

Developments in Exploring Fungal Secondary Metabolites as Antiviral Compounds and Advances in HIV-1 Inhibitor Screening Assays


Abstract
1. Introduction
2. The Biology of HIV and Currently Available Therapies for HIV Treatment
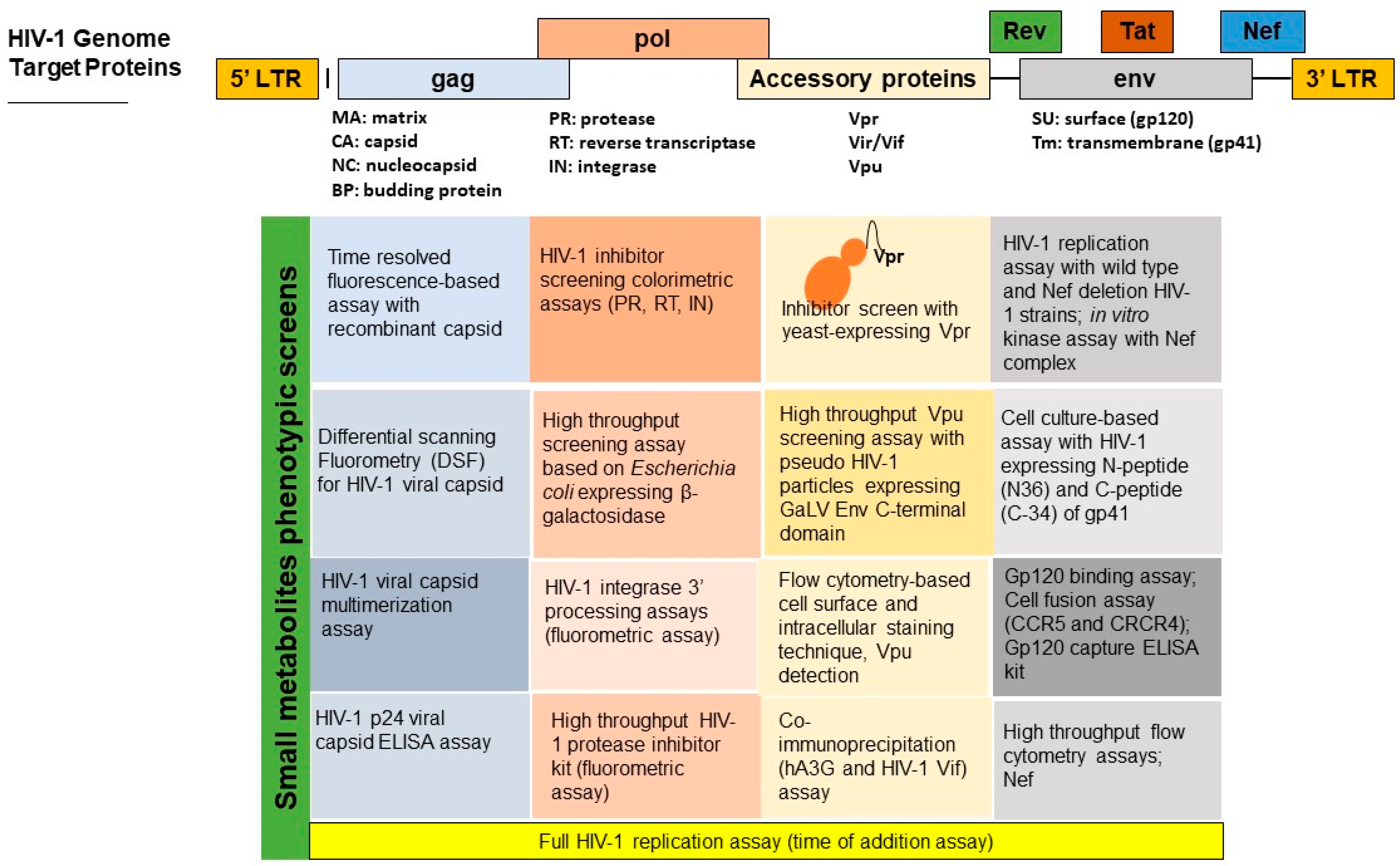
3. Advances in Antiviral Screening Assays
4. Advances in Developing Fungal-Derived Small Molecular Inhibitors for Major HIV-1 Target Sites
5. Fungal Compounds as HIV-1 Cell-Surface Receptor Attachment Inhibitors
6. Fungal Compounds as Reverse Transcriptase Inhibitors
7. Fungal Compounds as Integrase Inhibitors
8. Fungal Compounds as Protease Inhibitors
9. Advances in Developing Fungal-Derived Small Molecular Inhibitors for New HIV-1 Target Sites

| HIV-1 Life Cycle Stages | HIV-1 Agent/Compound | Structure | Fungal Species | Mechanism of Action | Activity Concentration | Reference |
|---|---|---|---|---|---|---|
| Fusion or entry inhibitors | Isochromophilones I C23H25O5Cl Mw 416.9 |  | Penicillium multicolor FO-2338 | Inhibited the gp120, CD4 binding | 6.6 μM | [85,86,122] |
| Isochromophilones II C22H27O4Cl Mw 390.9 |  | 3.9 µM | ||||
| Variecolin C25H36O2l Mw 368.55 |  | Emericella aurantiobrumea | Competed with macrophage inflammatory protein (MIP)-1 for binding to CCR5 | 9 µM | [87,122,123] | |
| Novel Sch 210972 C25H35NO6 Mw 445.55 |  | Chaetomium globosum Kunze SCF 1705 | Strongly inhibited CCR-5 inhibitory activity | 79 nM | [83,120,122,123] | |
| Penicillixanthone A C32H30O14 Mw 638.578 |  | Aspergillus fumigatus | Inhibited CCR5-tropic HIV-1 SF162 | 0.36 μM | [123,124] | |
| Inhibited CXCR4-tropic HIV-1 NL4-3 | 0.26 μM | |||||
| Reverse Transcription | Stachybotrin D C26H35NO5 Mw 441.56 |  | Stachybotrys chartarum | Inhibited the RT RNA-dependent DNA polymerase activity in a dose-dependent manner | 8.4 µM | [122,125] |
| Helotialins A C23H30O6 Mw 402.48 |  | Helotialean Ascomycete | Not identified | 8.01 nM | [122,126] | |
| Helotialins B C22H30O4 Mw 358.47 |  | 27.9 nM | ||||
| Altertoxin Ⅴ C20H14O6 Mw 350.32 |  | Alternaria tenuissima | Not identified | ≤2.20 µM | [90,122] | |
| Integration | Xanthoviridicatins E and F C27H20O9 Mw 488.44 |  | Penicillium chrysogenum | They inhibited the cleavage reaction of HIV-1 integrase. | 6 and 5 μM | [93,122] |
| Aquastatin A C36H52O12 MW 676.79 |  | Fusarium aquaeductuum | Moderately inhibited the strand transfer reaction of HIV-1 integrase | 50 µM | [122,127] | |
| Integracins B C35H54O7 Mw 586.8 |  | Cytonaema sp. | Inhibited coupled reaction of recombinant HIV-1 integrase | 6.1 µM | [122,128] | |
| Integric acid C25H52O6 Mw 430.53 |  | Xylaria feejeensis | Inhibited the 3′ end processing, strand transfer and disintegration | 5–10 μM | [122,129] | |
| Atrovenetinone methyl acetal C20H20O7 Mw 372.37 |  | Penicillium sp. FKI-1463 | Not identified | 19 µM | [97,122] | |
| Translation or budding | Hinnuliquinone C32H30N2O4 Mw 506.59 |  | Unidentified fungus | Not identified | 2.5 µM. | [100,122] |
| Mulberroside C C24H26O9 Mw 458.46 |  | Unidentified fungus | Inhibited HIV-1 protease in a dose-dependent manner | <7.8 µg/mL | [56,122] |
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data availability statement
Conflicts of Interest
Abbreviations
References
- Global HIV & AIDS statistics. Available online: https://www.unaids.org/en/resources/fact-sheet (accessed on 13 April 2023).
- Arts, E.J.; Hazuda, D.J. HIV-1 antiretroviral drug therapy. Cold Spring Harb. Perspect. Med. 2012, 2, a007161. [Google Scholar] [CrossRef]
- Andersen, R.J.; Ntie-Kang, F.; Tietjen, I. Natural product-derived compounds in HIV suppression, remission, and eradication strategies. Antivir. Res. 2018, 158, 63–77. [Google Scholar] [CrossRef]
- Reed, J.C.; Solas, D.; Kitaygorodskyy, A.; Freeman, B.; Ressler, D.T.B.; Phuong, D.J.; Swain, J.V.; Matlack, K.; Hurt, C.R.; Lingappa, V.R.; et al. Identification of an Antiretroviral Small Molecule That Appears to Be a Host-Targeting Inhibitor of HIV-1 Assembly. J. Virol. 2021, 95, e00883-20. [Google Scholar] [CrossRef] [PubMed]
- Andrae-Marobela, K.; Ghislain, F.W.; Okatch, H.; Majinda, R.R. Polyphenols: A diverse class of multi-target anti-HIV-1 agents. Curr. Drug Metab. 2013, 14, 392–413. [Google Scholar] [CrossRef] [PubMed]
- Farooq, T.; Hameed, A.; Rehman, K.; Ibrahim, M.; Qadir, M.I.; Akash, M.S. Antiretroviral Agents: Looking for the Best Possible Chemotherapeutic Options to Conquer HIV. Crit. Rev. Eukaryot. Gene Expr. 2016, 26, 363–381. [Google Scholar] [CrossRef]
- Joly, V.; Flandre, P.; Meiffredy, V.; Leturque, N.; Harel, M.; Aboulker, J.P.; Yeni, P. Increased risk of lipoatrophy under stavudine in HIV-1-infected patients: Results of a substudy from a comparative trial. Aids 2002, 16, 2447–2454. [Google Scholar] [CrossRef] [PubMed]
- Povar-Echeverría, M.; Comet-Bernad, M.; Gasso-Sánchez, A.; Ger-Buil, A.; Navarro-Aznarez, H.; Martínez-Álvarez, R.; Arazo-Garcés, P. Neuropsychiatric adverse effects of dolutegravir in real-life clinical practice. Enferm. Infecc. Microbiol. Clin. 2021, 39, 78–82. [Google Scholar] [CrossRef]
- Desai, M.; Iyer, G.; Dikshit, R.K. Antiretroviral drugs: Critical issues and recent advances. Indian J. Pharmacol. 2012, 44, 288–298. [Google Scholar] [CrossRef]
- Alexaki, A.; Liu, Y.; Wigdahl, B. Cellular reservoirs of HIV-1 and their role in viral persistence. Curr. HIV Res. 2008, 6, 388–400. [Google Scholar] [CrossRef]
- Siliciano, J.D.; Siliciano, R.F. A long-term latent reservoir for HIV-1: Discovery and clinical implications. J. Antimicrob. Chemother. 2004, 54, 6–9. [Google Scholar] [CrossRef]
- Sharapov, A.D.; Fatykhov, R.F.; Khalymbadzha, I.A.; Zyryanov, G.V.; Chupakhin, O.N.; Tsurkan, M.V. Plant Coumarins with Anti-HIV Activity: Isolation and Mechanisms of Action. Int. J. Mol. Sci. 2023, 24, 2839. [Google Scholar] [CrossRef]
- Gunatilaka, A.A. Natural products from plant-associated microorganisms: Distribution, structural diversity, bioactivity, and implications of their occurrence. J. Nat. Prod. 2006, 69, 509–526. [Google Scholar] [CrossRef]
- Hashemi, P.; Sadowski, I. Diversity of small molecule HIV-1 latency reversing agents identified in low- and high-throughput small molecule screens. Med. Res. Rev. 2020, 40, 881–908. [Google Scholar] [CrossRef]
- Linnakoski, R.; Reshamwala, D.; Veteli, P.; Cortina-Escribano, M.; Vanhanen, H.; Marjomäki, V. Antiviral Agents from Fungi: Diversity, Mechanisms and Potential Applications. Front. Microbiol. 2018, 9, 2325. [Google Scholar] [CrossRef] [PubMed]
- Roy, B.G. Potential of small-molecule fungal metabolites in antiviral chemotherapy. Antivir. Chem. Chemother. 2017, 25, 20–52. [Google Scholar] [CrossRef] [PubMed]
- Weiss, R.A.; Dalgleish, A.G.; Loveday, C.; Pillay, D. Immunodeficiency Viruses. In Principles and Practice of Clinical Virology; John Wiley and Sons: Hoboken, NJ, USA, 2004; pp. 721–757. [Google Scholar]
- Bhatti, A.B.; Usman, M.; Kandi, V. Current Scenario of HIV/AIDS, Treatment Options, and Major Challenges with Compliance to Antiretroviral Therapy. Cureus 2016, 8, e515. [Google Scholar] [CrossRef]
- Kapila, A.; Chaudhary, S.; Sharma, R.B.; Vashist, H.; Sisodia, S.S.; Gupta, A. A REVIEW on: HIV AIDS. Indian J. Pharm. Biol. Res. 2016, 4, 69–73. [Google Scholar] [CrossRef]
- Singh, I.P.; Bharate, S.; Bhutani, K. Anti-HIV natural products. Curr. Sci. 2005, 89, 269–290. [Google Scholar]
- Mesters, J.R.; Tan, J.; Hilgenfeld, R. Viral enzymes. Curr. Opin. Struct. Biol. 2006, 16, 776–786. [Google Scholar] [CrossRef] [PubMed]
- Sundquist, W.I.; Kräusslich, H.G. HIV-1 assembly, budding, and maturation. Cold Spring Harb. Perspect Med. 2012, 2, a006924. [Google Scholar] [CrossRef]
- Li, L.; Li, H.S.; Pauza, C.D.; Bukrinsky, M.; Zhao, R.Y. Roles of HIV-1 auxiliary proteins in viral pathogenesis and host-pathogen interactions. Cell Res. 2005, 15, 923–934. [Google Scholar] [CrossRef]
- Rheinemann, L.; Sundquist, W.I. Virus. Budding. In Encyclopedia of Virology; Academic Press: Cambridge, MA, USA, 2021; pp. 519–528. [Google Scholar] [CrossRef]
- Kessl, J.J.; Kutluay, S.B.; Townsend, D.; Rebensburg, S.; Slaughter, A.; Larue, R.C.; Shkriabai, N.; Bakouche, N.; Fuchs, J.R.; Bieniasz, P.D.; et al. HIV-1 Integrase Binds the Viral RNA Genome and Is Essential during Virion Morphogenesis. Cell 2016, 166, 1257–1268. [Google Scholar] [CrossRef] [PubMed]
- Piot, P.; Abdool Karim, S.S.; Hecht, R.; Legido-Quigley, H.; Buse, K.; Stover, J.; Resch, S.; Ryckman, T.; Møgedal, S.; Dybul, M.; et al. Defeating AIDS—Advancing global health. Lancet 2015, 386, 171–218. [Google Scholar] [CrossRef]
- Broder, S. The development of antiretroviral therapy and its impact on the HIV-1/AIDS pandemic. Antivir. Res. 2010, 85, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Serna-Arbeláez, M.S.; Florez-Sampedro, L.; Orozco, L.P.; Ramírez, K.; Galeano, E.; Zapata, W. Natural Products with Inhibitory Activity against Human Immunodeficiency Virus Type 1. Adv. Virol. 2021, 2021, 5552088. [Google Scholar] [CrossRef]
- Painter, G.R.; Almond, M.R.; Mao, S.; Liotta, D.C. Biochemical and mechanistic basis for the activity of nucleoside analogue inhibitors of HIV reverse transcriptase. Curr. Top Med. Chem. 2004, 4, 1035–1044. [Google Scholar] [CrossRef]
- Blanco, J.L.; Whitlock, G.; Milinkovic, A.; Moyle, G. HIV integrase inhibitors: A new era in the treatment of HIV. Expert Opin. Pharmacother. 2015, 16, 1313–1324. [Google Scholar] [CrossRef] [PubMed]
- Konvalinka, J.; Kräusslich, H.G.; Müller, B. Retroviral proteases and their roles in virion maturation. Virology 2015, 479–480, 403–417. [Google Scholar] [CrossRef]
- Peters, H.; Francis, K.; Sconza, R.; Horn, A.; Peckham, C.S.; Tookey, P.A.; Thorne, C. UK Mother-to-Child HIV Transmission Rates Continue to Decline: 2012–2014. Clin. Infect. Dis. 2017, 64, 527–528. [Google Scholar] [CrossRef]
- Fantauzzi, A.; Mezzaroma, I. Dolutegravir: Clinical efficacy and role in HIV therapy. Ther. Adv. Chronic Dis. 2014, 5, 164–177. [Google Scholar] [CrossRef]
- Cutrell, J.; Bedimo, R. Single-Tablet Regimens in the Treatment of HIV-1 Infection. Fed. Pract. 2016, 33, 24s–30s. [Google Scholar]
- Günthard, H.F.; Saag, M.S.; Benson, C.A.; del Rio, C.; Eron, J.J.; Gallant, J.E.; Hoy, J.F.; Mugavero, M.J.; Sax, P.E.; Thompson, M.A.; et al. Antiretroviral Drugs for Treatment and Prevention of HIV Infection in Adults: 2016 Recommendations of the International Antiviral Society-USA Panel. JAMA 2016, 316, 191–210. [Google Scholar] [CrossRef] [PubMed]
- Eggleton, J.S.; Nagalli, S. Highly Active Antiretroviral Therapy (HAART). In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2023. [Google Scholar]
- Montessori, V.; Press, N.; Harris, M.; Akagi, L.; Montaner, J.S. Adverse effects of antiretroviral therapy for HIV infection. Cmaj 2004, 170, 229–238. [Google Scholar] [PubMed]
- Margolis, A.M.; Heverling, H.; Pham, P.A.; Stolbach, A. A review of the toxicity of HIV medications. J. Med. Toxicol. 2014, 10, 26–39. [Google Scholar] [CrossRef]
- Das, K.; Arnold, E. HIV-1 reverse transcriptase and antiviral drug resistance. Part 2. Curr. Opin. Virol. 2013, 3, 119–128. [Google Scholar] [CrossRef]
- Fobofou, S.A.T.; Franke, K.; Brandt, W.; Manzin, A.; Madeddu, S.; Serreli, G.; Sanna, G.; Wessjohann, L.A. Bichromonol, a dimeric coumarin with anti-HIV activity from the stem bark of Hypericum roeperianum. Nat. Prod. Res. 2022, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Miethke, M.; Pieroni, M.; Weber, T.; Brönstrup, M.; Hammann, P.; Halby, L.; Arimondo, P.B.; Glaser, P.; Aigle, B.; Bode, H.B.; et al. Towards the sustainable discovery and development of new antibiotics. Nat. Rev. Chem. 2021, 5, 726–749. [Google Scholar] [CrossRef]
- Richman, D.D.; Nathanson, N. Chapter 20—Antiviral Therapy. In Viral Pathogenesis, 3rd ed.; Katze, M.G., Korth, M.J., Law, G.L., Nathanson, N., Eds.; Academic Press: Boston, MA, USA, 2016; pp. 271–287. [Google Scholar]
- Payne, S. Methods to Study Viruses. In Viruses; Elsevier Inc.: Houston, TX, USA, 2017; pp. 37–52. [Google Scholar] [CrossRef]
- Hudson, J.B.; Graham, E.A.; Towers, G.H. Antiviral assays on phytochemicals: The influence of reaction parameters. Planta Med. 1994, 60, 329–332. [Google Scholar] [CrossRef]
- Cao, J.; Isaacson, J.; Patick, A.K.; Blair, W.S. High-throughput human immunodeficiency virus type 1 (HIV-1) full replication assay that includes HIV-1 Vif as an antiviral target. Antimicrob. Agents Chemother. 2005, 49, 3833–3841. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.A.; Lyddon, T.D.; Gil, H.M.; Evans, D.T.; Kuzmichev, Y.V.; Richard, J.; Finzi, A.; Welbourn, S.; Rasmussen, L.; Nebane, N.M.; et al. Novel Compound Inhibitors of HIV-1NL4-3 Vpu. Viruses 2022, 14, 817. [Google Scholar] [CrossRef]
- Daelemans, D.; Pauwels, R.; De Clercq, E.; Pannecouque, C. A time-of-drug addition approach to target identification of antiviral compounds. Nat. Protoc. 2011, 6, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Adamson, C.S.; Chibale, K.; Goss, R.J.M.; Jaspars, M.; Newman, D.J.; Dorrington, R.A. Antiviral drug discovery: Preparing for the next pandemic. Chem. Soc. Rev. 2021, 50, 3647–3655. [Google Scholar] [CrossRef] [PubMed]
- Nzimande, B.; Kumalo, H.M.; Ndlovu, S.I.; Mkhwanazi, N.P. Secondary metabolites produced by endophytic fungi, Alternaria alternata, as potential inhibitors of the human immunodeficiency virus. Front. Genet. 2022, 13, 13.1–14. [Google Scholar] [CrossRef]
- McMahon, M.A.; Shen, L.; Siliciano, R.F. New approaches for quantitating the inhibition of HIV-1 replication by antiviral drugs in vitro and in vivo. Curr. Opin. Infect. Dis. 2009, 22, 574–582. [Google Scholar] [CrossRef]
- Shen, L.; Peterson, S.; Sedaghat, A.R.; McMahon, M.A.; Callender, M.; Zhang, H.; Zhou, Y.; Pitt, E.; Anderson, K.S.; Acosta, E.P.; et al. Dose-response curve slope sets class-specific limits on inhibitory potential of anti-HIV drugs. Nat. Med. 2008, 14, 762–766. [Google Scholar] [CrossRef]
- Banning, C.; Votteler, J.; Hoffmann, D.; Koppensteiner, H.; Warmer, M.; Reimer, R.; Kirchhoff, F.; Schubert, U.; Hauber, J.; Schindler, M. A Flow Cytometry-Based FRET Assay to Identify and Analyse Protein-Protein Interactions in Living Cells. PLoS ONE 2010, 5, e9344. [Google Scholar] [CrossRef]
- Arhel, N.; Kirchhoff, F. Host proteins involved in HIV infection: New therapeutic targets. Biochim. Biophys. Acta 2010, 1802, 313–321. [Google Scholar] [CrossRef]
- Passaes, C.; Delagreverie, H.M.; Avettand-Fenoel, V.; David, A.; Monceaux, V.; Essat, A.; Müller-Trutwin, M.; Duffy, D.; De Castro, N.; Wittkop, L.; et al. Ultrasensitive Detection of p24 in Plasma Samples from People with Primary and Chronic HIV-1 Infection. J. Virol. 2021, 95, e0001621. [Google Scholar] [CrossRef] [PubMed]
- Gray, E.R.; Bain, R.; Varsaneux, O.; Peeling, R.W.; Stevens, M.M.; McKendry, R.A. A landscape review of antigen detection for early HIV diagnosis. Aids 2018, 32, 2089–2102. [Google Scholar] [CrossRef]
- Vora, J.; Velhal, S.; Sinha, S.; Patel, V.; Shrivastava, N. Bioactive phytocompound mulberroside C and endophytes of Morus alba as potential inhibitors of HIV-1 replication: A mechanistic evaluation. HIV Med. 2021, 22, 690–704. [Google Scholar] [CrossRef]
- Cao, S.; Woodrow, K.A. Nanotechnology approaches to eradicating HIV reservoirs. Eur. J. Pharm. Biopharm. 2019, 138, 48–63. [Google Scholar] [CrossRef]
- Sinha, C.; Nischal, A.; Bandaru, S.; Kasera, P.; Rajput, A.; Nayarisseri, A.; Khattri, S. An in silico approach for identification of novel inhibitors as a potential therapeutics targeting HIV-1 viral infectivity factor. Curr. Top Med. Chem. 2015, 15, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; De Clercq, E. Overview of Antiviral Drug Discovery and Development: ViralVersusHost Targets. In Antiviral Discovery for Highly Pathogenic Emerging Viruses; Royal Society of Chemistry: Piccadilly, Australia, 2021. [Google Scholar]
- Zhang, D.W.; Xie, L.; Xu, X.S.; Li, Y.; Xu, X. A Broad-Spectrum Antiviral Molecule, Protoporphyrin IX, Acts as a Moderator of HIV-1 Capsid Assembly by Targeting the Capsid Hexamer. Microbiol. Spectr. 2023, 11, e0266322. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zhang, Z.; Liu, Z.; Pan, Q.; Wang, J.; Li, X.; Guo, F.; Liang, C.; Hu, L.; Zhou, J.; et al. Identification of small molecule compounds targeting the interaction of HIV-1 Vif and human APOBEC3G by virtual screening and biological evaluation. Sci. Rep. 2018, 8, 8067. [Google Scholar] [CrossRef]
- Meng, J.; Lai, M.-T.; Munshi, V.; Grobler, J.; McCauley, J.; Zuck, P.; Johnson, E.N.; Uebele, V.N.; Hermes, J.D.; Adam, G.C. Screening of HIV-1 protease using a combination of an ultra-high-throughput fluorescent-based assay and RapidFire mass spectrometry. J. Biomol. Screen. 2015, 20, 606–615. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, Y.; Doi, N.; Koma, T.; Adachi, A.; Nomaguchi, M. Novel In Vitro Screening System Based on Differential Scanning Fluorimetry to Search for Small Molecules against the Disassembly or Assembly of HIV-1 Capsid Protein. Front. Microbiol. 2017, 8, 1413. [Google Scholar] [CrossRef] [PubMed]
- Eglen, M.R.; Singh, R. Galactosidase Enzyme Fragment Complementation as A Novel Technology for High Throughput Screening. Comb. Chem. High Throughput Screen. 2003, 6, 381–387. [Google Scholar] [CrossRef]
- Ding, J.; Zhao, J.; Yang, Z.; Ma, L.; Mi, Z.; Wu, Y.; Guo, J.; Zhou, J.; Li, X.; Guo, Y.; et al. Microbial Natural Product Alternariol 5-O-Methyl Ether Inhibits HIV-1 Integration by Blocking Nuclear Import of the Pre-Integration Complex. Viruses 2017, 9, 105. [Google Scholar] [CrossRef] [PubMed]
- Sillapachaiyaporn, C.; Chuchawankul, S. HIV-1 protease and reverse transcriptase inhibition by tiger milk mushroom (Lignosus rhinocerus) sclerotium extracts: In vitro and in silico studies. J. Tradit. Complement Med. 2020, 10, 396–404. [Google Scholar] [CrossRef]
- Wang, X.; Wang, J.; Zhang, W.; Li, B.; Zhu, Y.; Hu, Q.; Yang, Y.; Zhang, X.; Yan, H.; Zeng, Y. Inhibition of Human Immunodeficiency Virus Type 1 Entry by a Keggin Polyoxometalate. Viruses 2018, 10, 265. [Google Scholar] [CrossRef]
- Emert-Sedlak, L.; Kodama, T.; Lerner, E.C.; Dai, W.; Foster, C.; Day, B.W.; Lazo, J.S.; Smithgall, T.E. Chemical library screens targeting an HIV-1 accessory factor/host cell kinase complex identify novel antiretroviral compounds. ACS Chem. Biol. 2009, 4, 939–947. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Jiang, S. High throughput screening and characterization of HIV-1 entry inhibitors targeting gp41: Theories and techniques. Curr. Pharm. Des. 2004, 10, 1827–1843. [Google Scholar] [CrossRef] [PubMed]
- Shui, X.; Lu, X.; Gao, Y.; Liu, C.; Ren, F.; Jiang, Q.; Zhang, H.; Zhao, B.; Zheng, Z. A mammalian two-hybrid system-based assay for small-molecular HIV fusion inhibitors targeting gp41. Antivir. Res. 2011, 90, 54–63. [Google Scholar] [CrossRef]
- Lin, P.F.; Blair, W.; Wang, T.; Spicer, T.; Guo, Q.; Zhou, N.; Gong, Y.F.; Wang, H.G.; Rose, R.; Yamanaka, G.; et al. A small molecule HIV-1 inhibitor that targets the HIV-1 envelope and inhibits CD4 receptor binding. Proc. Natl. Acad. Sci. USA 2003, 100, 11013–11018. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.-J.; Brik, A.; Wong, C.-H.; Kan, C.-C. Model System for High-Throughput Screening of Novel Human Immunodeficiency Virus Protease Inhibitors in Escherichia coli. Antimicrob. Agents Chemother. 2004, 48, 2437–2447. [Google Scholar] [CrossRef]
- Omoruyi, B.E.; Ighodaro, D.I.; Afolayan, A.J.; Bradley, G. Inhibition of HIV-1 Protease by Carpobrotus edulis (L.). Evid. Based Complement Altern. Med. 2020, 2020, 9648056. [Google Scholar] [CrossRef]
- Painter, M.M.; Zimmerman, G.E.; Merlino, M.S.; Robertson, A.W.; Terry, V.H.; Ren, X.; McLeod, M.R.; Gomez-Rodriguez, L.; Garcia, K.A.; Leonard, J.A.; et al. Concanamycin A counteracts HIV-1 Nef to enhance immune clearance of infected primary cells by cytotoxic T lymphocytes. Proc. Natl. Acad. Sci. USA 2020, 117, 23835–23846. [Google Scholar] [CrossRef]
- Hyde, K.D.; Xu, J.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T.; Abeywickrama, P.D.; Aluthmuhandiram, J.V.S.; Brahamanage, R.S.; Brooks, S.; et al. The amazing potential of fungi: 50 ways we can exploit fungi industrially. Fungal Divers. 2019, 97, 1–136. [Google Scholar] [CrossRef]
- Nair, D.N.; Padmavathy, S. Impact of endophytic microorganisms on plants, environment and humans. Sci. World J. 2014, 2014, 250693. [Google Scholar] [CrossRef]
- Gakuubi, M.M.; Munusamy, M.; Liang, Z.X.; Ng, S.B. Fungal Endophytes: A Promising Frontier for Discovery of Novel Bioactive Compounds. J. Fungi 2021, 7, 786. [Google Scholar] [CrossRef]
- Stierle, A.A.; Stierle, D.B. Bioactive Secondary Metabolites Produced by the Fungal Endophytes of Conifers. Nat. Prod. Commun. 2015, 10, 1671–1682. [Google Scholar] [CrossRef]
- Wellensiek, B.P.; Ramakrishnan, R.; Bashyal, B.P.; Eason, Y.; Gunatilaka, A.A.; Ahmad, N. Inhibition of HIV-1 Replication by Secondary Metabolites from Endophytic Fungi of Desert Plants. Open Virol. J. 2013, 7, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Raut, J.K. Mushroom: A potent source of natural antiviral drugs. Appl. Sci. Technol. Ann. 2020, 1, 81–91. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Rossman, A.Y. Where are all the undescribed fungi? Phytopathology 1997, 87, 888–891. [Google Scholar] [CrossRef]
- Schmit, J.; Mueller, G. An estimate of the lower limit of global fungal diversity. Biodivers. Conserv. 2007, 16, 99–111. [Google Scholar] [CrossRef]
- Yang, S.W.; Mierzwa, R.; Terracciano, J.; Patel, M.; Gullo, V.; Wagner, N.; Baroudy, B.; Puar, M.; Chan, T.M.; Chu, M. Sch 213766, a novel chemokine receptor CCR-5 inhibitor from Chaetomium globosum. J. Antibiot. 2007, 60, 524–528. [Google Scholar] [CrossRef]
- Henrich, T.J.; Kuritzkes, D.R. HIV-1 entry inhibitors: Recent development and clinical use. Curr. Opin. Virol. 2013, 3, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Omura, S.; Tanaka, H.; Matsuzaki, K.; Ikeda, H.; Masuma, R. Isochromophilones I and II, novel inhibitors against gp120-CD4 binding from Penicillium sp. J. Antibiot. 1993, 46, 1908–1911. [Google Scholar] [CrossRef]
- Luo, X.; Lin, X.; Tao, H.; Wang, J.; Li, J.; Yang, B.; Zhou, X.; Liu, Y. Isochromophilones A–F, Cytotoxic Chloroazaphilones from the Marine Mangrove Endophytic Fungus Diaporthe sp. SCSIO 41011. J. Nat. Prod. 2018, 81, 934–941. [Google Scholar] [CrossRef]
- Yoganathan, K.; Rossant, C.; Glover, R.P.; Cao, S.; Vittal, J.J.; Ng, S.; Huang, Y.; Buss, A.D.; Butler, M.S. Inhibition of the Human Chemokine Receptor CCR5 by Variecolin and Variecolol and Isolation of Four New Variecolin Analogues, Emericolins A−D, from Emericella aurantiobrunnea. J. Nat. Prod. 2004, 67, 1681–1684. [Google Scholar] [CrossRef] [PubMed]
- Min, B.S.; Miyashiro, H.; Hattori, M. Inhibitory effects of quinones on RNase H activity associated with HIV-1 reverse transcriptase. Phytother. Res. 2002, 16, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Anstett, K.; Brenner, B.; Mesplède, T.; Wainberg, M.A. HIV-1 Resistance to Dolutegravir Is Affected by Cellular Histone Acetyltransferase Activity. J. Virol. 2017, 91, e00912-17. [Google Scholar] [CrossRef] [PubMed]
- Bashyal, B.P.; Wellensiek, B.P.; Ramakrishnan, R.; Faeth, S.H.; Ahmad, N.; Gunatilaka, A.A. Altertoxins with potent anti-HIV activity from Alternaria tenuissima QUE1Se, a fungal endophyte of Quercus emoryi. Bioorg. Med. Chem. 2014, 22, 6112–6116. [Google Scholar] [CrossRef]
- Melappa, G.; Hemmanur, K.; Nithin, S.; Poojari, C.; Kakunje, G.; Ryavalad, C. In Vitro Anti-HIV Activity of Partially Purified Coumarin(s) Isolated from Fungal Endophyte, Alternaria Species of Calophyllum inophyllum. Pharmacol. Pharm. 2015, 6, 321–328. [Google Scholar] [CrossRef]
- Richetta, C.; Tu, N.Q.; Delelis, O. Different Pathways Conferring Integrase Strand-Transfer Inhibitors Resistance. Viruses 2022, 14, 2591. [Google Scholar] [CrossRef]
- Singh, S.B.; Zink, D.L.; Guan, Z.; Collado, J.; Pelaez, F.; Felock, P.J.; Hazuda, D.J. Isolation, structure, and HIV-1 integrase inhibitory activity of Xanthoviridicatin E and F, two novel fungal metabolites produced by Penicillium chrysogenum. Helv. Chim. Acta 2003, 86, 3380–3385. [Google Scholar] [CrossRef]
- Jayasuriya, H.; Guan, Z.; Polishook, J.D.; Dombrowski, A.W.; Felock, P.J.; Hazuda, D.J.; Singh, S.B. Isolation, structure, and HIV-1 integrase inhibitory activity of Cytosporic acid, a fungal metabolite produced by a Cytospora sp. J. Nat. Prod. 2003, 66, 551–553. [Google Scholar] [CrossRef] [PubMed]
- Mandala, S.M.; Thornton, R.A.; Frommer, B.R.; Curotto, J.E.; Rozdilsky, W.; Kurtz, M.B.; Giacobbe, R.A.; Bills, G.F.; Cabello, M.A.; Martín, I.; et al. The discovery of australifungin, a novel inhibitor of sphinganine N-acyltransferase from Sporormiella australis. Producing organism, fermentation, isolation, and biological activity. J. Antibiot. 1995, 48, 349–356. [Google Scholar] [CrossRef]
- Guo, Z. The modification of natural products for medical use. Acta Pharm Sin B. 2017, 7, 119–136. [Google Scholar] [CrossRef]
- Shiomi, K.; Matsui, R.; Isozaki, M.; Chiba, H.; Sugai, T.; Yamaguchi, Y.; Masuma, R.; Tomoda, H.; Chiba, T.; Yan, H.; et al. Fungal phenalenones inhibit HIV-1 integrase. J. Antibiot. 2005, 58, 65–68. [Google Scholar] [CrossRef]
- Venugopala, K.N.; Rashmi, V.; Odhav, B. Review on Natural Coumarin Lead Compounds for Their Pharmacological Activity. BioMed. Res. Int. 2013, 2013, 963248. [Google Scholar] [CrossRef] [PubMed]
- Ryang, J.; Yan, Y.; Song, Y.; Liu, F.; Ng, T.B. Anti-HIV, antitumor and immunomodulatory activities of paclitaxel from fermentation broth using molecular imprinting technique. AMB Express 2019, 9, 194. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.B.; Ondeyka, J.G.; Tsipouras, N.; Ruby, C.; Sardana, V.; Schulman, M.; Sanchez, M.; Pelaez, F.; Stahlhut, M.W.; Munshi, S.; et al. Hinnuliquinone, a C2-symmetric dimeric non-peptide fungal metabolite inhibitor of HIV-1 protease. Biochem. Biophys. Res. Commun. 2004, 324, 108–113. [Google Scholar] [CrossRef]
- Sato, N.; Ma, C.M.; Komatsu, K.; Hattori, M. Triterpene-farnesyl hydroquinone conjugates from Ganoderma sinense. J. Nat. Prod. 2009, 72, 958–961. [Google Scholar] [CrossRef] [PubMed]
- Sebastian, N.T.; Collins, K.L. Targeting HIV latency: Resting memory T cells, hematopoietic progenitor cells and future directions. Expert Rev. Anti-Infect. Ther. 2014, 12, 1187–1201. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fromentin, R.; Chomont, N. HIV persistence in subsets of CD4+ T cells: 50 shades of reservoirs. Semin. Immunol. 2021, 51, 101438. [Google Scholar] [CrossRef]
- Haigentz, M.; Moore, P.; Bimali, M.; Cooley, T.; Sparano, J.; Rudek, M.; Ratner, L.; Henry, D.; Ramos, J.; Deeken, J.; et al. Safety and Tolerability of Carboplatin and Paclitaxel in Cancer Patients with HIV (AMC-078), an AIDS Malignancy Consortium (AMC) Study. Oncologist 2022, 27, e623–e624. [Google Scholar] [CrossRef] [PubMed]
- Abbas, U.L.; Glaubius, R.; Mubayi, A.; Hood, G.; Mellors, J.W. Antiretroviral therapy and pre-exposure prophylaxis: Combined impact on HIV transmission and drug resistance in South Africa. J. Infect. Dis. 2013, 208, 224–234. [Google Scholar] [CrossRef]
- Churchill, M.J.; Deeks, S.G.; Margolis, D.M.; Siliciano, R.F.; Swanstrom, R. HIV reservoirs: What, where and how to target them. Nat. Rev. Microbiol. 2016, 14, 55–60. [Google Scholar] [CrossRef]
- Niu, S.; Liu, D.; Shao, Z.; Proksch, P.; Lin, W. Eutypellazines N−S, new thiodiketopiperazines from a deep sea sediment derived fungus Eutypella sp. with anti-VRE activities. Tetrahedron Lett. 2017, 58, 3695–3699. [Google Scholar] [CrossRef]
- Stoszko, M.; Al-Hatmi, A.M.S.; Skriba, A.; Roling, M.; Ne, E.; Crespo, R.; Mueller, Y.M.; Najafzadeh, M.J.; Kang, J.; Ptackova, R.; et al. Gliotoxin, identified from a screen of fungal metabolites, disrupts 7SK snRNP, releases P-TEFb, and reverses HIV-1 latency. Sci. Adv. 2020, 6, eaba6617. [Google Scholar] [CrossRef]
- Xing, S.; Siliciano, R.F. Targeting HIV latency: Pharmacologic strategies toward eradication. Drug Discov. Today 2013, 18, 541–551. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; del-Val, E.; Larsen, J. Ecological functions of Trichoderma spp. and their secondary metabolites in the rhizosphere: Interactions with plants. FEMS Microbiol. Ecol. 2012, 18, 541–551. [Google Scholar] [CrossRef]
- Spivak, A.M.; Planelles, V. Novel Latency Reversal Agents for HIV-1 Cure. Annu. Rev. Med. 2018, 69, 421–436. [Google Scholar] [CrossRef] [PubMed]
- Richter, S.N. Strategies for inhibiting function of HIV-1 accessory proteins: A necessary route to AIDS therapy. Curr. Med. Chem. 2009, 16, 267–286. [Google Scholar] [CrossRef]
- Sugui, J.A.; Pardo, J.; Chang, Y.C.; Zarember, K.A.; Nardone, G.; Galvez, E.M.; Müllbacher, A.; Gallin, J.I.; Simon, M.M.; Kwon-Chung, K.J. Gliotoxin is a virulence factor of Aspergillus fumigatus: GliP deletion attenuates virulence in mice immunosuppressed with hydrocortisone. Eukaryot. Cell 2007, 6, 1562–1569. [Google Scholar] [CrossRef]
- Scharf, D.H.; Heinekamp, T.; Remme, N.; Hortschansky, P.; Brakhage, A.A.; Hertweck, C. Biosynthesis and function of gliotoxin in Aspergillus fumigatus. Appl. Microbiol. Biotechnol. 2012, 93, 467–472. [Google Scholar] [CrossRef]
- Watanabe, N.; Nishihara, Y.; Yamaguchi, T.; Koito, A.; Miyoshi, H.; Kakeya, H.; Osada, H. Fumagillin suppresses HIV-1 infection of macrophages through the inhibition of Vpr activity. FEBS Lett. 2006, 580, 2598–2602. [Google Scholar] [CrossRef] [PubMed]
- Kamata, M.; Wu, R.P.; An, D.S.; Saxe, J.P.; Damoiseaux, R.; Phelps, M.E.; Huang, J.; Chen, I.S. Cell-based chemical genetic screen identifies damnacanthal as an inhibitor of HIV-1 Vpr induced cell death. Biochem. Biophys. Res. Commun. 2006, 348, 1101–1106. [Google Scholar] [CrossRef] [PubMed]
- van den Heever, J.P.; Thompson, T.S.; Curtis, J.M.; Ibrahim, A.; Pernal, S.F. Fumagillin: An overview of recent scientific advances and their significance for apiculture. J. Agric. Food Chem. 2014, 62, 2728–2737. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Orhan, I.E.; Banach, M.; Rollinger, J.M.; Barreca, D.; Weckwerth, W.; Bauer, R.; Bayer, E.A.; et al. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Kaul, S.; Gupta, S.; Ahmed, M.; Dhar, M.K. Endophytic fungi from medicinal plants: A treasure hunt for bioactive metabolites. Phytochem. Rev. 2012, 11, 487–505. [Google Scholar] [CrossRef]
- Truax, N.J.; Romo, D. Bridging the gap between natural product synthesis and drug discovery. Nat. Prod. Rep. 2020, 37, 1436–1453. [Google Scholar] [CrossRef]
- Frediansyah, A.; Sofyantoro, F.; Alhumaid, S.; Al Mutair, A.; Albayat, H.; Altaweil, H.I.; Al-Afghani, H.M.; AlRamadhan, A.A.; AlGhazal, M.R.; Turkistani, S.A.; et al. Microbial Natural Products with Antiviral Activities, Including Anti-SARS-CoV-2: A Review. Molecules 2022, 27, 4305. [Google Scholar] [CrossRef]
- Sorokina, M.; Merseburger, P.; Rajan, K.; Yirik, M.A.; Steinbeck, C. COCONUT online: Collection of Open Natural Products database. J. Cheminf. 2021, 13, 2. [Google Scholar] [CrossRef]
- van Santen, J.; Poynton, E.; Iskakova, D.; McMann, E.; Alsup, T.; Clark, T.; Fergusson, C.; Fewer, D.; Hughes, A.; McCadden, C.; et al. The Natural Products Atlas 2.0: A Database of microbially-derived natural products. Nucleic Acids Res. 2022, 50, D1317–D1323. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Yang, B.; Liu, J.; Xun, T.; Liu, Y.; Zhou, X. Penicillixanthone A, a marine-derived dual-coreceptor antagonist as anti-HIV-1 agent. Nat. Prod. Res. 2019, 33, 1467–1471. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Li, L.; Zhu, T.; Ba, M.; Li, G.; Gu, Q.; Guo, Y.; Li, D. Phenylspirodrimanes with Anti-HIV Activity from the Sponge-Derived Fungus Stachybotrys chartarum MXH-X73. J. Nat. Prod. 2013, 76, 2298–2306. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.-W.; Sun, B.-D.; Chen, X.-L.; Liu, X.-Z.; Che, Y.-S. Helotialins A—C, Anti-HIV Metabolites from a Helotialean Ascomycete. Chin. J. Nat. Med. 2009, 7, 140–144. [Google Scholar] [CrossRef]
- Hamano, K.; Kinoshita-Okami, M.; Minagawa, K.; Haruyama, H.; Kinoshita, T.; Hosoya, T.; Furuya, K.; Kinoshita, K.; Tabata, K.; Hemmi, A.; et al. Aquastatin A, an inhibitor of mammalian adenosine triphosphatases from Fusarium aquaeductuum. Taxonomy, fermentation, isolation, structure determination and biological properties. J. Antibiot. 1993, 46, 1648–1657. [Google Scholar] [CrossRef][Green Version]
- Singh, S.B.; Zink, D.L.; Bills, G.F.; Pelaez, F.; Teran, A.; Collado, J.; Silverman, K.C.; Lingham, R.B.; Felock, P.; Hazuda, D.J. Discovery, structure and HIV-1 integrase-inhibitory activities of integracins, novel dimeric alkyl aromatics from Cytonaema. Tetrahedron Lett. 2002, 43, 1617–1620. [Google Scholar] [CrossRef]
- Srisapoomi, T.; Ichiyanagi, T.; Nakajima, H.; Aimi, T.; Boonlue, S. Biological Activities of Integric Acid Isolated from the Wood-Decay Fungus Xylaria feejeensis 2FB-PPM08M. Chiang Mai J. Sci. 2015, 42, 71–79. [Google Scholar]

{kind=link}
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nzimande, B.; Makhwitine, J.P.; Mkhwanazi, N.P.; Ndlovu, S.I. Developments in Exploring Fungal Secondary Metabolites as Antiviral Compounds and Advances in HIV-1 Inhibitor Screening Assays. Viruses 2023, 15, 1039. https://doi.org/10.3390/v15051039
Nzimande B, Makhwitine JP, Mkhwanazi NP, Ndlovu SI. Developments in Exploring Fungal Secondary Metabolites as Antiviral Compounds and Advances in HIV-1 Inhibitor Screening Assays. Viruses. 2023; 15(5):1039. https://doi.org/10.3390/v15051039
Chicago/Turabian StyleNzimande, Bruce, John P. Makhwitine, Nompumelelo P. Mkhwanazi, and Sizwe I. Ndlovu. 2023. "Developments in Exploring Fungal Secondary Metabolites as Antiviral Compounds and Advances in HIV-1 Inhibitor Screening Assays" Viruses 15, no. 5: 1039. https://doi.org/10.3390/v15051039
APA StyleNzimande, B., Makhwitine, J. P., Mkhwanazi, N. P., & Ndlovu, S. I. (2023). Developments in Exploring Fungal Secondary Metabolites as Antiviral Compounds and Advances in HIV-1 Inhibitor Screening Assays. Viruses, 15(5), 1039. https://doi.org/10.3390/v15051039