. Microbiol. 2011, 149, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Flores, C.; Ariyama, N.; Bennett, B.; Mena, J.; Verdugo, C.; Mor, S.; Brito, B.; Ramírez-Toloza, G.; Neira, V. Case Report: First Report and Phylogenetic Analysis of Porcine Astroviruses in Chile. Front. Vet. Sci. 2021, 8, 76483. [Google Scholar] [CrossRef] [PubMed]
- Geyer, A.; Steele, A.D.; Peenze, I.; Lecatsas, G. Astrovirus-like particles, adenoviruses and rotaviruses associated with diarrhoea in piglets. J. S. Afr. Vet. Assoc. 1994, 65, 164–166. [Google Scholar] [PubMed]
- Ulloa, J.C.; Gutiérrez, M.F. Genomic analysis of two ORF2 segments of new porcine astrovirus isolates and their close relationship with human astroviruses. Can. J. Microbiol. 2010, 56, 569–577. [Google Scholar] [CrossRef]
- Rahe, M.C.; Michael, A.; Piñeyro, P.E.; Groeltz-Thrush, J.; Derscheid, R.J. Porcine Astrovirus 4 Detection in Lesions of Epitheliotropic Viral Infection in the Porcine Respiratory Tract. Transbound. Emerg. Dis. 2023, 2023, 9113355. [Google Scholar] [CrossRef]
- Opriessnig, T.; Xiao, C.-T.; Halbur, P.G. Porcine Astrovirus Type 5-Associated Enteritis in Pigs. J. Comp. Pathol. 2020, 181, 38–46. [Google Scholar] [CrossRef]
- Arruda, B.; Arruda, P.; Hensch, M.; Chen, Q.; Zheng, Y.; Yang, C.; Gatto, I.R.H.; Ferreyra, F.M.; Gauger, P.; Schwartz, K.; et al. Porcine Astrovirus Type 3 in Central Nervous System of Swine with Polioencephalomyelitis. Emerg. Infect. Dis. 2017, 23, 2097–2100. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The Rapid Generation of Mutation Data Matrices from Protein Sequences. Bioinformatics 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Abramson, J.; Adler, J.; Dunger, J.; Evans, R.; Green, T.; Pritzel, A.; Ronneberger, O.; Willmore, L.; Ballard, A.J.; Bambrick, J.; et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 2024, 630, 493–500. [Google Scholar] [CrossRef]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making protein folding accessible to all. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and Development of Coot. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [PubMed]
- Liebschner, D.; Afonine, P.V.; Baker, M.L.; Bunkóczi, G.; Chen, V.B.; Croll, T.I.; Hintze, B.; Hung, L.-W.; Jain, S.; McCoy, A.J.; et al. Macromolecular Structure Determination Using X-rays, Neutrons and Electrons: Recent Developments in Phenix. Acta Crystallogr. Sect. D Struct. Biol. 2019, 75, 861–877. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Cooper, D.R.; Porebski, P.J.; Shabalin, I.G.; Handing, K.B.; Minor, W. CheckMyMetal: A macromolecular metal-binding validation tool. Acta Crystallogr. Sect. D 2017, 73, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Welch, M.; Park, J.; Harmon, K.; Zhang, J.; Piñeyro, P.; Giménez-Lirola, L.; Zhang, M.; Wang, C.; Patterson, A.; Gauger, P.C. Pathogenesis of a novel porcine parainfluenza virus type 1 isolate in conventional and colostrum deprived/caesarean derived pigs. Virology 2021, 563, 88–97. [Google Scholar] [CrossRef]
- Zhang, Y.; Skolnick, J. TM-Align: A Protein Structure Alignment Algorithm Based on the TM-Score. Nucleic Acids Res. 2005, 33, 2302–2309. [Google Scholar] [CrossRef]
- Pan, L.X.; Zhang, W.C.; Yang, D.F.; Yang, L.Y.; Liu, H. Crystal structure of the porcine astrovirus capsid spike domain. 2023, unpublished. [CrossRef]
- Ghosh, A.; Delgado-Cunningham, K.; López, T.; Green, K.; Arias, C.F.; DuBois, R.M. Structure and Antigenicity of the Divergent Human Astrovirus VA1 Capsid Spike. PLoS Pathog. 2024, 20, e1012028. [Google Scholar] [CrossRef]
- Delgado-Cunningham, K.; López, T.; Khatib, F.; Arias, C.F.; DuBois, R.M. Structure of the Divergent Human Astrovirus MLB Capsid Spike. Structure 2022, 30, 1573–1581.e3. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PoAstV4 Spike | |
|---|---|
| PDB entry: | 9CP0 |
| Data collection: | |
| Wavelength (Å) | 0.97 |
| Space Group | P1 |
| Cell dimensions: | |
| a, b, c (Å) | 46.39, 66.21, 87.75 |
| α, β, ɣ (°) | 74.54, 81.67, 78.61 |
| Resolution (Å) | 38.33–1.85 (1.91–1.85) * |
| Rmerge | 0.144 (0.474) |
| I/σl | 8.1 (2.1) |
| Completeness | 96.1% (93.9%) |
| Multiplicity | 3.2 (3.4) |
| CC1/2 | 0.984 (0.814) |
| Refinement: | |
| Resolution | 39.78–1.85 |
| No. reflections for refinement | 80,308 |
| No. reflections for Rfree | 1998 |
| Rwork/Rfree | 0.316/0.362 |
| No. atoms | 7412 |
| Protein | 7128 |
| Ligand/ion | 4 |
| Water | 280 |
| B factors (Å2): | 15.29 |
| Protein | 15.30 |
| Ligand/ion | 16.29 |
| Water | 14.99 |
| RMSD: | |
| Bond lengths (Å) | 0.005 |
| Bond angles (°) | 0.74 |
| Ramachandran statistics: | |
| Favored (%) | 96.77 |
| Allowed (%) | 3.23 |
| Outliers (%) | 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haley, D.J.; Lanning, S.; Henricson, K.E.; Mardirossian, A.A.; Cirillo, I.; Rahe, M.C.; DuBois, R.M. Structure and Antigenicity of the Porcine Astrovirus 4 Capsid Spike. Viruses 2024, 16, 1596. https://doi.org/10.3390/v16101596
Haley DJ, Lanning S, Henricson KE, Mardirossian AA, Cirillo I, Rahe MC, DuBois RM. Structure and Antigenicity of the Porcine Astrovirus 4 Capsid Spike. Viruses. 2024; 16(10):1596. https://doi.org/10.3390/v16101596
Chicago/Turabian StyleHaley, Danielle J., Sarah Lanning, Kyle E. Henricson, Andre A. Mardirossian, Iyan Cirillo, Michael C. Rahe, and Rebecca M. DuBois. 2024. "Structure and Antigenicity of the Porcine Astrovirus 4 Capsid Spike" Viruses 16, no. 10: 1596. https://doi.org/10.3390/v16101596
APA StyleHaley, D. J., Lanning, S., Henricson, K. E., Mardirossian, A. A., Cirillo, I., Rahe, M. C., & DuBois, R. M. (2024). Structure and Antigenicity of the Porcine Astrovirus 4 Capsid Spike. Viruses, 16(10), 1596. https://doi.org/10.3390/v16101596