
Pet Rats as the Likely Reservoir for Human Seoul Orthohantavirus Infection

 , , , , , , , , , , , and add
Show full author list
, , , , , , , , , , , and add
Show full author list
Abstract
1. Introduction
2. Materials and Methods
2.1. Epidemiological Investigation of the Origin of the Human SEOV Infection in Germany
2.2. Collection of Rats
2.3. Dissection of Rats
2.4. Rat Control Sera
2.5. Screening Strategy for the Different Rat Panels
2.6. ELISA
2.6.1. SEOV-IgG ELISA (A)
2.6.2. Hantavirus-IgG ELISA (B)
2.6.3. SEOV-IgG ELISA (C)
2.7. Line-Blot Analysis
2.8. SEOV RNA Detection and Sequence Determination
2.9. Enrichment of Target Viral Sequences and High-Throughput Sequencing (HTS)
2.10. Sequence Comparison and Phylogenetic Analyses
3. Results
3.1. Identification of the Origin of the Human SEOV Infection
3.2. Analysis of Rats from Patient/Pet-Rat Owner 1 and Pet-Rat Owner 2
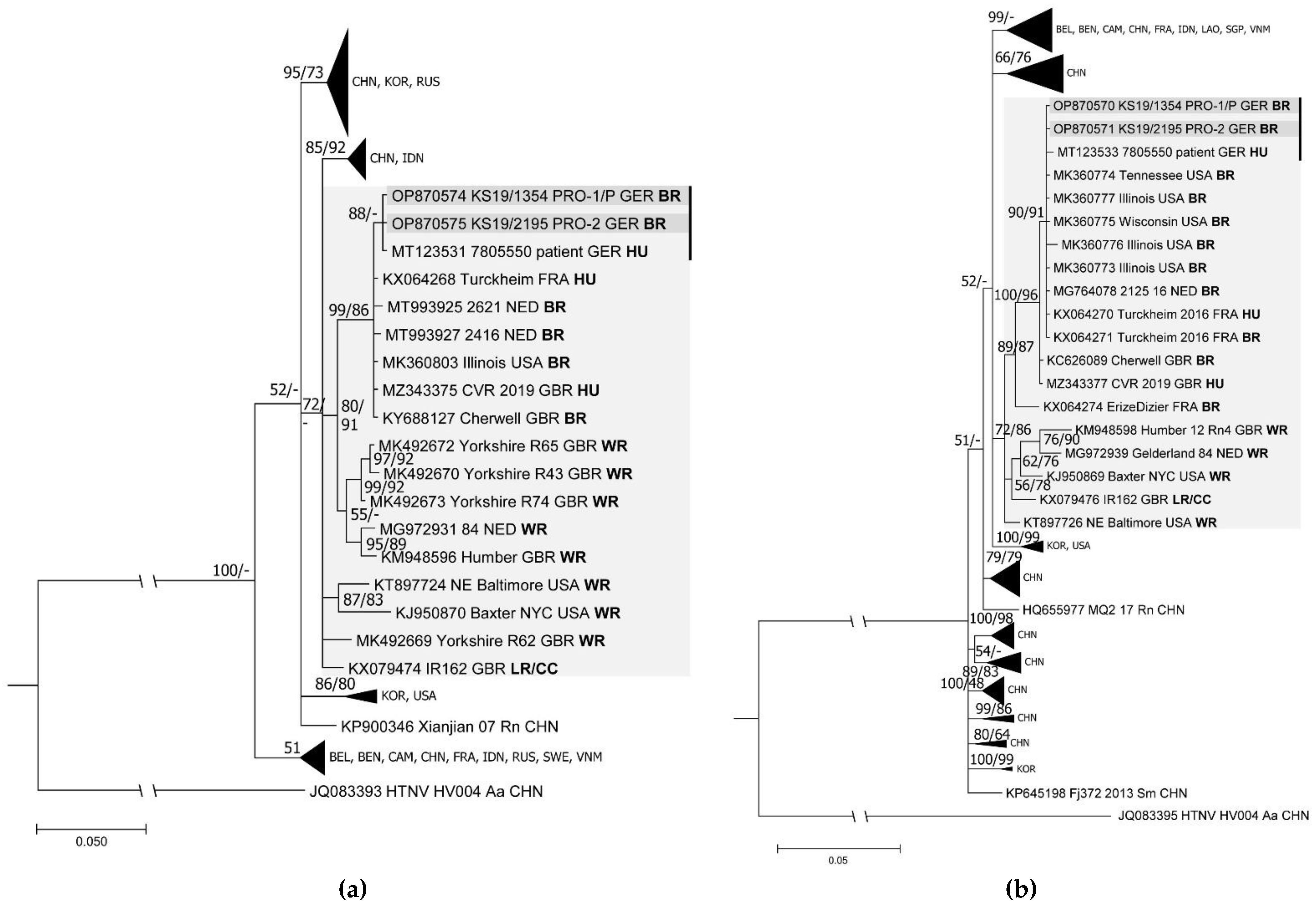
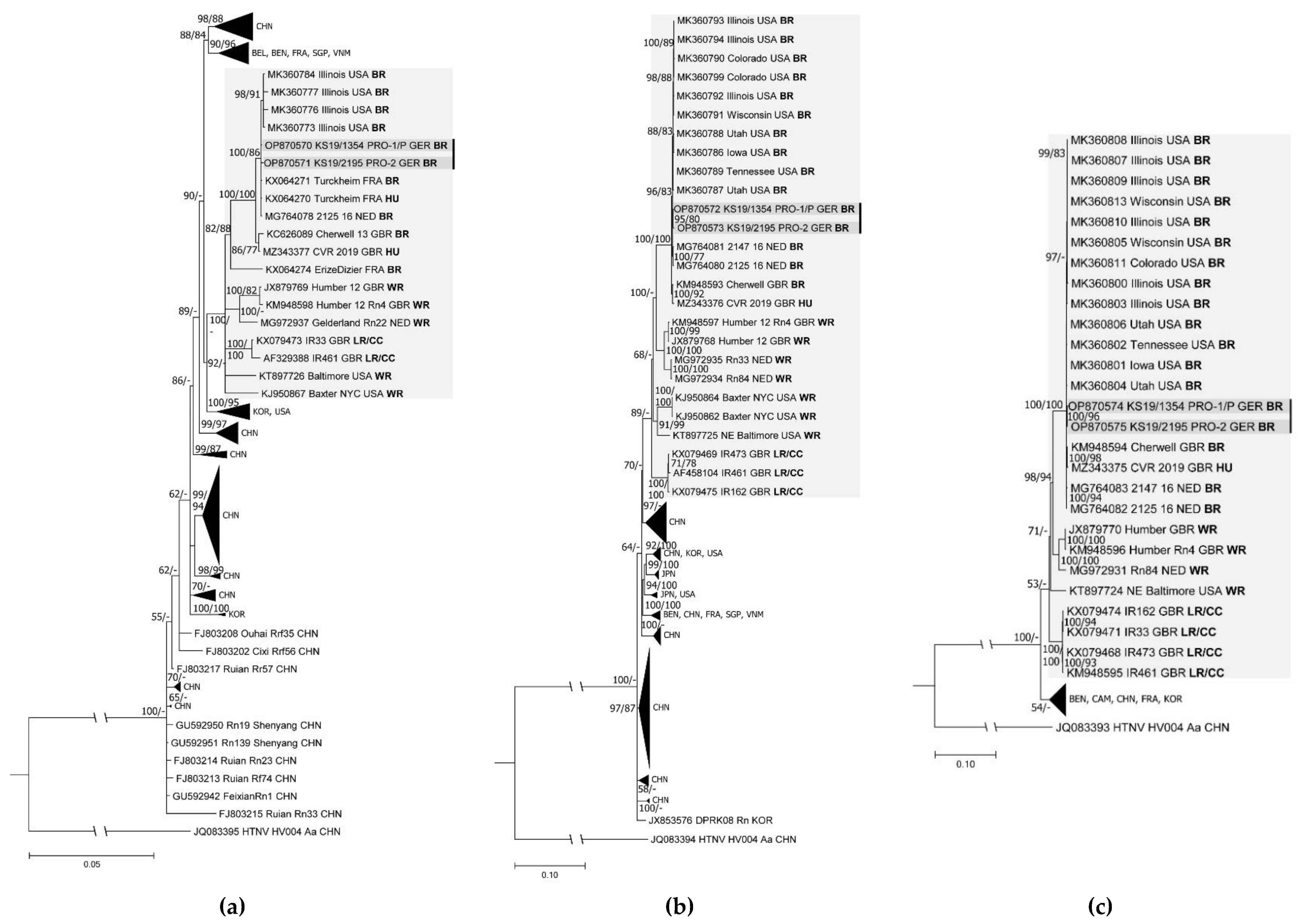
3.3. Complete Genome Identification and Phylogenetic Analysis
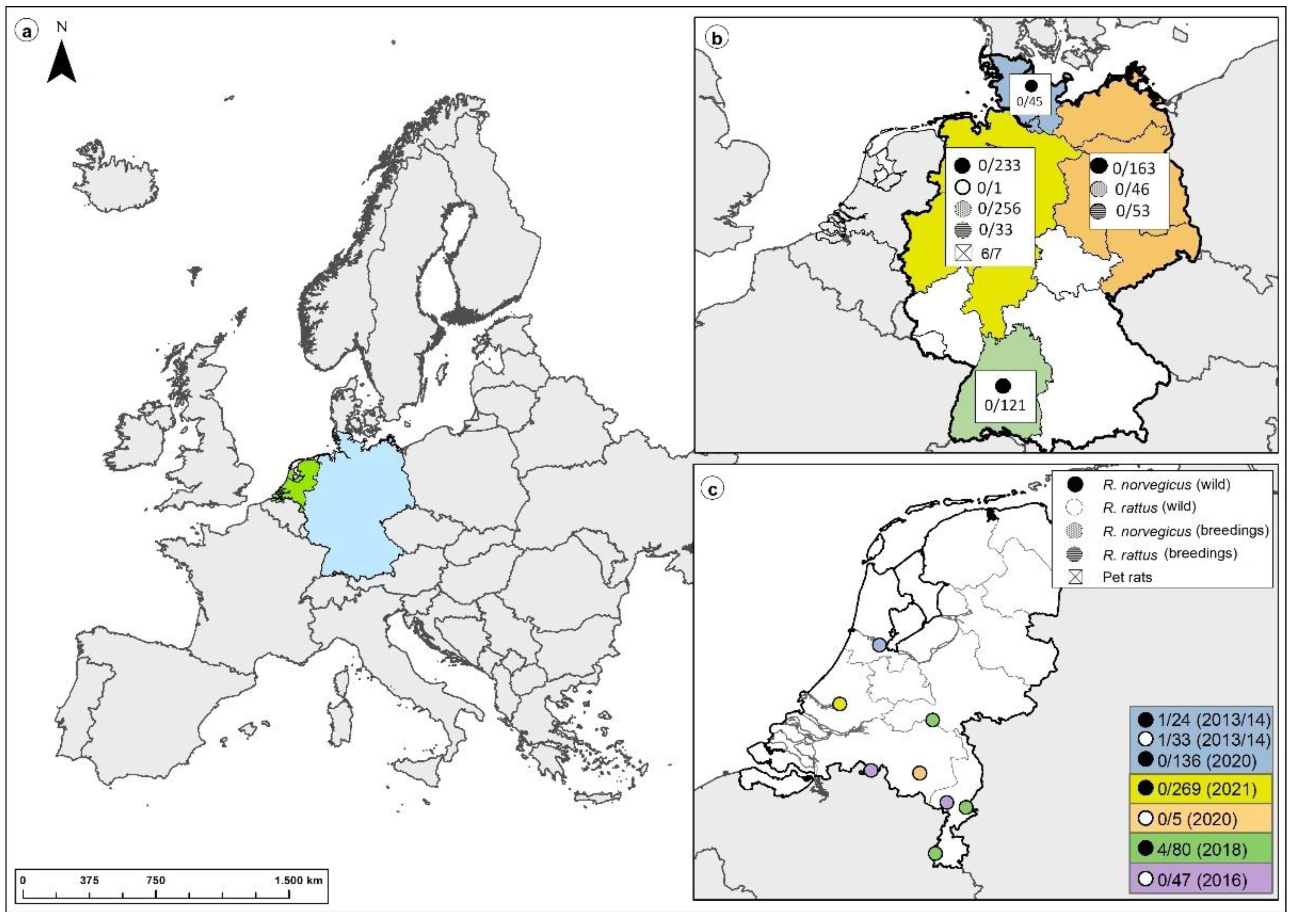
3.4. Large-Scale Screening of Non-Pet Rats from Germany and The Netherlands
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krüger, D.H.; Figueiredo, L.T.M.; Song, J.-W.; Klempa, B. Hantaviruses--globally emerging pathogens. J. Clin. Virol. 2015, 64, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Plyusnin, A.; Vapalahti, O.; Vaheri, A. Hantaviruses: Genome structure, expression and evolution. J. Gen. Virol. 1996, 77 Pt 11, 2677–2687. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W. Hemorrhagic Fever with Renal Syndrome in Korea. Clin. Infect. Dis. 1989, 11, S846–S876. [Google Scholar] [CrossRef]
- Clement, J.; LeDuc, J.W.; McElhinney, L.M.; Reynes, J.M.; Van Ranst, M.; Calisher, C.H. Clinical Characteristics of Ratborne Seoul Hantavirus Disease. Emerg. Infect. Dis. 2019, 25, 387–388. [Google Scholar] [CrossRef] [PubMed]
- Krüger, D.H.; Ulrich, R.G.; Hofmann, J. Hantaviruses as zoonotic pathogens in Germany. Dtsch. Arztebl. Int. 2013, 110, 461–467. [Google Scholar] [CrossRef]
- Lee, H.W.; Baek, L.J.; Johnson, K.M. Isolation of Hantaan virus, the etiologic agent of Korean hemorrhagic fever, from wild urban rats. J. Infect. Dis. 1982, 146, 638–644. [Google Scholar] [CrossRef]
- Hofmann, J.; Weiss, S.; Kuhns, M.; Zinke, A.; Heinsberger, H.; Krüger, D.H. Importation of Human Seoul Virus Infection to Germany from Indonesia. Emerg. Infect. Dis. 2018, 24, 1099–1102. [Google Scholar] [CrossRef]
- Zhang, Y.-Z.; Zou, Y.; Fu, Z.F.; Plyusnin, A. Hantavirus infections in humans and animals, China. Emerg. Infect. Dis. 2010, 16, 1195–1203. [Google Scholar] [CrossRef]
- Lin, X.-D.; Guo, W.-P.; Wang, W.; Zou, Y.; Hao, Z.-Y.; Zhou, D.-J.; Dong, X.; Qu, Y.-G.; Li, M.-H.; Tian, H.-F.; et al. Migration of Norway rats resulted in the worldwide distribution of Seoul hantavirus today. J. Virol. 2012, 86, 972–981. [Google Scholar] [CrossRef]
- Clement, J.; LeDuc, J.W.; Lloyd, G.; Reynes, J.-M.; McElhinney, L.; van Ranst, M.; Lee, H.-W. Wild Rats, Laboratory Rats, Pet Rats: Global Seoul Hantavirus Disease Revisited. Viruses 2019, 11, 652. [Google Scholar] [CrossRef]
- Castel, G.; Kant, R.; Badou, S.; Etougbétché, J.; Dossou, H.-J.; Gauthier, P.; Houéménou, G.; Smura, T.; Sironen, T.; Dobigny, G. Genetic Characterization of Seoul Virus in the Seaport of Cotonou, Benin. Emerg. Infect. Dis. 2021, 27, 2704–2706. [Google Scholar] [CrossRef]
- LeDuc, J.W.; Smith, G.A.; Childs, J.E.; Pinheiro, F.P.; Maiztegui, J.I.; Niklasson, B.; Antoniades, A.; Robinson, D.M.; Khin, M.; Shortridge, K.F.; et al. Global survey of antibody to Hantaan-related viruses among peridomestic rodents*. Bull. World Health Organ. 1986, 64, 139–144. [Google Scholar] [PubMed]
- Shepherd, J.G.; Blunsum, A.E.; Carmichael, S.; Smollett, K.; Maxwell-Scott, H.; Farmer, E.C.W.; Osborne, J.; MacLean, A.; Ashraf, S.; Shah, R.; et al. Seoul Virus Associated with Pet Rats, Scotland, UK, 2019. Emerg. Infect. Dis. 2021, 27, 2677–2680. [Google Scholar] [CrossRef]
- Reynes, J.-M.; Carli, D.; Bour, J.-B.; Boudjeltia, S.; Dewilde, A.; Gerbier, G.; Nussbaumer, T.; Jacomo, V.; Rapt, M.-P.; Rollin, P.E.; et al. Seoul Virus Infection in Humans, France, 2014–2016. Emerg. Infect. Dis. 2017, 23, 973–977. [Google Scholar] [CrossRef]
- McElhinney, L.; Fooks, A.R.; Featherstone, C.; Smith, R.; Morgan, D. Hantavirus (Seoul virus) in pet rats: A zoonotic viral threat. Vet. Rec. 2016, 178, 171–172. [Google Scholar] [CrossRef]
- Swanink, C.; Reimerink, J.; Gisolf, J.; de Vries, A.; Claassen, M.; Martens, L.; Waegemaekers, T.; Rozendaal, H.; Valkenburgh, S.; Hoornweg, T.; et al. Autochthonous Human Case of Seoul Virus Infection, the Netherlands. Emerg. Infect. Dis. 2018, 24, 2158–2163. [Google Scholar] [CrossRef]
- Lundkvist, Å.; Verner-Carlsson, J.; Plyusnina, A.; Forslund, L.; Feinstein, R.; Plyusnin, A. Pet rat harbouring Seoul hantavirus in Sweden, June 2013. Eurosurveillance 2013, 18, 20521. [Google Scholar] [CrossRef]
- Heyman, P.; Baert, K.; Plyusnina, A.; Cochez, C.; Lundkvist, A.; van Esbroeck, M.; Goossens, E.; Vandenvelde, C.; Plyusnin, A.; Stuyck, J. Serological and genetic evidence for the presence of Seoul hantavirus in Rattus norvegicus in Flanders, Belgium. Scand. J. Infect. Dis. 2009, 41, 51–56. [Google Scholar] [CrossRef]
- Murphy, E.G.; Williams, N.J.; Bennett, M.; Jennings, D.; Chantrey, J.; McElhinney, L.M. Detection of Seoul virus in wild brown rats (Rattus norvegicus) from pig farms in Northern England. Vet. Rec. 2019, 184, 525. [Google Scholar] [CrossRef] [PubMed]
- Verner-Carlsson, J.; Lõhmus, M.; Sundström, K.; Strand, T.M.; Verkerk, M.; Reusken, C.; Yoshimatsu, K.; Arikawa, J.; van de Goot, F.; Lundkvist, Å. First evidence of Seoul hantavirus in the wild rat population in the Netherlands. Infect. Ecol. Epidemiol. 2015, 5, 27215. [Google Scholar] [CrossRef] [PubMed]
- Meisel, H.; Wolbert, A.; Razanskiene, A.; Marg, A.; Kazaks, A.; Sasnauskas, K.; Pauli, G.; Ulrich, R.; Krüger, D.H. Development of novel immunoglobulin G (IgG), IgA, and IgM enzyme immunoassays based on recombinant Puumala and Dobrava hantavirus nucleocapsid proteins. Clin. Vaccine Immunol. 2006, 13, 1349–1357. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, R.; Meisel, H.; Schütt, M.; Schmidt, J.; Kunz, A.; Klempa, B.; Niedrig, M.; Pauli, G.; Krüger, D.H.; Koch, J. Verbreitung von Hantavirusinfektionen in Deutschland. Bundesgesundheitsblatt Gesundh. Gesundh. 2004, 47, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, J.; Heuser, E.; Weiss, S.; Tenner, B.; Schoppmeyer, K.; Esser, J.; Klier, C.; Drewes, S.; Ulrich, R.G.; Krüger, D.H. Autochthonous Ratborne Seoul Virus Infection in Woman with Acute Kidney Injury. Emerg. Infect. Dis. 2020, 26, 3096–3099. [Google Scholar] [CrossRef] [PubMed]
- Fawzy, A.; Giel, A.-S.; Fenske, L.; Bach, A.; Herden, C.; Engel, K.; Heuser, E.; Boelhauve, M.; Ulrich, R.G.; Vogel, K.; et al. Development and validation of a triplex real-time qPCR for sensitive detection and quantification of major rat bite fever pathogen Streptobacillus moniliformis. J. Microbiol. Methods 2022, 199, 106525. [Google Scholar] [CrossRef] [PubMed]
- Raafat, D.; Mrochen, D.M.; Al’Sholui, F.; Heuser, E.; Ryll, R.; Pritchett-Corning, K.R.; Jacob, J.; Walther, B.; Matuschka, F.-R.; Richter, D.; et al. Molecular Epidemiology of Methicillin-Susceptible and Methicillin-Resistant Staphylococcus aureus in Wild, Captive and Laboratory Rats: Effect of Habitat on the Nasal S. aureus Population. Toxins 2020, 12, 80. [Google Scholar] [CrossRef]
- Maas, M.; de Vries, A.; Reusken, C.; Buijs, J.; Goris, M.; Hartskeerl, R.; Ahmed, A.; van Tulden, P.; Swart, A.; Pijnacker, R.; et al. Prevalence of Leptospira spp. and Seoul hantavirus in brown rats (Rattus norvegicus) in four regions in the Netherlands, 2011–2015. Infect. Ecol. Epidemiol. 2018, 8, 1490135. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.; Jandrig, B.; Klempa, B.; Yoshimatsu, K.; Arikawa, J.; Meisel, H.; Niedrig, M.; Pitra, C.; Krüger, D.H.; Ulrich, R. Nucleocapsid protein of cell culture-adapted Seoul virus strain 80–39: Analysis of its encoding sequence, expression in yeast and immuno-reactivity. Virus Genes 2005, 30, 37–48. [Google Scholar] [CrossRef]
- Kramski, M.; Meisel, H.; Klempa, B.; Krüger, D.H.; Pauli, G.; Nitsche, A. Detection and typing of human pathogenic hantaviruses by real-time reverse transcription-PCR and pyrosequencing. Clin. Chem. 2007, 53, 1899–1905. [Google Scholar] [CrossRef]
- Hoornweg, T.E.; Zutt, I.; de Vries, A.; Maas, M.; Hoogerwerf, M.N.; Avšič-Županc, T.; Korva, M.; Reimerink, J.H.J.; Reusken, C.B.E.M. Development of a Comparative European Orthohantavirus Microneutralization Assay With Multi- Species Validation and Evaluation in a Human Diagnostic Cohort. Front. Cell. Infect. Microbiol. 2020, 10, 580478. [Google Scholar] [CrossRef]
- Mertens, M.; Wölfel, R.; Ullrich, K.; Yoshimatsu, K.; Blumhardt, J.; Römer, I.; Esser, J.; Schmidt-Chanasit, J.; Groschup, M.H.; Dobler, G.; et al. Seroepidemiological study in a Puumala virus outbreak area in South-East Germany. Med. Microbiol. Immunol. 2009, 198, 83–91. [Google Scholar] [CrossRef]
- Klempa, B.; Fichet-Calvet, E.; Lecompte, E.; Auste, B.; Aniskin, V.; Meisel, H.; Denys, C.; Koivogui, L.; ter Meulen, J.; Krüger, D.H. Hantavirus in African wood mouse, Guinea. Emerg. Infect. Dis. 2006, 12, 838–840. [Google Scholar] [CrossRef] [PubMed]
- Hall, T. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve years of SAMtools and BCFtools. Gigascience 2021, 10, giab008. [Google Scholar] [CrossRef]
- Robinson, J.T.; Thorvaldsdóttir, H.; Wenger, A.M.; Zehir, A.; Mesirov, J.P. Variant Review with the Integrative Genomics Viewer. Cancer Res. 2017, 77, e31–e34. [Google Scholar] [CrossRef]
- Garrison, E.; Marth, G. Haplotype-Based Variant Detection from Short-Read Sequencing. 2012. Available online: http://arxiv.org/pdf/1207.3907v2 (accessed on 17 August 2022).
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; IEEE: New York, NY, USA, 2010; pp. 1–8, ISBN 978-1-4244-9751-5. [Google Scholar]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2--approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Ling, J.; Verner-Carlsson, J.; Eriksson, P.; Plyusnina, A.; Löhmus, M.; Järhult, J.D.; van de Goot, F.; Plyusnin, A.; Lundkvist, Å.; Sironen, T. Genetic analyses of Seoul hantavirus genome recovered from rats (Rattus norvegicus) in the Netherlands unveils diverse routes of spread into Europe. J. Med. Virol. 2019, 91, 724–730. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, G.; Bowen, E.; Jones, N.; Pendry, A. Hfrs Outbreak Associated with Laboratory Rats In UK. The Lancet 1984, 323, 1175–1176. [Google Scholar] [CrossRef]
- Lloyd, G.; Jones, N. Infection of laboratory workers with hantavirus acquired from immunocytomas propagated in laboratory rats. J. Infect. 1986, 12, 117–125. [Google Scholar] [CrossRef]
- Shi, X.; McCaughey, C.; Elliott, R.M. Genetic characterisation of a Hantavirus isolated from a laboratory-acquired infection. J. Med. Virol. 2003, 71, 105–109. [Google Scholar] [CrossRef]
- Knust, B.; Brown, S.; de St Maurice, A.; Whitmer, S.; Koske, S.E.; Ervin, E.; Patel, K.; Graziano, J.; Morales-Betoulle, M.E.; House, J.; et al. Seoul Virus Infection and Spread in United States Home-Based Ratteries: Rat and Human Testing Results From a Multistate Outbreak Investigation. J. Infect. Dis. 2020, 222, 1311–1319. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.L.; Marson, A.L.; Scott, A.L.; Ketner, G.; Glass, G.E. Neonatal sex steroids affect responses to Seoul virus infection in male but not female Norway rats. Brain Behav. Immun. 2002, 16, 736–746. [Google Scholar] [CrossRef]
- Drewes, S.; Ali, H.S.; Saxenhofer, M.; Rosenfeld, U.M.; Binder, F.; Cuypers, F.; Schlegel, M.; Röhrs, S.; Heckel, G.; Ulrich, R.G. Host-Associated Absence of Human Puumala Virus Infections in Northern and Eastern Germany. Emerg. Infect. Dis. 2017, 23, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Cuperus, T.; de Vries, A.; Hoornweg, T.E.; Fonville, M.; Jaarsma, R.I.; Opsteegh, M.; Maas, M. Seoul Virus in Pet and Feeder Rats in The Netherlands. Viruses 2021, 13, 443. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Origin | ID | Sex | Conventional RT-PCR * | Serological Investigations | HTS | Accession Number | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| SEOV S | SEOV M | SEOV L | IgG ELISA SEOV (A) | Hantavirus IgG ELISA (B) | Line-Blot ** | |||||
| Pet-rat owner 1 | KS19/1352 | m | + | + | + | + | + | + | n.d. | OP870576 (S, par) |
| KS19/1353 | m | + | + | - | + | + | + | n.d. | n.a. | |
| KS19/1354 *** | m | + | + | + | + | + | + | spleen, liver | OP870570 (S, CDS); OP870572 (M, CDS); OP870574 (L, CDS) | |
| KS19/1414 | f | - | n.d. | - | - | - | - | n.d. | n.a. | |
| Pet-rat owner 2 | KS19/2195 | m | + | n.d. | n.d. | + | + | + | spleen, liver | OP870571 (S, CDS); OP870573 (M, CDS); OP870575 (L, CDS) |
| KS19/2196 | m | + | n.d. | n.d. | + | + | + | n.d. | n.a. | |
| KS19/2197 | m | - | n.d. | n.d. | - | - | - | n.d. | n.a. | |
| Rat Colony | Number | Species | Hantavirus IgG ELISA (B) ** | Line-Blot Assay | SEOV-S RT-PCR |
|---|---|---|---|---|---|
| A | 39 | R. norvegicus | 0/39 | 0/1 | 0/39 |
| 33 | R. rattus | 0/33 | 0/1 | 0/33 | |
| C | 24 | R. norvegicus | 0/24 | 0/3 | 0/24 |
| 53 | R. rattus | 0/53 | 0/6 | 0/53 | |
| D * | 22 | R. norvegicus | 0/22 | n.d. | 0/22 |
| E | 40 | R. norvegicus | 0/40 | n.d. | 0/40 |
| F | 277 | R. norvegicus | 0/276 | n.d. | 0/277 |
| G | 2 | R. norvegicus | 0/2 | n.d. | 0/2 |
| Total | 490 | 0/489 | 0/11 | 0/490 |
| Region * | Number | Species | Hantavirus IgG ELISA (B) ** | Line-Blot Assay | SEOV-S RT-PCR |
|---|---|---|---|---|---|
| North | 45 | R. norvegicus | 0/33 | 0/4 | 0/45 |
| West | 233 | R. norvegicus | 1 ***/220 | 1 ***/16 | 0/228 |
| 1 | R. rattus | 0/1 | n.d. | 0/1 | |
| South | 121 | R. norvegicus | 0/106 | 0/5 | 0/119 |
| East + Central | 163 | R. norvegicus | 0/150 | 0/9 | 0/152 |
| Total | 563 | 1/510 | 0/34 | 0/545 |
| Study Location (s) | Year of Sample Collection | Location Type | Number | SEOV-IgG ELISA (C) | Real-Time RT-PCR |
|---|---|---|---|---|---|
| Amsterdam | 2013–2014 | Urban | 57 | 2/57 * | 0/2 |
| Municipalities in the provinces of North-Brabant and Limburg | 2016 | Agricultural | 47 | 0/47 | n.d. |
| Appeltern, Roermond, and Maastricht | 2018 | Recreational water | 80 | 4/80 ** | 0/80 |
| Amsterdam | 2020 | Urban | 136 | 0/136 | 0/136 |
| Eindhoven | 2020 | Urban | 5 | 0/5 | 0/5 |
| Rotterdam | 2021 | Urban | 269 | n.d. | 0/269 |
| Total | 594 | 6/325 | 0/492 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heuser, E.; Drewes, S.; Trimpert, J.; Kunec, D.; Mehl, C.; de Cock, M.P.; de Vries, A.; Klier, C.; Oskamp, M.; Tenhaken, P.; et al. Pet Rats as the Likely Reservoir for Human Seoul Orthohantavirus Infection. Viruses 2023, 15, 467. https://doi.org/10.3390/v15020467
Heuser E, Drewes S, Trimpert J, Kunec D, Mehl C, de Cock MP, de Vries A, Klier C, Oskamp M, Tenhaken P, et al. Pet Rats as the Likely Reservoir for Human Seoul Orthohantavirus Infection. Viruses. 2023; 15(2):467. https://doi.org/10.3390/v15020467
Chicago/Turabian StyleHeuser, Elisa, Stephan Drewes, Jakob Trimpert, Dusan Kunec, Calvin Mehl, Marieke P. de Cock, Ankje de Vries, Christiane Klier, Martin Oskamp, Peter Tenhaken, and et al. 2023. "Pet Rats as the Likely Reservoir for Human Seoul Orthohantavirus Infection" Viruses 15, no. 2: 467. https://doi.org/10.3390/v15020467
APA StyleHeuser, E., Drewes, S., Trimpert, J., Kunec, D., Mehl, C., de Cock, M. P., de Vries, A., Klier, C., Oskamp, M., Tenhaken, P., Hashemi, F., Heinz, D., Nascimento, M., Boelhauve, M., Petraityte-Burneikiene, R., Raafat, D., Maas, M., Krüger, D. H., Latz, A., ... Ulrich, R. G. (2023). Pet Rats as the Likely Reservoir for Human Seoul Orthohantavirus Infection. Viruses, 15(2), 467. https://doi.org/10.3390/v15020467