
Characterization of a Panel of Cross-Reactive Hantavirus Nucleocapsid Protein-Specific Monoclonal Antibodies

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
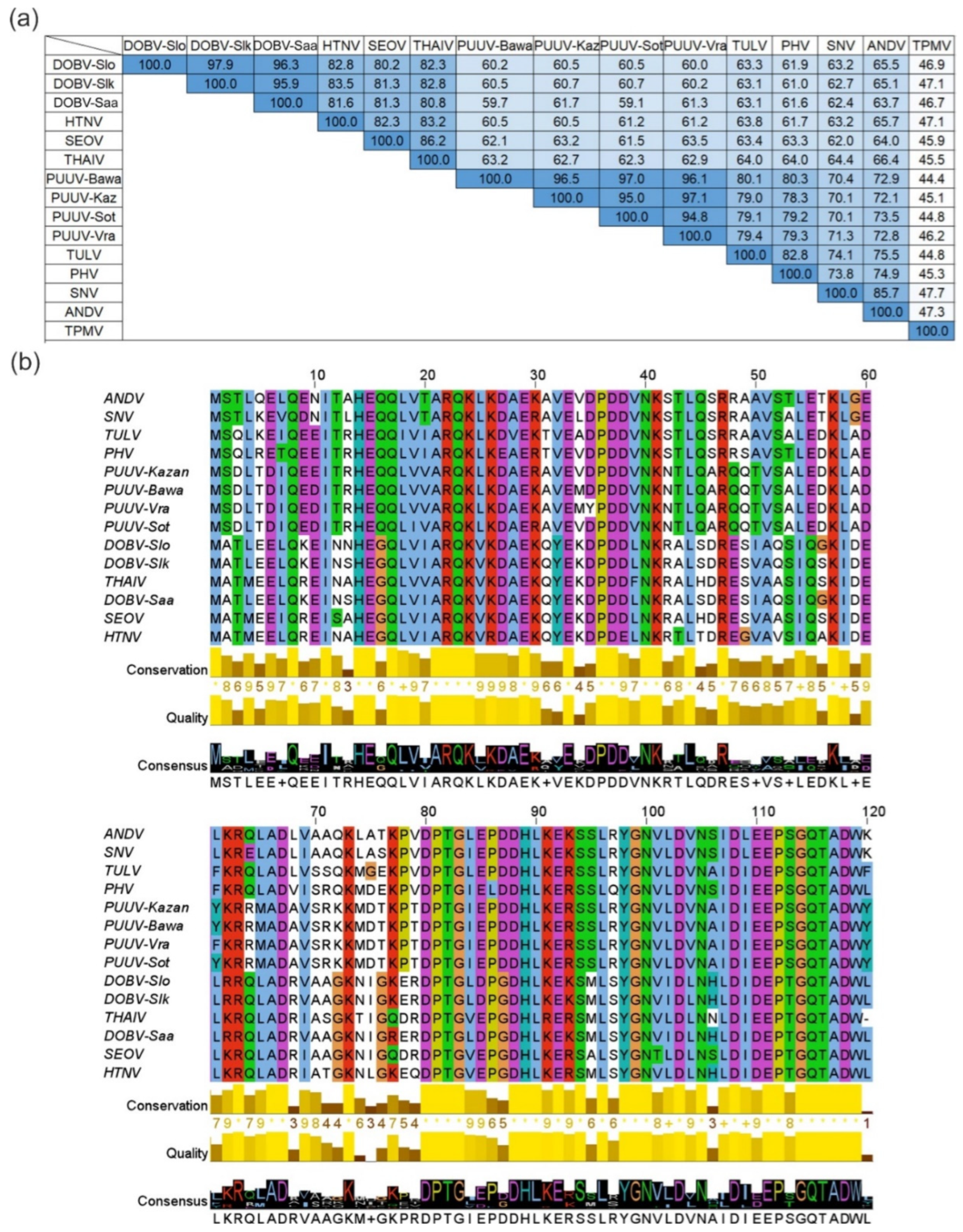
2.1. Bioinformatics Analysis
2.2. Construction of Plasmids

{kind=link}
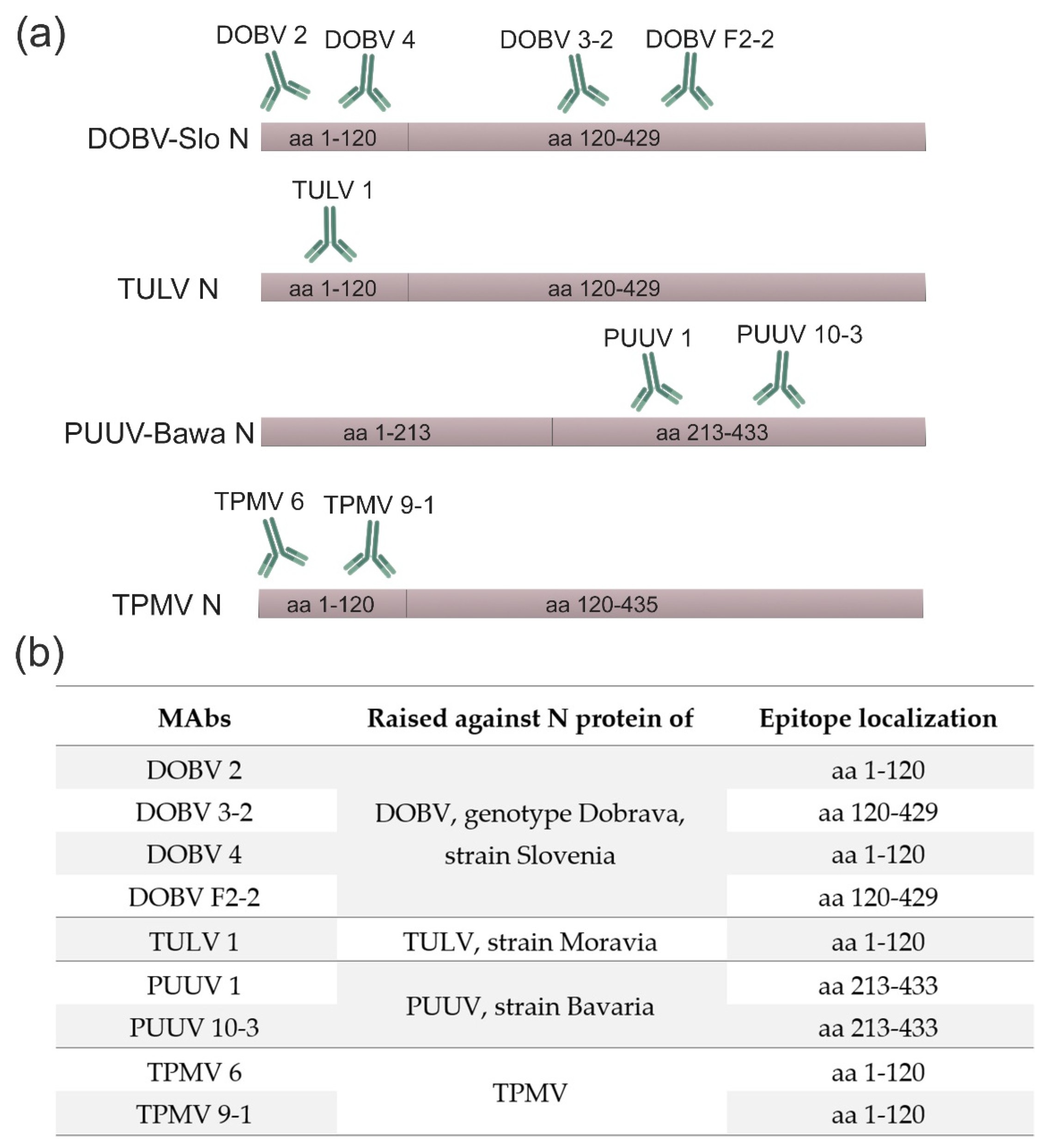{kind=link}
| Origin of the N Protein | GenBank Accession No. | Differences of Published N Protein aa Sequences to Those Used Here a | Reference | ||
|---|---|---|---|---|---|
| Virus Genus | Virus Species, Genotype, Strain | Abbreviation | |||
| Orthohantavirus | Dobrava-Belgrade virus, Kurkino, strain Slovakia | DOBV-Slk | AY533118 | - | [54] |
| Orthohantavirus | Dobrava-Belgrade virus, Dobrava, strain Slovenia | DOBV-Slo | L41916 | - | [54] |
| Orthohantavirus | Dobrava-Belgrade virus, Saaremaa, strain Saar/90Aa/97 | DOBV-Saa | AJ009775 | R77K | [58] |
| Orthohantavirus | Hantaan virus, strain Fojnica | HTNV | M14626 | G377D, A410V | [54] |
| Orthohantavirus | Thailand virus, strain Thai749 | THAIV | AB186420 | - | [59] |
| Orthohantavirus | Seoul virus, strain 80-39 * | SEOV | AY273791 | - | [47,60] |
| Orthohantavirus | Puumala virus, strain Bavaria | PUUV-Bawa | JN696374 | - | [61] |
| Orthohantavirus | Puumala virus, strain Kazan * | PUUV-Kaz | Z84204 | S244C | [54] |
| Orthohantavirus | Puumala virus, strain Sotkamo * | PUUV-Sot | P27313 | - | [54] |
| Orthohantavirus | Puumala virus, strain Vranica/Hällnäs | PUUV-Vra | U14137 | - | [62] |
| Orthohantavirus | Tula virus, strain Moravia | TULV | Z69991 | Y212H, R311A | [47,60] |
| Orthohantavirus | Prospect Hill virus, strain 3571 | PHV | M34011 | T7I, Q97R, E128G, H139Y | [47] |
| Orthohantavirus | Sin Nombre virus, strain 3H226 | SNV | NC_005216 | - | [47,63] |
| Orthohantavirus | Andes virus, strain AH-1 | ANDV | AF004660 | - | [47,64] |
| Thottimvirus | Thottapalayam virus | TPMV | AY526097 | Q20K, A102T, G346S, G357A | [11] |
| Phlebovirus | Rift valley fever virus | RVFV | P21700 | - | [65] |
| Orthobunyavirus | Schmallenberg virus | SBV | HE649914 | - | [66] |
| Tospovirus | Tomato spotted wilt virus | TSWV | NC_002051 | - | [57] |
2.3. Synthesis and Purification of Recombinant N Proteins in S. cerevisiae and E. coli
2.4. Production and Purification of Monoclonal Antibodies
2.5. Indirect ELISA and Determination of the Apparent Dissociation Constants (Kd)
2.6. SDS-PAGE and Western Blot Analysis
2.7. Immunofluorescence Assay (IFA)
3. Results
3.1. Bioinformatics Analysis of Hantavirus N Proteins
3.2. Generation, Initial Characterization and Epitope Mapping of DOBV, TULV, TPMV and PUUV N Protein-Specific MAbs
3.3. Cross-Reactivity Evaluation of Hantavirus-Specific MAbs with Hantavirus N Proteins in ELISA and Western Blot Assay
3.4. Reactivities of Hantavirus-Specific MAbs with Hantavirus-Infected Vero Cells in IFA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Plyusnin, A.; Vapalahti, O.; Vaheri, A. Hantaviruses: Genome structure, expression and evolution. J. Gen. Virol. 1996, 77, 2677–2687. [Google Scholar] [CrossRef] [PubMed]
- Spiropoulou, C.F.; Morzunov, S.; Feldmann, H.; Sanchez, A.; Peters, C.J.; Nichol, S.T. Genome structure and variability of a virus causing hantavirus pulmonary syndrome. Virology 1994, 200, 715–723. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, W.; Sun, Y.; Ma, C.; Wang, X.; Wang, X.; Liu, P.; Shen, S.; Li, B.; Lin, J.; et al. Crystal structure of the core region of hantavirus nucleocapsid protein reveals the mechanism for ribonucleoprotein complex formation. J. Virol. 2016, 90, 1048–1061. [Google Scholar] [CrossRef] [PubMed]
- Yoshimatsu, K.; Arikawa, J. Antigenic properties of N protein of Hantavirus. Viruses 2014, 6, 3097–3109. [Google Scholar] [CrossRef]
- Yoshimatsu, K.; Arikawa, J. Serological diagnosis with recombinant N antigen for hantavirus infection. Virus Res. 2014, 187, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Elgh, F.; Lundkvist, A.; Alexeyev, O.A.; Stenlund, H.; Avsic-Zupanc, T.; Hjelle, B.; Lee, H.W.; Smith, K.J.; Vainionpää, R.; Wiger, D.; et al. Serological diagnosis of hantavirus infections by an enzyme-linked immunosorbent assay based on detection of immunoglobulin G and M responses to recombinant nucleocapsid proteins of five viral serotypes. J. Clin. Microbiol. 1997, 35, 1122–1130. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, I.; Yoshimatsu, K.; Lee, B.H.; Okumura, M.; Taruishi, M.; Araki, K.; Kariwa, H.; Takashima, I.; Arikawa, J. Development of a serotyping ELISA system for Thailand virus infection. Arch. Virol. 2008, 153, 1537–1542. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, S.P.; Yoshimatsu, K.; Koma, T.; Shimizu, K.; Endo, R.; Isozumi, R.; Arikawa, J. Application of truncated nucleocapsid protein (N) for serotyping ELISA of murinae-associated hantavirus infection in rats. Virology 2012, 74, 215–219. [Google Scholar] [CrossRef]
- Koma, T.; Yoshimatsu, K.; Taruishi, M.; Miyashita, D.; Endo, R.; Shimizu, K.; Yasuda, S.P.; Amada, T.; Seto, T.; Murata, R.; et al. Development of a serotyping enzyme-linked immunosorbent assay system based on recombinant truncated hantavirus nucleocapsid proteins for New World hantavirus infection. J. Virol. Methods 2012, 185, 74–81. [Google Scholar] [CrossRef]
- Araki, K.; Yoshimatsu, K.; Ogino, M.; Ebihara, H.; Lundkvist, A.; Kariwa, H.; Takashima, I.; Arikawa, J. Truncated hantavirus nucleocapsid proteins for serotyping Hantaan, Seoul, and Dobrava hantavirus infections. J. Clin. Microbiol. 2001, 39, 2397–2404. [Google Scholar] [CrossRef]
- Schlegel, M.; Tegshduuren, E.; Yoshimatsu, K.; Petraityte, R.; Sasnauskas, K.; Hammerschmidt, B.; Friedrich, R.; Mertens, M.; Groschup, M.H.; Arai, S.; et al. Novel serological tools for detection of Thottapalayam virus, a Soricomorpha-borne hantavirus. Arch. Virol. 2012, 157, 2179–2187. [Google Scholar] [CrossRef]
- Lundkvist, Å.; Meisel, H.; Koletzki, D.; Lankinen, H.; Cifire, F.; Geldmacher, A.; Sibold, C.; Gött, P.; Vaheri, A.; Krüger, D.H. Mapping of B-cell epitopes in the nucleocapsid protein of Puumala hantavirus. Viral Immunol. 2002, 15, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yoshimatsu, K.; Ebihara, H.; Ogino, M.; Araki, K.; Kariwa, H.; Wang, Z.; Luo, Z.; Li, D.; Hang, C.; et al. Genetic diversity of hantaviruses isolated in China and characterization of novel hantaviruses isolated from Niviventer confucianus and Rattus rattus. Virology 2000, 278, 332–345. [Google Scholar] [CrossRef] [PubMed]
- Tischler, N.D.; Rosemblatt, M.; Valenzuela, P.D.T. Characterization of cross-reactive and serotype-specific epitopes on the nucleocapsid proteins of hantaviruses. Virus Res. 2008, 135, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gallo, G.; Kotlik, P.; Roingeard, P.; Monot, M.; Chevreux, G.; Ulrich, R.G.; Tordo, N.; Ermonval, M. Diverse susceptibilities and responses of human and rodent cells to orthohantavirus infection reveal different levels of cellular restriction. PLoS Negl. Trop. Dis. 2022, 16, e0010844. [Google Scholar] [CrossRef]
- Nusshag, C.; Boegelein, L.; Schreiber, P.; Essbauer, S.; Osberghaus, A.; Zeier, M.; Krautkrämer, E. Expression profile of human renal mesangial Cells Is Altered by Infection with Pathogenic Puumala Orthohantavirus. Viruses 2022, 14, 823. [Google Scholar] [CrossRef]
- Holmes, E.C.; Zhang, Y. The evolution and emergence of hantaviruses. Curr. Opin. Virol. 2015, 10, 27–33. [Google Scholar] [CrossRef]
- Yanagihara, R.; Gu, S.H.; Arai, S.; Kang, H.J.; Song, J.W. Hantaviruses: Rediscovery and new beginnings. Virus Res. 2014, 187, 6–14. [Google Scholar] [CrossRef]
- Jiang, H.; Zheng, X.; Wang, L.; Du, H.; Wang, P.; Bai, X. Hantavirus infection: A global zoonotic challenge. Virol. Sin. 2017, 32, 32–43. [Google Scholar] [CrossRef]
- Hierweger, M.M.; Koch, M.C.; Rupp, M.; Maes, P.; Di Paola, N.; Bruggmann, R.; Kuhn, J.H.; Schmidt-Posthaus, H.; Seuberlich, T. Novel Filoviruses, Hantavirus, and Rhabdovirus in freshwater fish, Switzerland, 2017. Emerg. Infect. Dis. 2021, 27, 3082. [Google Scholar] [CrossRef]
- Shi, M.; Lin, X.D.; Chen, X.; Tian, J.H.; Chen, L.J.; Li, K.; Wang, W.; Eden, J.S.; Shen, J.J.; Liu, L.; et al. The evolutionary history of vertebrate RNA viruses. Nature 2018, 556, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Arai, S.; Taniguchi, S.; Aoki, K.; Yoshikawa, Y.; Kyuwa, S.; Tanaka-Taya, K.; Masangkay, J.S.; Omatsu, T.; Puentespina, R.; Watanabe, S.; et al. Molecular phylogeny of a genetically divergent hantavirus harbored by the Geoffroy’s rousette (Rousettus amplexicaudatus), a frugivorous bat species in the Philippines. Infect. Genet. Evol. 2016, 45, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.P.; Lin, X.D.; Wang, W.; Tian, J.H.; Cong, M.L.; Zhang, H.L.; Wang, M.R.; Zhou, R.H.; Wang, J.B.; Li, M.H.; et al. Phylogeny and origins of hantaviruses harbored by bats, insectivores, and rodents. PLoS Pathog. 2013, 9, e1003159. [Google Scholar] [CrossRef]
- Kruger, D.H.; Tadeu, L.; Figueiredo, M.; Song, J.; Klempa, B. Hantaviruses—Globally emerging pathogens. J. Clin. Virol. 2015, 64, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Klempa, B.; Jan, T.A.; Tamara, C.; Heikki, K.D. Complex evolution and epidemiology of Dobrava-Belgrade hantavirus: Definition of genotypes and their characteristics. Arch. Virol. 2013, 158, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Krüger, D.H.; Schönrich, G.; Klempa, B. Human pathogenic hantaviruses and prevention of infection. Hum. Vaccin. 2011, 7, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Plyusnin, A.; Morzunov, S.P. Virus evolution and genetic diversity of hantaviruses and their rodent hosts. Curr. Top. Microbiol. Immunol. 2001, 256, 47–75. [Google Scholar] [CrossRef]
- Song, J.; Baek, L.J.; Schmaljohn, C.S.; Yanagihara, R. Thottapalayam virus, a prototype shrewborne hantavirus. Emerg. Infect. Dis. 2007, 13, 980–985. [Google Scholar] [CrossRef]
- Bennett, S.N.; Gu, S.H.; Kang, H.J.; Arai, S.; Yanagihara, R. Reconstructing the evolutionary origins and phylogeography of hantaviruses. Trends Microbiol. 2014, 22, 473–482. [Google Scholar] [CrossRef]
- Parvate, A.; Williams, E.P.; Taylor, M.K.; Chu, Y.; Lanman, J.; Saphire, E.O.; Jonsson, C.B. Diverse morphology and structural features of Old and New world hantaviruses. Viruses 2019, 11, 862. [Google Scholar] [CrossRef]
- Kang, H.J.; Bennett, N.S.; Sumibcay, L.; Arai, S.; Hope, G.A.; Mocz, G.; Song, J.W.; Cook, A.J.; Yanagihara, R. Evolutionary insights from a genetically divergent hantavirus harbored by the European common mole (Talpa europaea). PLoS ONE 2009, 4, e6149. [Google Scholar] [CrossRef]
- Jonsson, C.B.; Figueiredo, L.T.M.; Vapalahti, O. A global perspective on hantavirus ecology, epidemiology, and disease. Clin. Microbiol. Rev. 2010, 23, 412. [Google Scholar] [CrossRef]
- Klempa, B. Hantaviruses and climate change. Clin. Microbiol. Infect. 2009, 15, 518–523. [Google Scholar] [CrossRef]
- Vapalahti, O.; Mustonen, J.; Lundkvist, Å.; Henttonen, H.; Plyusnin, A.; Vaheri, A. Hantavirus infections in Europe. Lancet Infect. Dis. 2003, 3, 653–661. [Google Scholar] [CrossRef]
- Figueiredo, L.T.M.; Souza, W.M.d.; Ferrés, M.; Enria, D.A. Hantaviruses and cardiopulmonary syndrome in South America. Virus Res. 2014, 187, 43–54. [Google Scholar] [CrossRef]
- Clement, J.; Maes, P.; Lagrou, K.; Van Ranst, M.; Lameire, N. A unifying hypothesis and a single name for a complex globally emerging infection: Hantavirus disease. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1–5. [Google Scholar] [CrossRef]
- Rasmuson, J.; Andersson, C.; Norrman, E.; Haney, M.; Evander, M.; Ahlm, C. Time to revise the paradigm of hantavirus syndromes? Hantavirus pulmonary syndrome caused by European hantavirus. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 30, 685–690. [Google Scholar] [CrossRef]
- Saavedra, F.; Díaz, F.E.; Retamal-Díaz, A.; Covián, C.; González, P.A.; Kalergis, A.M. Immune response during hantavirus diseases: Implications for immunotherapies and vaccine design. Immunology 2021, 163, 262–277. [Google Scholar] [CrossRef]
- Song, J.Y.; Woo, H.J.; Cheong, H.J.; Noh, J.Y.; Baek, L.J.; Kim, W.J. Long-term immunogenicity and safety of inactivated Hantaan virus vaccine (HantavaxTM) in healthy adults. Vaccine 2016, 34, 1289–1295. [Google Scholar] [CrossRef]
- Mittler, E.; Wec, A.Z.; Tynell, J.; Guardado-Calvo, P.; Wigren-Byström, J.; Polanco, L.C.; O’Brien, C.M.; Slough, M.M.; Abelson, D.M.; Serris, A.; et al. Human antibody recognizing a quaternary epitope in the Puumala virus glycoprotein provides broad protection against orthohantaviruses. Sci. Transl. Med. 2022, 14, eabl5399. [Google Scholar] [CrossRef]
- Klingström, J.; Falk, K.I.; Lundkvist, Å. Delayed viremia and antibody responses in Puumala hantavirus challenged passively immunized cynomolgus macaques. Arch. Virol. 2005, 150, 79–92. [Google Scholar] [CrossRef]
- Hepojoki, S.; Rusanen, J.; Hepojoki, J.; Nurmi, V.; Vaheri, A.; Lundkvist, Å.; Hedman, K.; Vapalahti, O. Competitive homogeneous immunoassay for rapid serodiagnosis of hantavirus disease. J. Clin. Microbiol. 2015, 53, 2292–2297. [Google Scholar] [CrossRef]
- Okumura, M.; Yoshimatsu, K.; Kumperasart, S.; Nakamura, I.; Ogino, M.; Taruishi, M.; Sungdee, A.; Pattamadilok, S.; Ibrahim, I.N.; Erlina, S.; et al. Development of serological assays for Thottapalayam virus, an insectivore-borne hantavirus. Clin. Vaccine Immunol. 2007, 14, 173–181. [Google Scholar] [CrossRef]
- Mazzarotto, G.A.C.A.; Raboni, S.M.; Stella, V.; Carstensen, S.; de Noronha, L.; Levis, S.; Zanluca, C.; Zanetti, C.R.; Bordignon, J.; Duarte dos Santos, C.N. Production and characterization of monoclonal antibodies against the recombinant nucleoprotein of Araucaria hantavirus. J. Virol. Methods 2009, 162, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Lundkvist, A.; Bjorsten, S.; Niklasson, B.; Ahlborg, N. Mapping of B-cell determinants in the nucleocapsid protein of Puumala virus: Definition of epitopes specific for acute immunoglobulin G recognition in humans. Clin. Diagn. Lab. Immunol. 1995, 2, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Elgh, F.; Lundkvist, Å.; Alexeyev, O.A.; Wadell, G.; Juto, P. A major antigenic domain for the human humoral response to Puumala virus nucleocapsid protein is located at the amino-terminus. J. Virol. Methods 1996, 59, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Kucinskaite-Kodze, I.; Petraityte-Burneikiene, R.; Zvirbliene, A.; Hjelle, B.; Medina, R.A.; Gedvilaite, A.; Razanskiene, A.; Schmidt-Chanasit, J.; Mertens, M.; Padula, P.; et al. Characterization of monoclonal antibodies against hantavirus nucleocapsid protein and their use for immunohistochemistry on rodent and human samples. Arch. Virol. 2011, 156, 443–456. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Wu, J.; Li, Q.; Wei, Y.; Tan, Z.; Cai, J.; Guo, H.; Yang, L.; Huang, X.; Chen, J.; et al. Seroprevalence, cross antigenicity and circulation sphere of bat-borne hantaviruses revealed by serological and antigenic analyses. PLoS Pathog. 2019, 15, e1007545. [Google Scholar] [CrossRef]
- Zvirbliene, A.; Samonskyte, L.; Gedvilaite, A.; Voronkova, T.; Ulrich, R.; Sasnauskas, K. Generation of monoclonal antibodies of desired specificity using chimeric polyomavirus-derived virus-like particles. J. Immunol. Methods 2006, 311, 57–70. [Google Scholar] [CrossRef] [PubMed]
- BLAST. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 20 February 2022).
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russel, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Razanskiene, A.; Schmidt, J.; Geldmacher, A.; Ritzi, A.; Niedrig, M.; Lundkvist, Å.; Krüger, D.H.; Meisel, H.; Sasnauskas, K.; Ulrich, R. High yields of stable and highly pure nucleocapsid proteins of different hantaviruses can be generated in the yeast Saccharomyces cerevisiae. J. Biotechnol. 2004, 111, 319–333. [Google Scholar] [CrossRef]
- Mertens, M.; Hofmann, J.; Petraityte-Burneikiene, R.; Ziller, M.; Sasnauskas, K.; Friedrich, R.; Niederstrasser, O.; Krüger, D.H.; Groschup, M.H.; Petri, E.; et al. Seroprevalence study in forestry workers of a non-endemic region in eastern Germany reveals infections by Tula and Dobrava-Belgrade hantaviruses. Med. Microbiol. Immunol. 2011, 200, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Cifire, F. Expression und Immunologische Charakterisierung des Nukleokapsidproteins vom Hantavirusisolat Vranica. Ph.D. Dissertation, Freie Universität Berlin, Berlin, Germany, 1997. [Google Scholar]
- de Haan, P.; Wagemakers, L.; Peters, D.; Goldbach, R. The S RNA segment of Tomato spotted wilt virus has an ambisense character. J. Gen. Virol. 1990, 71, 1001–1007. [Google Scholar] [CrossRef] [PubMed]
- Nemirov, K.; Vapalahti, O.; Lundkvist, A.; Vasilenko, V.; Golovljova, I.; Plyusnina, A.; Niemimaa, J.; Laakkonen, J.; Henttonen, H.; Vaheri, A.; et al. Isolation and characterization of Dobrava hantavirus carried by the striped field mouse (Apodemus agrarius) in Estonia. J. Gen. Virol. 1999, 80, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Pattamadilok, S.; Lee, B.H.; Kumperasart, S.; Yoshimatsu, K.; Okumura, M.; Nakamura, I.; Araki, K.; Khoprasert, Y.; Dangsupa, P.; Panlar, P.; et al. Geographical distribution of hantaviruses in Thailand and potential human health significance of Thailand virus. Am. J. Trop. Med. Hyg. 2006, 75, 994–1002. [Google Scholar] [CrossRef]
- Schmidt, J.; Jandrig, B.; Klempa, B.; Yoshimatsu, K.; Arikawa, J.; Meisel, H.; Niedrig, M.; Pitra, C. Nucleocapsid protein of cell culture-adapted Seoul virus strain 80-39: Analysis of its encoding sequence, expression in yeast and immuno-reactivity. Virus Genes 2005, 30, 37–48. [Google Scholar] [CrossRef]
- Mertens, M.; Kindler, E.; Emmerich, P.; Esser, J.; Wagner-Wiening, C.; Wölfel, R.; Petraityte-Burneikiene, R.; Schmidt-Chanasit, J.; Zvirbliene, A.; Groschup, M.H.; et al. Phylogenetic analysis of Puumala virus subtype Bavaria, characterization and diagnostic use of its recombinant nucleocapsid protein. Virus Genes 2011, 43, 177–191. [Google Scholar] [CrossRef]
- Dargeviciute, A.; Sjölander, B.K.; Sasnauskas, K.; Krüger, D.H.; Meisel, H.; Ulrich, R.; Lundkvist, Å.; Brus Sjölander, K.; Sasnauskas, K.; Krüger, D.H.; et al. Yeast-expressed Puumala hantavirus nucleocapsid protein induces protection in a bank vole model. Vaccine 2002, 20, 3523–3531. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.; Meisel, H.; Hjelle, B.; Krüger, D.H.; Ulrich, R. Development and evaluation of serological assays for detection of human hantavirus infections caused by Sin Nombre virus. J. Clin. Virol. 2005, 33, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.; Meisel, H.; Capria, S.G.; Petraityte, R.; Lundkvist, Å.; Hjelle, B.; Vial, P.A.; Padula, P.; Krüger, D.H.; Ulrich, R. Serological assays for the detection of human Andes hantavirus infections based on its yeast-expressed nucleocapsid protein. Intervirology 2006, 49, 173–184. [Google Scholar] [CrossRef]
- Giorgi, C.; Accardi, L.; Nicoletti, L.; Gro, M.C.; Takehara, K.; Hilditch, C.; Morikawa, S.; Bishop, D.H.L. Sequences and coding strategies of the S RNAs of Toscana and Rift Valley fever viruses compared to those of Punta Toro, Sicilian sandfly fever, and Uukuniemi viruses. Virology 1991, 180, 738–753. [Google Scholar] [CrossRef]
- Lazutka, J.; Zvirbliene, A.; Dalgediene, I.; Petraityte-Burneikiene, R.; Spakova, A.; Sereika, V.; Lelesius, R.; Wernike, K.; Beer, M.; Sasnauskas, K.; et al. Generation of recombinant Schmallenberg virus nucleocapsid protein in yeast and development of virus-specific monoclonal antibodies. J. Immunol. Res. 2014, 2014, 160316. [Google Scholar] [CrossRef]
- The QIA Expressionist. A Handbook for High-Level Expression and Purification of 6xHis-Tagged Protein, 5th ed.; Qiagen: Valencia, Spain, 2003; ISBN 8003627737. [Google Scholar]
- Jäckel, S.; Eiden, M.; Dauber, M.; Balkema-Buschmann, A.; Brun, A.; Groschup, M.H. Generation and application of monoclonal antibodies against Rift Valley fever virus nucleocapsid protein NP and glycoproteins Gn and Gc. Arch. Virol. 2014, 159, 535–546. [Google Scholar] [CrossRef]
- De Avila, A.C.; Huguenot, C.; De Resende, O.R.; Kitajima, E.W.; Goldbach, R.W.; Peters, D. Serological differentiation of 20 isolates of tomato spotted wilt virus. J. Gen. Virol. 1990, 71, 2801–2807. [Google Scholar] [CrossRef]
- Špakova, A.; Dalgėdienė, I.; Insodaitė, R.; Sasnauskienė, A.; Žvirblienė, A.; Petraitytė-Burneikienė, R. vB_EcoS_NBD2 bacteriophage-originated polytubes as a carrier for the presentation of foreign sequences. Virus Res. 2020, 290, 198194. [Google Scholar] [CrossRef]
- Sundström, K.B.; Stoltz, M.; Lagerqvist, N.; Lundkvist, Å.; Nemirov, K.; Klingström, J. Characterization of two substrains of Puumala virus that show phenotypes that are different from each other and from the original strain. J. Virol. 2011, 85, 1747. [Google Scholar] [CrossRef] [PubMed]
- Koletzki, D.; Lundkvist, A.; Sjölander, K.B.; Gelderblom, H.R.; Niedrig, M.; Meisel, H.; Krüger, D.H.; Ulrich, R. Puumala (PUU) hantavirus strain differences and insertion positions in the hepatitis B virus core antigen influence B-cell immunogenicity and protective potential of core-derived particles. Virology 2000, 276, 364–375. [Google Scholar] [CrossRef]
- Castel, G.; Chevenet, F.; Razzauti, M.; Murri, S.; Marianneau, P.; Cosson, J.F.; Tordo, N.; Plyusnin, A. Phylogeography of Puumala orthohantavirus in Europe. Viruses 2019, 11, 679. [Google Scholar] [CrossRef]
- Kang, H.J.; Kosoy, M.Y.; Shrestha, S.K.; Shrestha, M.P.; Pavlin, J.A.; Gibbons, R.V.; Yanagihara, R. Genetic diversity of Thottapalayam virus, a hantavirus harbored by the Asian house shrew (Suncus murinus) in Nepal. Am. J. Trop. Med. Hyg. 2011, 85, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Elgh, F.; Linderholm, M.; Wadell, G.; Tärnvik, A.; Juto, P. Development of humoral cross-reactivity to the nucleocapsid protein of heterologous hantaviruses in nephropathia epidemica. FEMS Immunol. Med. Microbiol. 1998, 22, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Okumura, M.; Yoshimatsu, K.; Araki, K.; Lee, B.H.; Asano, A.; Agui, T.; Arikawa, J. Epitope analysis of monoclonal antibody E5/G6, which binds to a linear epitope in the nucleocapsid protein of hantaviruses. Arch. Virol. 2004, 149, 2427–2434. [Google Scholar] [CrossRef] [PubMed]
- Yoshimatsu, K.; Arikawa, J.; Tamura, M.; Yoshida, R.; Lundkvist, A.; Niklasson, B. Characterization of the nucleocapsid protein of Hantaan virus strain 76-118 using monoclonal antibodies. J. Gen. Virol. 1996, 77, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Lundkvist, Å.; Kallio-Kokko, H.; Sjölander, K.B.; Lankinen, H.; Niklasson, B.; Vaheri, A.; Vapalahti, O. Characterization of Puumala virus nucleocapsid protein: Identification of B-cell epitopes and domains involved in protective immunity. Virology 1996, 216, 397–406. [Google Scholar] [CrossRef] [PubMed]


| Recombinant N Proteins | Reactivity of Hantavirus-Specific MAbs | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Novel MAbs | Previously Generated MAbs | ||||||||||||||
| DOBV 2 | DOBV 3-2 | DOBV 4 | DOBV F2-2 | TULV 1 | PUUV 1 | PUUV 10-3 | TPMV 6 | TPMV 9-1 | Against N Proteins of SNV/ANDV | Against PUUV-Vra N Protein Segment (aa 1-120) | |||||
| 4H3 a | 7G2 a | 2C6 b | 5C5 b | 5E11 b | 7A5 b | ||||||||||
| DOBV-Slo | + | ++ | +++ | +++ | + | − | − | − | − | − * | − * | − * | − * | ++ * | − * |
| DOBV-Slk | ++ | ++ | +++ | +++ | + | − | − | − | − | − | − | − | − | ++ | − |
| DOBV-Saa | ++ | +++ | +++ | +++ | + | − | − | − | − | − | − | − | − | ++ | − |
| HTNV | ++ | +++ | +++ | +++ | +++ | − | − | − | − | − * | +++ (+) * | − * | − * | + (−) * | − * |
| SEOV | + | +++ | +++ | +++ | +++ | − | − | − | − | − * | +++ * | − * | + (−) * | +++ * | − * |
| THAIV | +++ | +++ | + | +++ | + | − | − | − | − | − | + | − | − | + | − |
| PUUV-Bawa | ++ | + | − | − | +++ | +++ | +++ | − | − | − | + | − | + | +++ | +++ |
| PUUV-Kaz | ++ | − | − | − | + | +++ | +++ | − | − | − * | + * | − * | + * | +++ * | +++ * |
| PUUV-Sot | +++ | − | − | − | + | +++ | +++ | − | − | − * | ++ * | − * | + * | +++ * | +++ * |
| PUUV-Vra | − | − | − | − | − | +++ | +++ | − | − | − * | ++ * | +++ * | +++ * | +++ * | +++ * |
| TULV | − | − | + | − | +++ | − | − | − | − | − * | ++ * | − * | + * | +++ * | ++ * |
| PHV | ++ | − | − | − | +++ | − | + | − | − | − | +++ | − | + | ++ | − |
| SNV | +++ | + | + | + | +++ | − | − | − | − | +++ * | +++ * | − * | ++ * | ++ * | +++ * |
| ANDV | ++ | +++ | − | +++ | +++ | − | ++ | − | − | +++ * | +++ * | − * | + * | ++ * | +++ * |
| TPMV | − | − | − | − | − | − | − | +++ | +++ | − | − | − | − | − | − |
| SBV | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| RVFV | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| TSWV | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| Recombinant N Proteins | Reactivity of Hantavirus-Specific MAbs | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Novel MAbs | Previously Generated MAbs | ||||||||||||||
| DOBV 2 | DOBV 3-2 | DOBV 4 | DOBV F2-2 | TULV 1 | PUUV 1 | PUUV 10-3 | TPMV 6 | TPMV 9-1 | Against N Proteins of SNV/ANDV | Against PUUV-Vra N Protein Segment (aa 1-120) | |||||
| 4H3 a | 7G2 a | 2C6 b | 5C5 b | 5E11 b | 7A5 b | ||||||||||
| DOBV-Slo | + | + | + | + | + | − | − | − | − | − * | − * | − * | − * | + * | − * |
| DOBV-Slk | + | + | + | + | + | − | − | − | − | − | − | − | − | + | − |
| DOBV-Saa | + | + | + | + | +/− | − | − | − | − | − | − | − | − | + | − |
| HTNV | + | + | + | + | + | − | − | − | − | − * | + * | − * | − * | − * | − * |
| SEOV | + | + | + | + | + | − | − | − | − | − * | + * | − * | + * | + * | − * |
| THAIV | + | + | + | + | + | − | − | − | − | − | + | − | − | + | − |
| PUUV-Bawa | + | − | − | − | + | + | + | − | − | − | + | − | + | + | + |
| PUUV-Kaz | +/− | − | − | − | +/− | + | + | − | − | − * | + * | − * | + * | + * | + * |
| PUUV-Sot | + | − | − | − | + | + | + | − | − | − * | + * | − * | + * | + * | + * |
| PUUV-Vra | − | − | − | − | − | + | + | − | − | − * | + * | + * | + * | + * | + * |
| TULV | − | − | + | − | + | − | − | − | − | − * | + * | − * | + * | + * | + * |
| PHV | + | − | + | − | + | − | − | − | − | − | + | − | + | + | + |
| SNV | + | − | + | − | + | − | − | − | − | + * | + * | − * | + * | + * | + * |
| ANDV | + | + | + | + | + | − | − | − | − | + * | + * | − * | + * | + * | + * |
| TPMV | − | − | − | − | − | − | − | + | + | − | − | − | − | − | − |
| SBV | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| RVFV | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| TSWV | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| Hantaviruses Used in the Test | Reactivity of Hantavirus-Specific MAbs | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Novel MAbs | Previously Generated MAbs | ||||||||||||
| DOBV 2 | DOBV 3-2 | DOBV 4 | DOBV F2-2 | TULV 1 | PUUV 1 | PUUV 10-3 | Against N Proteins of SNV/ANDV | Against PUUV-Vra N Protein Segment (aa 1-120) | |||||
| 4H3 a | 7G2 a | 2C6 b | 5C5 b | 5E11 b | 7A5 b | ||||||||
| DOBV-Slo | ++ | + | ++ | + | ++ | − | − | − | − | − | − | ++ (+) | − |
| DOBV-Saa | ++ | + | ++ | + | + | − | − | − | − | − | − | ++ (+) | − |
| HTNV | +++ | + | +++ | + | +++ | − | − | − | ++ (+) | − | − | − | − |
| SEOV | ++ | + | ++ | + | +++ | − | − | − | + | − | − (+) | ++ (+++) | − |
| PUUV-Kaz | − | − | − | − | ++ | + | + | − | + | +++ | +++ | +++ | + (+++) |
| SNV | +++ | − | ++ | − | +++ | − | − | +++ | ++ | − | +++ (++) | +++ | ++ |
| ANDV | +++ | ++ | ++ | ++ | +++ | − | − | +++ | ++ (−) | − | +++ (+) | +++ (++) | + (−) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avižinienė, A.; Kučinskaitė-Kodzė, I.; Petraitytė-Burneikienė, R.; Žvirblienė, A.; Mertens, M.L.; Schmidt, S.; Schlegel, M.; Lattwein, E.; Koellner, B.; Ulrich, R.G. Characterization of a Panel of Cross-Reactive Hantavirus Nucleocapsid Protein-Specific Monoclonal Antibodies. Viruses 2023, 15, 532. https://doi.org/10.3390/v15020532
Avižinienė A, Kučinskaitė-Kodzė I, Petraitytė-Burneikienė R, Žvirblienė A, Mertens ML, Schmidt S, Schlegel M, Lattwein E, Koellner B, Ulrich RG. Characterization of a Panel of Cross-Reactive Hantavirus Nucleocapsid Protein-Specific Monoclonal Antibodies. Viruses. 2023; 15(2):532. https://doi.org/10.3390/v15020532
Chicago/Turabian StyleAvižinienė, Aliona, Indrė Kučinskaitė-Kodzė, Rasa Petraitytė-Burneikienė, Aurelija Žvirblienė, Marc L. Mertens, Sabrina Schmidt, Mathias Schlegel, Erik Lattwein, Bernd Koellner, and Rainer G. Ulrich. 2023. "Characterization of a Panel of Cross-Reactive Hantavirus Nucleocapsid Protein-Specific Monoclonal Antibodies" Viruses 15, no. 2: 532. https://doi.org/10.3390/v15020532
APA StyleAvižinienė, A., Kučinskaitė-Kodzė, I., Petraitytė-Burneikienė, R., Žvirblienė, A., Mertens, M. L., Schmidt, S., Schlegel, M., Lattwein, E., Koellner, B., & Ulrich, R. G. (2023). Characterization of a Panel of Cross-Reactive Hantavirus Nucleocapsid Protein-Specific Monoclonal Antibodies. Viruses, 15(2), 532. https://doi.org/10.3390/v15020532