
Epidemiological Surveillance Reveals the Rise and Establishment of the Omicron SARS-CoV-2 Variant in Brazil
 , , , , ,
, , , , ,  , , , and
, , , and 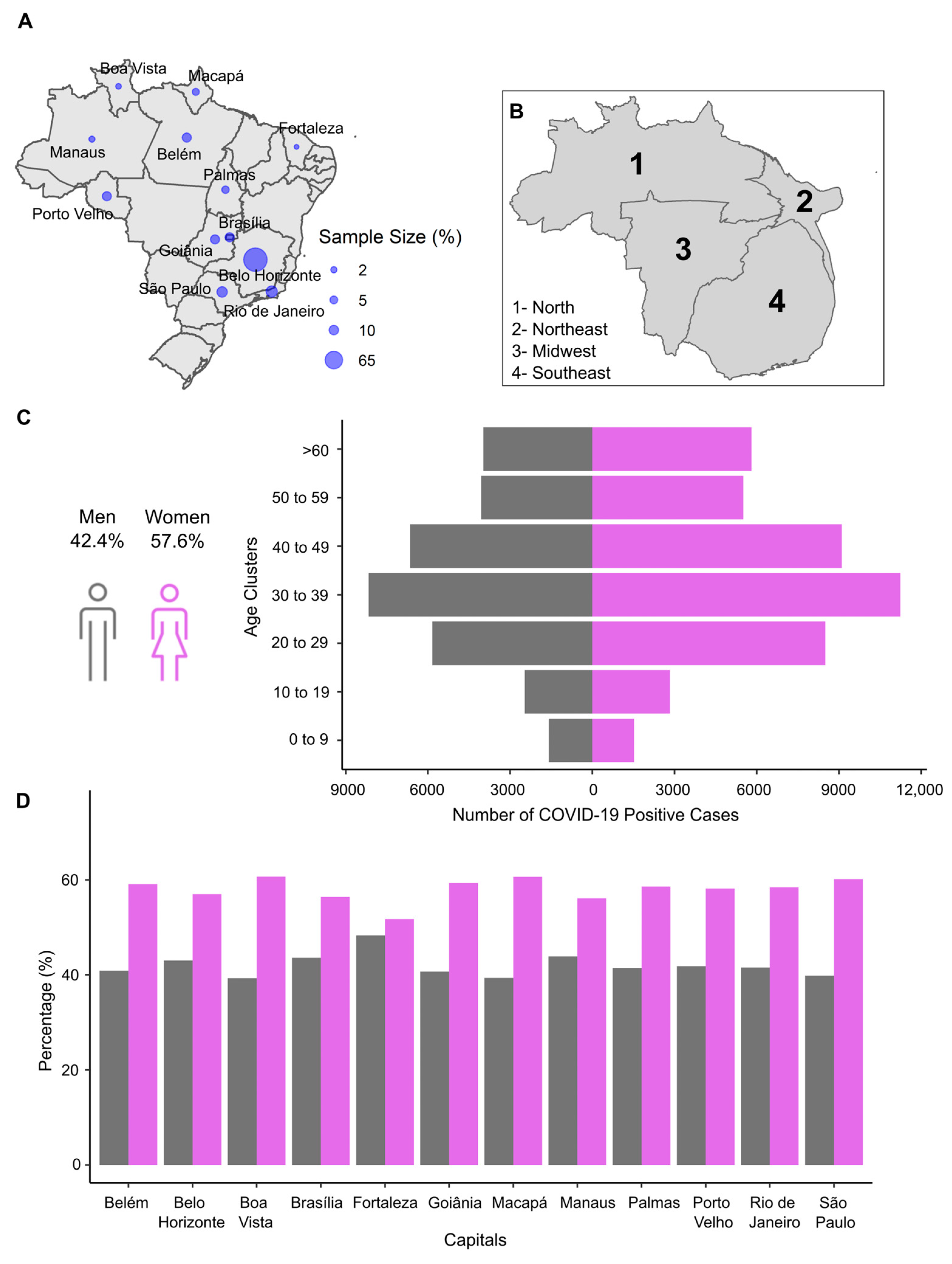{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Genomic Surveillance by RT-qPCR Genotyping and Whole-Genome Sequencing
2.2.1. Descriptive Sample Profile and SARS-CoV-2 Variants Genotyping by RT-qPCR
2.2.2. SARS-CoV-2 Genome Sequencing
2.2.3. Viral Genome Assembly
2.2.4. Lineage Classification and Phylogenetic Analysis
2.3. Ct Value and Viral Load
2.4. Symptomatology Meta-Analysis
3. Results
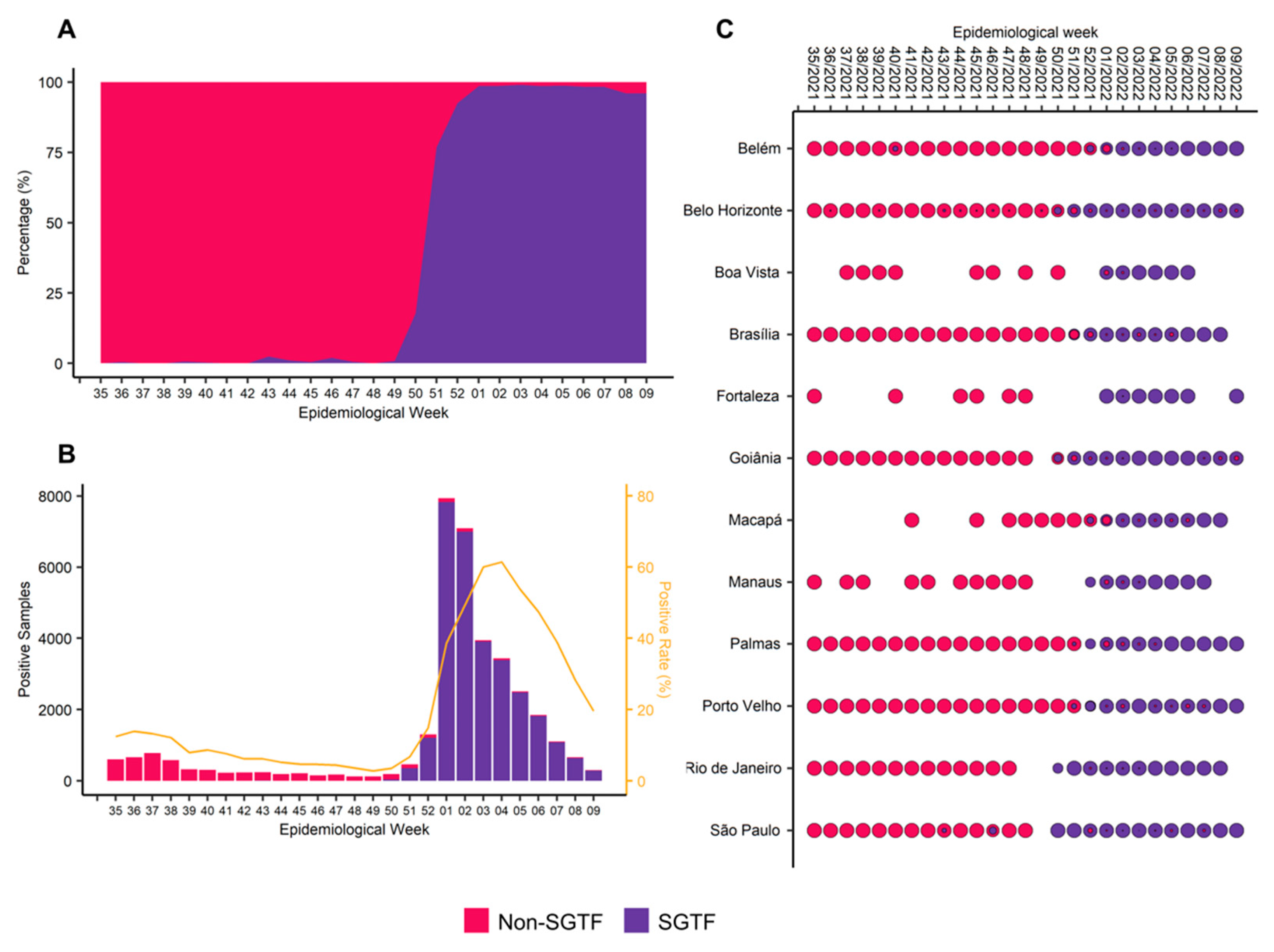
3.1. The Rapid Replacement and Establishment of Omicron VOC in Brazilian Territory
3.2. Sequencing Metrics, Classification, and Phylogeny
3.3. Omicron-Infected Individuals Present a Lower Viral Load in the Upper Respiratory Tract
3.4. Effect of SARS-CoV-2 Variants on Symptomatology and Clinical Data
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 23 February 2023).
- Brasil Painel Coronavírus. Available online: https://covid.saude.gov.br (accessed on 11 December 2022).
- Thanh Le, T.; Andreadakis, Z.; Kumar, A.; Gómez Román, R.; Tollefsen, S.; Saville, M.; Mayhew, S. The COVID-19 Vaccine Development Landscape. Nat. Rev. Drug Discov. 2020, 19, 305–306. [Google Scholar] [CrossRef]
- Sobczak, M.; Pawliczak, R. Factors That Affect the COVID-19 Pandemic in Summer 2022 Compared to Summer 2021. Int. J. Environ. Res. Public Health 2022, 19, 12561. [Google Scholar] [CrossRef]
- Basso, P.; Negro, C.; Cegolon, L.; Larese Filon, F. Risk of Vaccine Breakthrough SARS-CoV-2 Infection and Associated Factors in Healthcare Workers of Trieste Teaching Hospitals (North-Eastern Italy). Viruses 2022, 14, 336. [Google Scholar] [CrossRef]
- Cegolon, L.; Ronchese, F.; Ricci, F.; Negro, C.; Larese-Filon, F. SARS-CoV-2 Infection in Health Care Workers of Trieste (North-Eastern Italy), 1 October 2020-7 February 2022: Occupational Risk and the Impact of the Omicron Variant. Viruses 2022, 14, 1663. [Google Scholar] [CrossRef] [PubMed]
- Cegolon, L.; Negro, C.; Mastrangelo, G.; Filon, F.L. ORCHESTRA working group Primary SARS-CoV-2 Infections, Re-Infections and Vaccine Effectiveness during the Omicron Transmission Period in Healthcare Workers of Trieste and Gorizia (Northeast Italy), 1 December 2021–31 May 2022. Viruses 2022, 14, 2688. [Google Scholar] [CrossRef]
- World Health Organization. Tracking SARS-CoV-2 Variants. Available online: https://www.who.int/activities/tracking-SARS-CoV-2-variants (accessed on 13 December 2022).
- Alcantara, L.C.J.; Nogueira, E.; Shuab, G.; Tosta, S.; Fristch, H.; Pimentel, V.; Souza-Neto, J.A.; Coutinho, L.L.; Fukumasu, H.; Sampaio, S.C.; et al. SARS-CoV-2 Epidemic in Brazil: How the Displacement of Variants Has Driven Distinct Epidemic Waves. Virus Res. 2022, 315, 198785. [Google Scholar] [CrossRef]
- Thakur, V.; Bhola, S.; Thakur, P.; Patel, S.K.S.; Kulshrestha, S.; Ratho, R.K.; Kumar, P. Waves and Variants of SARS-CoV-2: Understanding the Causes and Effect of the COVID-19 Catastrophe. Infection 2022, 50, 309–325. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Gao, L.; Zhou, Q.; Yu, K.; Sun, F. Effectiveness of COVID-19 Vaccines against SARS-CoV-2 Variants of Concern: A Systematic Review and Meta-Analysis. BMC Med. 2022, 20, 200. [Google Scholar] [CrossRef] [PubMed]
- Volz, E.; Mishra, S.; Chand, M.; Barrett, J.C.; Johnson, R.; Geidelberg, L.; Hinsley, W.R.; Laydon, D.J.; Dabrera, G.; O’Toole, Á.; et al. Assessing Transmissibility of SARS-CoV-2 Lineage B.1.1.7 in England. Nature 2021, 593, 266–269. [Google Scholar] [CrossRef]
- Tegally, H.; Wilkinson, E.; Giovanetti, M.; Iranzadeh, A.; Fonseca, V.; Giandhari, J.; Doolabh, D.; Pillay, S.; San, E.J.; Msomi, N.; et al. Detection of a SARS-CoV-2 Variant of Concern in South Africa. Nature 2021, 592, 438–443. [Google Scholar] [CrossRef]
- Faria, N.R.; Mellan, T.A.; Whittaker, C.; Claro, I.M.; Candido, D.d.S.; Mishra, S.; Crispim, M.A.E.; Sales, F.C.S.; Hawryluk, I.; McCrone, J.T.; et al. Genomics and Epidemiology of the P.1 SARS-CoV-2 Lineage in Manaus, Brazil. Science 2021, 372, 815–821. [Google Scholar] [CrossRef]
- Naveca, F.G.; Nascimento, V.; de Souza, V.C.; Corado, A.d.L.; Nascimento, F.; Silva, G.; Costa, Á.; Duarte, D.; Pessoa, K.; Mejía, M.; et al. COVID-19 in Amazonas, Brazil, Was Driven by the Persistence of Endemic Lineages and P.1 Emergence. Nat. Med. 2021, 27, 1230–1238. [Google Scholar] [CrossRef]
- Cherian, S.; Potdar, V.; Jadhav, S.; Yadav, P.; Gupta, N.; Das, M.; Rakshit, P.; Singh, S.; Abraham, P.; Panda, S.; et al. SARS-CoV-2 Spike Mutations, L452R, T478K, E484Q and P681R, in the Second Wave of COVID-19 in Maharashtra, India. Microorganisms 2021, 9, 1542. [Google Scholar] [CrossRef] [PubMed]
- Viana, R.; Moyo, S.; Amoako, D.G.; Tegally, H.; Scheepers, C.; Althaus, C.L.; Anyaneji, U.J.; Bester, P.A.; Boni, M.F.; Chand, M.; et al. Rapid Epidemic Expansion of the SARS-CoV-2 Omicron Variant in Southern Africa. Nature 2022, 603, 679–686. [Google Scholar] [CrossRef]
- Tao, K.; Tzou, P.L.; Nouhin, J.; Gupta, R.K.; de Oliveira, T.; Kosakovsky Pond, S.L.; Fera, D.; Shafer, R.W. The Biological and Clinical Significance of Emerging SARS-CoV-2 Variants. Nat. Rev. Genet. 2021, 22, 757–773. [Google Scholar] [CrossRef]
- Nonaka, C.K.V.; Gräf, T.; Barcia, C.A.d.L.; Costa, V.F.; de Oliveira, J.L.; Passos, R.d.H.; Bastos, I.N.; de Santana, M.C.B.; Santos, I.M.; de Sousa, K.A.F.; et al. SARS-CoV-2 Variant of Concern P.1 (Gamma) Infection in Young and Middle-Aged Patients Admitted to the Intensive Care Units of a Single Hospital in Salvador, Northeast Brazil, February 2021. Int. J. Infect. Dis. 2021, 111, 47–54. [Google Scholar] [CrossRef]
- Taylor, C.A.; Patel, K.; Pham, H.; Whitaker, M.; Anglin, O.; Kambhampati, A.K.; Milucky, J.; Chai, S.J.; Kirley, P.D.; Alden, N.B.; et al. Severity of Disease Among Adults Hospitalized with Laboratory-Confirmed COVID-19 Before and During the Period of SARS-CoV-2 B.1.617.2 (Delta) Predominance—COVID-NET, 14 States, January-August 2021. MMWR. Morb. Mortal. Wkly. Rep. 2021, 70, 1513–1519. [Google Scholar] [CrossRef] [PubMed]
- Kupferschmidt, K.; Vogel, G. How Bad Is Omicron? Some Clues Are Emerging. Science 2021, 374, 1304–1305. [Google Scholar] [CrossRef] [PubMed]
- Kannan, S.; Shaik Syed Ali, P.; Sheeza, A. Omicron (B.1.1.529)—Variant of Concern—Molecular Profile and Epidemiology: A Mini Review. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 8019–8022. [Google Scholar] [CrossRef]
- World Health Organization Classification of Omicron (B.1.1.529): SARS-CoV-2 Variant of Concern. Available online: https://www.who.int/news/item/26-11-2021-classification-of-omicron-(b.1.1.529)-sars-cov-2-variant-of-concern (accessed on 23 February 2023).
- Li, B.; Lu, X.; McAndrews, K.M.; Kalluri, R. Mutations in the Spike RBD of SARS-CoV-2 Omicron Variant May Increase Infectivity without Dramatically Altering the Efficacy of Current Multi-Dosage Vaccinations. bioRxiv 2021. [Google Scholar] [CrossRef]
- McMillen, T.; Jani, K.; Robilotti, E.V.; Kamboj, M.; Babady, N.E. The Spike Gene Target Failure (SGTF) Genomic Signature Is Highly Accurate for the Identification of Alpha and Omicron SARS-CoV-2 Variants. Sci. Rep. 2022, 12, 18968. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhi, H.; Teng, Y. The Outbreak of SARS-CoV-2 Omicron Lineages, Immune Escape, and Vaccine Effectivity. J. Med. Virol. 2022, 95, e28138. [Google Scholar] [CrossRef] [PubMed]
- Focosi, D.; McConnell, S.; Casadevall, A. The Omicron Variant of Concern: Diversification and Convergent Evolution in Spike Protein, and Escape from Anti-Spike Monoclonal Antibodies. Drug Resist. Updat. 2022, 65, 100882. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.P.; de Lima, A.B.; Alvim, L.B.; Malta, F.S.V.; Mendonça, C.P.T.B.; Fonseca, P.L.C.; Moreira, F.R.R.; Queiroz, D.C.; Ferreira, J.G.G.; Ferreira, A.C.S.; et al. Delta Variant of SARS-CoV-2 Replacement in Brazil: A National Epidemiologic Surveillance Program. Viruses 2022, 14, 847. [Google Scholar] [CrossRef] [PubMed]
- Francis, A.I.; Ghany, S.; Gilkes, T.; Umakanthan, S. Review of COVID-19 Vaccine Subtypes, Efficacy and Geographical Distributions. Postgrad. Med. J. 2022, 98, 389–394. [Google Scholar] [CrossRef]
- Menezes, D.; Fonseca, P.L.C.; de Araújo, J.L.F.; de Souza, R.P. SARS-CoV-2 Genomic Surveillance in Brazil: A Systematic Review with Scientometric Analysis. Viruses 2022, 14, 2715. [Google Scholar] [CrossRef]
- Koelle, K.; Martin, M.A.; Antia, R.; Lopman, B.; Dean, N.E. The Changing Epidemiology of SARS-CoV-2. Science 2022, 375, 1116–1121. [Google Scholar] [CrossRef]
- Noureddine, F.Y.; Chakkour, M.; El Roz, A.; Reda, J.; Al Sahily, R.; Assi, A.; Joma, M.; Salami, H.; Hashem, S.J.; Harb, B.; et al. The Emergence of SARS-CoV-2 Variant(s) and Its Impact on the Prevalence of COVID-19 Cases in the Nabatieh Region, Lebanon. Med. Sci. 2021, 9, 40. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2—Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009; ISBN 978-0-387-98140-6. [Google Scholar]
- Moreira, F. ViralUnity. Available online: https://github.com/filiperomero2/ViralUnity (accessed on 25 December 2022).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Li, H. Minimap2: Pairwise Alignment for Nucleotide Sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Li, H. A Statistical Framework for SNP Calling, Mutation Discovery, Association Mapping and Population Genetical Parameter Estimation from Sequencing Data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef]
- Quinlan, A.R.; Hall, I.M. BEDTools: A Flexible Suite of Utilities for Comparing Genomic Features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef]
- O’Toole, Á.; Scher, E.; Underwood, A.; Jackson, B.; Hill, V.; McCrone, J.T.; Colquhoun, R.; Ruis, C.; Abu-Dahab, K.; Taylor, B.; et al. Assignment of Epidemiological Lineages in an Emerging Pandemic Using the Pangolin Tool. Virus Evol. 2021, 7, veab064. [Google Scholar] [CrossRef]
- Aksamentov, I.; Roemer, C.; Hodcroft, E.; Neher, R. Nextclade: Clade Assignment, Mutation Calling and Quality Control for Viral Genomes. J. Open Source Softw. 2021, 6, 3773. [Google Scholar] [CrossRef]
- Hadfield, J.; Megill, C.; Bell, S.M.; Huddleston, J.; Potter, B.; Callender, C.; Sagulenko, P.; Bedford, T.; Neher, R.A. NextStrain: Real-Time Tracking of Pathogen Evolution. Bioinformatics 2018, 34, 4121–4123. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R.; Teeling, E. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Yang, Z. Maximum Likelihood Phylogenetic Estimation from DNA Sequences with Variable Rates over Sites: Approximate Methods. J. Mol. Evol. 1994, 39, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Moreira, F.R.R.; D’Arc, M.; Mariani, D.; Herlinger, A.L.; Schiffler, F.B.; Rossi, Á.D.; Leitão, I.D.C.; Miranda, T.D.S.; Cosentino, M.A.C.; Tôrres, M.C.D.P.; et al. Epidemiological Dynamics of SARS-CoV-2 VOC Gamma in Rio de Janeiro, Brazil. Virus Evol. 2021, 7, veab087. [Google Scholar] [CrossRef] [PubMed]
- Hay, J.A.; Kennedy-Shaffer, L.; Kanjilal, S.; Lennon, N.J.; Gabriel, S.B.; Lipsitch, M.; Mina, M.J. Estimating Epidemiologic Dynamics from Cross-Sectional Viral Load Distributions. Science 2021, 373, eabh0635. [Google Scholar] [CrossRef]
- Ministério da Saúde Banco de Dados de Síndrome Respiratória Aguda Grave. Available online: https://opendatasus.saude.gov.br/dataset/srag-2021-e-2022 (accessed on 29 September 2022).
- Center for Disease Control and Prevention. About COVID-19 Epidemiology. Available online: https://www.cdc.gov/coronavirus/2019-ncov/science/about-epidemiology/index.html (accessed on 26 December 2022).
- World Health Organization COVID-19: Variants. Available online: https://www.who.int/westernpacific/emergencies/covid-19/information/covid-19-variants (accessed on 28 December 2022).
- Governo do Estado de São Paulo. SP Confirma Os Dois Primeiros Casos Da Variante Ômicron No Brasil|Governo Do Estado de São Paulo. Available online: https://www.saopaulo.sp.gov.br/noticias-coronavirus/sp-confirma-os-dois-primeiros-casos-da-variante-omicron-no-brasil/ (accessed on 28 December 2022).
- Viana, P. Aeroportos Mais Movimentados Do Brasil. Available online: https://www.aeroflap.com.br/veja-quais-foram-os-aeroportos-mais-movimentados-do-brasil-no-inicio-de-2022/ (accessed on 23 February 2023).
- Chakkour, M.; Salami, A.; Olleik, D.; Kamal, I.; Noureddine, F.Y.; Roz, A.E.; Ghssein, G. Risk Markers of COVID-19, a Study from South-Lebanon. COVID 2022, 2, 867–876. [Google Scholar] [CrossRef]
- Lewis, D. Superspreading Drives the COVID Pandemic—And Could Help to Tame It. Nature 2021, 590, 544–546. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.Z.; Koopmans, M.; Fisman, D.N.; Gu, F.X. Understanding Why Superspreading Drives the COVID-19 Pandemic but Not the H1N1 Pandemic. Lancet Infect. Dis. 2021, 21, 1203–1204. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhao, S.; Guo, Z.; Yuan, Z.; Ran, J.; Wu, L.; Yu, L.; Li, H.; Shi, Y.; He, D. Differences in the Superspreading Potentials of COVID-19 across Contact Settings. BMC Infect. Dis. 2022, 22, 936. [Google Scholar] [CrossRef]
- Aragão, D.P.; Dos Santos, D.H.; Mondini, A.; Gonçalves, L.M.G. National Holidays and Social Mobility Behaviors: Alternatives for Forecasting Covid-19 Deaths in Brazil. Int. J. Environ. Res. Public Health 2021, 18, 11595. [Google Scholar] [CrossRef]
- Kimura, I.; Yamasoba, D.; Tamura, T.; Nao, N.; Suzuki, T.; Oda, Y.; Mitoma, S.; Ito, J.; Nasser, H.; Zahradnik, J.; et al. Virological Characteristics of the SARS-CoV-2 Omicron BA.2 Subvariants, Including BA.4 and BA.5. Cell 2022, 185, 3992–4007.e16. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, M.; Priyanka; Choudhary, O.P. Emergence of Omicron Sub-Variant BA.2: Is It a Matter of Concern amid the COVID-19 Pandemic? Int. J. Surg. 2022, 99, 106581. [Google Scholar] [CrossRef]
- Valente, J. Brazil Confirms First Case of Omicron Subvariant. Available online: https://agenciabrasil.ebc.com.br/en/saude/noticia/2020-02/brazil-confirms-first-case-coronavirus (accessed on 28 December 2022).
- Teyssou, E.; Delagrèverie, H.; Visseaux, B.; Lambert-Niclot, S.; Brichler, S.; Ferre, V.; Marot, S.; Jary, A.; Todesco, E.; Schnuriger, A.; et al. The Delta SARS-CoV-2 Variant Has a Higher Viral Load than the Beta and the Historical Variants in Nasopharyngeal Samples from Newly Diagnosed COVID-19 Patients. J. Infect. 2021, 83, e1–e3. [Google Scholar] [CrossRef]
- Kidd, M.; Richter, A.; Best, A.; Cumley, N.; Mirza, J.; Percival, B.; Mayhew, M.; Megram, O.; Ashford, F.; White, T.; et al. S-Variant SARS-CoV-2 Lineage B1.1.7 Is Associated With Significantly Higher Viral Load in Samples Tested by TaqPath Polymerase Chain Reaction. J. Infect. Dis. 2021, 223, 1666–1670. [Google Scholar] [CrossRef]
- Yuasa, S.; Nakajima, J.; Takatsuki, Y.; Takahashi, Y.; Tani-Sassa, C.; Iwasaki, Y.; Nagano, K.; Sonobe, K.; Yoshimoto, T.; Nukui, Y.; et al. Viral Load of SARS-CoV-2 Omicron Is Not High despite Its High Infectivity. J. Med. Virol. 2022, 94, 5543–5546. [Google Scholar] [CrossRef]
- Sentis, C.; Billaud, G.; Bal, A.; Frobert, E.; Bouscambert, M.; Destras, G.; Josset, L.; Lina, B.; Morfin, F.; Gaymard, A.; et al. SARS-CoV-2 Omicron Variant, Lineage BA.1, Is Associated with Lower Viral Load in Nasopharyngeal Samples Compared to Delta Variant. Viruses 2022, 14, 919. [Google Scholar] [CrossRef]
- Puhach, O.; Adea, K.; Hulo, N.; Sattonnet, P.; Genecand, C.; Iten, A.; Jacquérioz, F.; Kaiser, L.; Vetter, P.; Eckerle, I.; et al. Infectious Viral Load in Unvaccinated and Vaccinated Individuals Infected with Ancestral, Delta or Omicron SARS-CoV-2. Nat. Med. 2022, 28, 1491–1500. [Google Scholar] [CrossRef]
- Laitman, A.M.; Lieberman, J.A.; Hoffman, N.G.; Roychoudhury, P.; Mathias, P.C.; Greninger, A.L. The SARS-CoV-2 Omicron Variant Does Not Have Higher Nasal Viral Loads Compared to the Delta Variant in Symptomatic and Asymptomatic Individuals. J. Clin. Microbiol. 2022, 60, e00139-22. [Google Scholar] [CrossRef] [PubMed]
- Iketani, S.; Liu, L.; Guo, Y.; Liu, L.; Chan, J.F.W.; Huang, Y.; Wang, M.; Luo, Y.; Yu, J.; Chu, H.; et al. Antibody Evasion Properties of SARS-CoV-2 Omicron Sublineages. Nature 2022, 604, 553–556. [Google Scholar] [CrossRef] [PubMed]
- Hong, Q.; Han, W.; Li, J.; Xu, S.; Wang, Y.; Xu, C.; Li, Z.; Wang, Y.; Zhang, C.; Huang, Z.; et al. Molecular Basis of Receptor Binding and Antibody Neutralization of Omicron. Nature 2022, 604, 546–552. [Google Scholar] [CrossRef]
- Nyberg, T.; Ferguson, N.M.; Nash, S.G.; Webster, H.H.; Flaxman, S.; Andrews, N.; Hinsley, W.; Bernal, J.L.; Kall, M.; Bhatt, S.; et al. Comparative Analysis of the Risks of Hospitalisation and Death Associated with SARS-CoV-2 Omicron (B.1.1.529) and Delta (B.1.617.2) Variants in England: A Cohort Study. Lancet 2022, 399, 1303–1312. [Google Scholar] [CrossRef]
- Menni, C.; Valdes, A.M.; Polidori, L.; Antonelli, M.; Penamakuri, S.; Nogal, A.; Louca, P.; May, A.; Figueiredo, J.C.; Hu, C.; et al. Symptom Prevalence, Duration, and Risk of Hospital Admission in Individuals Infected with SARS-CoV-2 during Periods of Omicron and Delta Variant Dominance: A Prospective Observational Study from the ZOE COVID Study. Lancet 2022, 399, 1618–1624. [Google Scholar] [CrossRef]
- Kim, M.K.; Lee, B.; Choi, Y.Y.; Um, J.; Lee, K.S.; Sung, H.K.; Kim, Y.; Park, J.S.; Lee, M.; Jang, H.C.; et al. Clinical Characteristics of 40 Patients Infected With the SARS-CoV-2 Omicron Variant in Korea. J. Korean Med. Sci. 2022, 37, 6–10. [Google Scholar] [CrossRef]
- Wolter, N.; Jassat, W.; Walaza, S.; Welch, R.; Moultrie, H.; Groome, M.; Amoako, D.G.; Everatt, J.; Bhiman, J.N.; Scheepers, C.; et al. Early Assessment of the Clinical Severity of the SARS-CoV-2 Omicron Variant in South Africa: A Data Linkage Study. Lancet 2022, 399, 437–446. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, J.d.P.; Lima, A.B.d.; Alvim, L.B.; Malta, F.S.V.; Mendonça, C.P.T.B.; Carvalho, A.H.B.d.; Rios, J.S.H.; Fonseca, P.L.C.; Queiroz, D.C.; Santos, L.C.G.d.A.e.; et al. Epidemiological Surveillance Reveals the Rise and Establishment of the Omicron SARS-CoV-2 Variant in Brazil. Viruses 2023, 15, 1017. https://doi.org/10.3390/v15041017
Silva JdP, Lima ABd, Alvim LB, Malta FSV, Mendonça CPTB, Carvalho AHBd, Rios JSH, Fonseca PLC, Queiroz DC, Santos LCGdAe, et al. Epidemiological Surveillance Reveals the Rise and Establishment of the Omicron SARS-CoV-2 Variant in Brazil. Viruses. 2023; 15(4):1017. https://doi.org/10.3390/v15041017
Chicago/Turabian StyleSilva, Joice do Prado, Aline Brito de Lima, Luige Biciati Alvim, Frederico Scott Varella Malta, Cristiane Pinheiro Toscano Brito Mendonça, André Henrique Barbosa de Carvalho, Jéssica Silqueira Hickson Rios, Paula Luize Camargos Fonseca, Daniel Costa Queiroz, Luíza Campos Guerra de Araújo e Santos, and et al. 2023. "Epidemiological Surveillance Reveals the Rise and Establishment of the Omicron SARS-CoV-2 Variant in Brazil" Viruses 15, no. 4: 1017. https://doi.org/10.3390/v15041017
APA StyleSilva, J. d. P., Lima, A. B. d., Alvim, L. B., Malta, F. S. V., Mendonça, C. P. T. B., Carvalho, A. H. B. d., Rios, J. S. H., Fonseca, P. L. C., Queiroz, D. C., Santos, L. C. G. d. A. e., Ferreira, A. C. d. S., Souza, R. P. d., Aguiar, R. S. d., & Zauli, D. A. G. (2023). Epidemiological Surveillance Reveals the Rise and Establishment of the Omicron SARS-CoV-2 Variant in Brazil. Viruses, 15(4), 1017. https://doi.org/10.3390/v15041017