
Coinfection with Yellow Head Virus Genotype 8 (YHV-8) and Oriental Wenrivirus 1 (OWV1) in Wild Penaeus chinensis from the Yellow Sea
 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Information
2.2. DNA and RNA Extraction
2.3. Pathogens Testing with Pooled Samples
2.4. YHVs and OWV1 Testing with Individual Samples
2.5. Transmission Electron Microscopy
2.6. Virus Purification
2.7. Phylogenetic Analyses
3. Results
3.1. Testing Results of Pooled Samples
3.2. Testing Results of Individual Samples for YHVs and OWV1
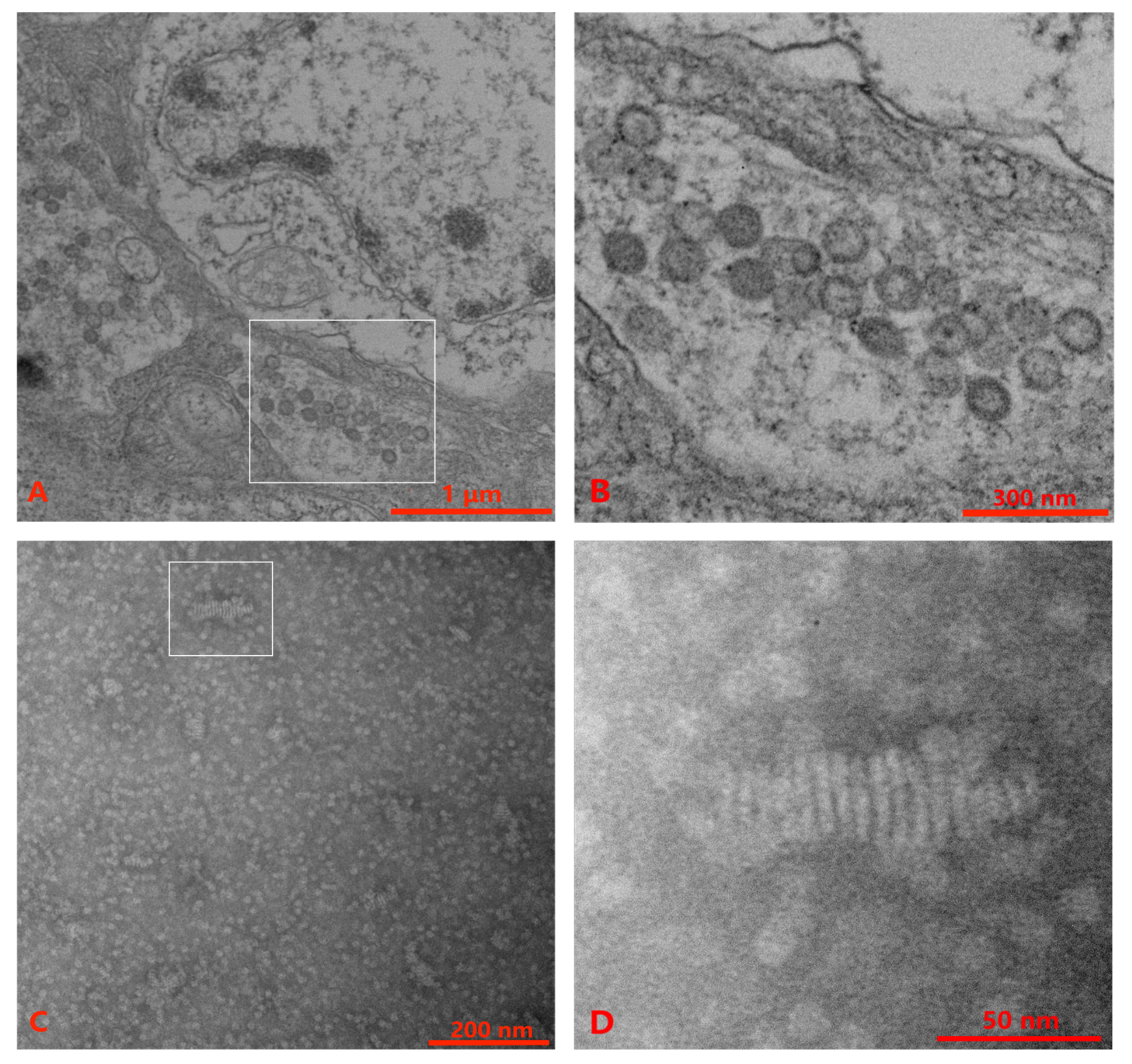
3.3. TEM Examination
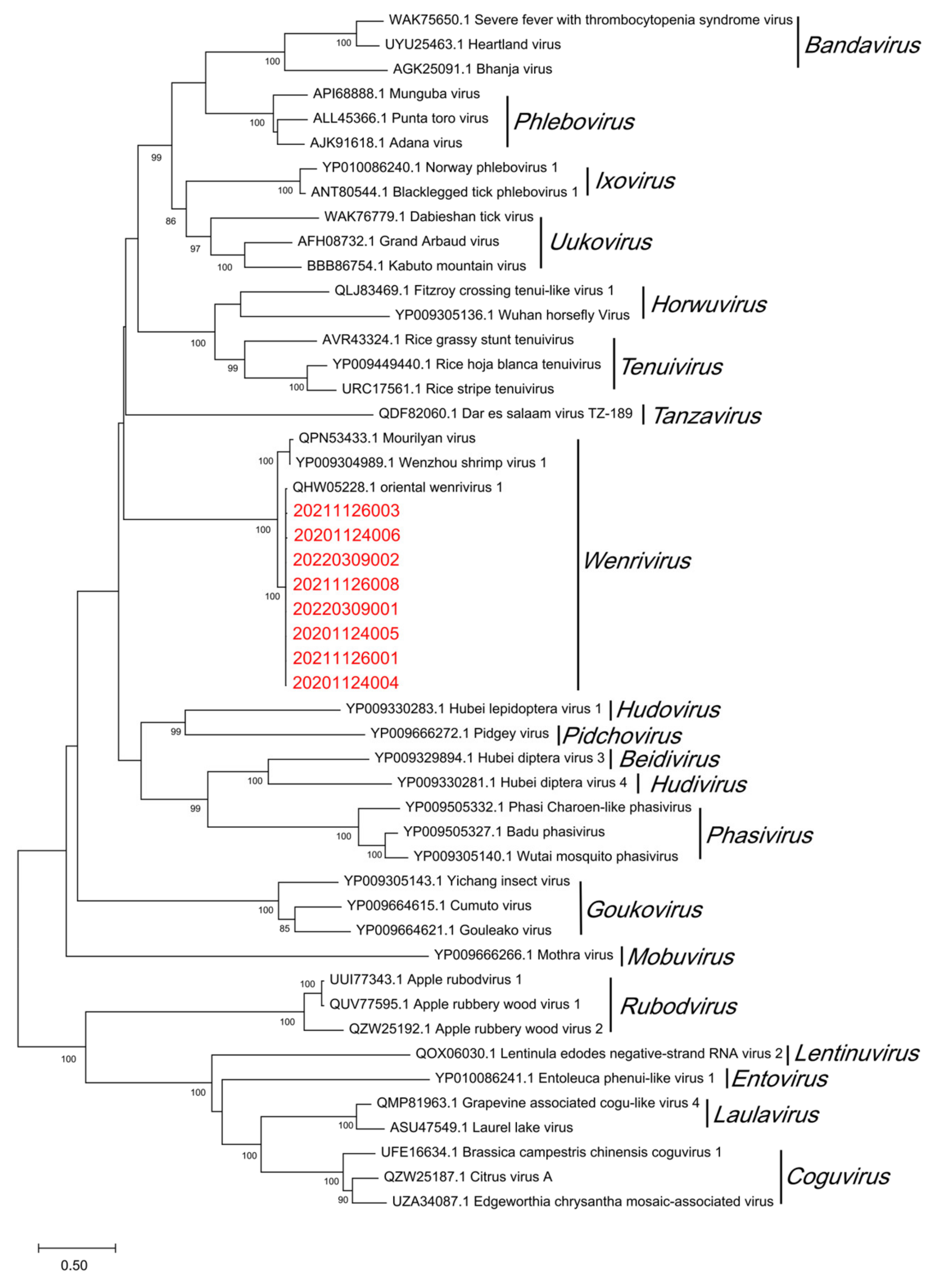
3.4. Homology and Phylogenetic Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Qiu, L.; Chen, X.; Zhao, R.H.; Li, C.; Gao, W.; Zhang, Q.L.; Huang, J. Description of a natural infection with decapod iridescent virus 1 in farmed giant freshwater prawn, Macrobrachium rosenbergii. Viruses 2019, 11, 354. [Google Scholar] [CrossRef]
- Benzie, J.A.H. Use and exchange of genetic resources of penaeid shrimps for food and aquaculture. Rev. Aquac. 2009, 1, 232–250. [Google Scholar] [CrossRef]
- Wang, Q.; Zhuang, Z.; Deng, J.; Ye, Y. Stock enhancement and translocation of the shrimp Penaeus chinensis in China. Fish. Res. 2006, 80, 67–79. [Google Scholar] [CrossRef]
- Arbon, P.M.; Condon, K.; Andrade Martinez, M.; Jerry, D.R. Molecular detection of six viral pathogens from Australian wild sourced giant black tiger shrimp (Penaeus monodon) broodstock. Aquaculture 2022, 548, 737651. [Google Scholar] [CrossRef]
- Gholamhosseini, A.; Mohammadi, A.; Akbari, S.; Banaee, M. Molecular, histopathologic and electron microscopic analysis of white spot syndrome virus in wild shrimp (Fenneropenaeus indicus) in the coastal waters of Iran. Arch. Virol. 2020, 165, 1433–1440. [Google Scholar] [CrossRef] [PubMed]
- Srisala, J.; Sanguanrut, P.; Thaiue, D.; Laiphrom, S.; Siriwattano, J.; Khudet, J.; Thaiue, D.; Powtongsook, S.; Flegel, T.W.; Sritunyalucksana, K. Infectious myonecrosis virus (IMNV) and decapod iridescent virus 1 (DIV1) detected in captured, wild Penaeus monodon from the Indian Ocean. Aquaculture 2021, 545, 737262. [Google Scholar] [CrossRef]
- Hamano, K.; Maeno, Y.; Klomkling, S.; Aue-Umneoy, D.; Tsutsui, I. Presence of viral pathogens amongst wild Penaeus monodon in Thailand. Jpn. Agric. Res. Q. 2017, 51, 191–197. [Google Scholar] [CrossRef]
- Kotob, M.H.; Gorgoglione, B.; Kumar, G.; Abdelzaher, M.; Saleh, M.; El-Matbouli, M. The impact of Tetracapsuloides bryosalmonae and Myxobolus cerebralis co-infections on pathology in rainbow trout. Parasites Vectors 2017, 10, 442. [Google Scholar] [CrossRef]
- Tang, K.F.J.; Durand, S.V.; White, B.L.; Redman, R.M.; Mohney, L.L.; Lightner, D.V. Induced resistance to white spot syndrome virus infection in Penaeus stylirostris through pre-infection with infectious hypodermal and hematopoietic necrosis virus—A preliminary study. Aquaculture 2003, 216, 19–29. [Google Scholar] [CrossRef]
- Kotob, M.H.; Kumar, G.; Saleh, M.; Gorgoglione, B.; Abdelzaher, M.; El-Matbouli, M. Differential modulation of host immune genes in the kidney and cranium of the rainbow trout (Oncorhynchus mykiss) in response to Tetracapsuloides bryosalmonae and Myxobolus cerebralis co-infections. Parasites Vectors 2018, 11, 326. [Google Scholar] [CrossRef]
- Saravanan, K.; Praveenraj, J.; Kiruba-Sankar, R.; Devi, V.; Biswas, U.; Kumar, T.S.; Sudhagar, A.; El-Matbouli, M.; Kumar, G. Co-Infection of infectious hypodermal and hematopoietic necrosis virus (IHHNV) and white spot syndrome virus (WSSV) in the wild crustaceans of Andaman and Nicobar Archipelago, India. Viruses 2021, 13, 1378. [Google Scholar] [CrossRef]
- Lightner, D.V. The penaeid shrimp viral pandemics due to IHHNV, WSSV, TSV and YHV: History in the Americas and current status. In Proceedings of the 32nd Joint Meeting of the United States-Japan Cooperative Program in Natural Resource, Davis and Santa Barbara, CA, USA, 17, 18 and 20 November 2003. [Google Scholar]
- Morales-Covarrubias, M.S.; Nunan, L.M.; Lightner, D.V.; Mota-Urbina, J.C.; Garza-Aguirre, M.C.; Chávez-Sánchez, M.C. Prevalence of infectious hypodermal and hematopoietic necrosis virus (IHHNV) in wild adult blue shrimp Penaeus stylirostris from the Northern Gulf of California, Mexico. J. Aquat. Anim. Health 1999, 11, 296–301. [Google Scholar] [CrossRef]
- Walker, P.J.; Cowley, J.A.; Dong, X.; Huang, J.; Moody, N.; Ziebuhr, J.; ICTV Report Consortium. ICTV Virus Taxonomy Profile: Roniviridae. J. Gen. Virol. 2020, 102, jgv001514. [Google Scholar] [CrossRef]
- WOAH. Manual of Diagnostic Tests for Aquatic Animals. World Organisation for Aquatic Animal Health. Available online: https://www.woah.org/en/what-we-do/standards/codes-and-manuals/aquatic-manual-online-access/ (accessed on 9 January 2023).
- Walker, P.J.; Cowley, J.A.; Spann, K.; Hodgson, R.; Hall, M.; Withyachumnarnkul, B. Yellow Head Complex Viruses: Transmission Cycles and Topographical Distribution in the Asia-Pacific Region; World Aquaculture Society: Baton Rouge, LA, USA, 2001; pp. 292–302. Available online: http://hdl.handle.net/102.100.100/201029?index=1 (accessed on 18 January 2023).
- Wijegoonawardane, P.K.M.; Cowley, J.A.; Phan, T.; Hodgson, R.A.J.; Nielsen, L.; Kiatpathomchai, W.; Walker, P.J. Genetic diversity in the yellow head nidovirus complex. Virology 2008, 380, 213–225. [Google Scholar] [CrossRef]
- Chen, J.Y.; Wang, W.C.; Wang, X.H.; Zhang, Q.L.; Ren, Y.B.; Song, J.P.; Wang, X.P.; Dong, X.; Huang, J. First detection of yellow head virus genotype 3 (YHV-3) in cultured Penaeus monodon, mainland China. J. Fish Dis. 2018, 41, 1449–1451. [Google Scholar] [CrossRef]
- Mohr, P.G.; Moody, N.J.; Hoad, J.G.; Williams, L.M.; Bowater, R.O.; Cummins, D.M.; Cowley, J.A.; Crane, M.S. New yellow head virus genotype (YHV7) in giant tiger shrimp Penaeus monodon indigenous to northern Australia. Dis. Aquat. Organ. 2015, 115, 263–268. [Google Scholar] [CrossRef]
- Liu, Q.; Huang, J.; Yang, H.L.; Yang, B.; Liu, S.; Wang, H.L.; Wang, Q.T.; Liu, F.; Zhang, Q.L. Detection of a new genotype of yellow head virus in farmed shrimp suspicious of EMS/AHPNS infection. Oceanol. Limnol. Sin. 2014, 45, 703–709. [Google Scholar] [CrossRef]
- Zhu, L.L.; Zhang, Q.L.; Wan, X.Y.; Qiu, L.; Huang, J. Molecular epidemiology of a new yellow head virus strain in China. Prog. Fish. Sci. 2016, 37, 68–77. [Google Scholar] [CrossRef]
- Dong, X.; Liu, S.; Zhu, L.L.; Wan, X.Y.; Liu, Q.; Qiu, L.; Zhou, P.Z.; Zhang, Q.L.; Huang, J. Complete genome sequence of an isolate of a novel genotype of yellow head virus from Fenneropenaeus chinensis indigenous in China. Arch. Virol. 2017, 162, 1149–1152. [Google Scholar] [CrossRef]
- Kim, S.R.; Cwr, G.; Shmp, W.; Shin, G.W. Detection and genetic characteristic of Yellow-head virus genotype 8 (YHV-8) cultured Litopanaeus vanamei, in Korea. J. Fish Pathol. 2020, 33, 77–81. [Google Scholar] [CrossRef]
- Dong, X.; Hu, T.; Ren, Y.b.; Meng, F.Z.; Li, C.; Zhang, Q.L.; Chen, J.Y.; Song, J.P.; Wang, R.Y.; Shi, M.; et al. A novel bunyavirus discovered in oriental shrimp (Penaeus chinensis). Front. Microbiol. 2021, 12, 751112. [Google Scholar] [CrossRef]
- Lightner, D.V.; Redman, R.M.; Bell, T.A. Infectious hypodermal and hematopoietic necrosis, a newly recognized virus disease of penaeid shrimp. J. Invertebr. Pathol. 1983, 42, 62–70. [Google Scholar] [CrossRef]
- Bell, T.A.; Lightner, D.V. IHHN virus: Infectivity and pathogenicity studies in Penaeus styliostris and Penaeus vannamei. Aquaculture 1984, 38, 185–194. [Google Scholar] [CrossRef]
- Kalagayan, H.; Godin, D.; Kanna, R.; Hagino, G.; Sweeney, J.; Wyban, J.; James, B. IHHN virus as an etiological factor in runt-deformity syndrome of juvenile Penaeus vannamei culturted in Hawaii. World Aquac. Soc. 1991, 22, 235–243. [Google Scholar] [CrossRef]
- Primavera, J.H.; Quinitio, E.T. Runt-deformity syndrome in cultured fiant tiger prawn Penaeus monodon. J. Crustac. Biol. 2000, 20, 796–802. [Google Scholar] [CrossRef]
- Lotz, J.M. Viruses, biosecurity and specific pathogen-free stocks in shrimp aquaculture. World J. Microbiol. Biotechnol. 1997, 13, 405–413. [Google Scholar] [CrossRef]
- ICTV. Iridoviridae, Subfamily: Betairidovirinae, Genus: Decapodiridovirus. The ICTV Report on Virus Classification and Taxon Nomenclature. Available online: https://ictv.global/report/chapter/iridoviridae/iridoviridae/decapodiridovirus (accessed on 9 January 2023).
- Qiu, L.; Chen, M.M.; Wan, X.Y.; Li, C.; Zhang, Q.L.; Wang, R.Y.; Cheng, D.Y.; Dong, X.; Yang, B.; Wang, X.H.; et al. Characterization of a new member of Iridoviridae, Shrimp hemocyte iridescent virus (SHIV), found in white leg shrimp (Litopenaeus vannamei). Sci. Rep. 2017, 7, 11834. [Google Scholar] [CrossRef]
- Sun, W.F.; Huang, X.S.; Hu, X.J.; Wen, G.L.; Cao, Y.C.; Zhang, J.S. Detection and analysis of Enterocytozoon hepatopenaei (EHP), Vibrio parahaemolyticus acutehepatopancreatic necrosis disease (VPAHPND) and shrimp hemocyte iridescent virus (SHIV) from Litopenaeus vannamei in coastal areas of Guangdong Province. J. South. Agric. 2019, 50, 2343–2349. [Google Scholar]
- Qiu, L.; Chen, X.; Gao, W.; Li, C.; Guo, X.M.; Zhang, Q.L.; Yang, B.; Huang, J. Molecular epidemiology and histopathological study of a natural infection with decapod iridescent virus 1 in farmed white leg shrimp, Penaeus vannamei. Aquaculture 2021, 533, 736105. [Google Scholar] [CrossRef]
- Lo, C.F.; Leu, J.H.; Ho, C.H.; Chen, C.H.; Peng, S.E.; Chen, Y.T.; Chou, C.M.; Yeh, P.Y.; Huang, C.J.; Chou, H.Y. Detection of baculovirus associated with white spot syndrome (WSBV) in penaeid shrimps using polymerase chain reaction. Dis. Aquat. Org. 1996, 24, 133–141. [Google Scholar] [CrossRef]
- Durand, S.V.; Lightner, D.V. Quantitative real time PCR for the measurement of white spot syndrome virus in shrimp. J. Fish Dis. 2002, 25, 381–389. [Google Scholar] [CrossRef]
- Dhar, A.K.; Roux, M.M.; Klimpel, K.R. Detection and quantification of infectious hypodermal and hematopoietic necrosis virus and white spot virus in shrimp using real-time quantitative PCR and SYBR green chemistry. J. Clin. Microbiol. 2001, 39, 2835–2845. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.M.; Qiu, L.; Sheng, A.Z.; Wan, X.Y.; Cheng, D.Y.; Huang, J. Quantitative detection method of Enterocytozoon hepatopenaei using TaqMan probe real-time PCR. J. Invertebr. Pathol. 2018, 151, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.F.J.; Durand, S.V.; White, B.L.; Redman, R.M.; Pantoja, C.R.; Lightner, D.V. Postlarvae and juveniles of a selected line of Penaeus stylirostris are resistant to infectious hypodermal and hematopoietic necrosis virus infection. Aquaculture 2000, 190, 203–210. [Google Scholar] [CrossRef]
- Jaroenlak, P.; Sanguanrut, P.; Williams, B.A.P.; Stentiford, G.D.; Flegel, T.W.; Sritunyalucksana, K.; Itsathitphaisarn, O. A Nested PCR assay to avoid false positive detection of the microsporidian Enterocytozoon hepatopenaei (EHP) in environmental samples in shrimp farms. Public Libr. Sci. 2016, 11, e0166320. [Google Scholar] [CrossRef] [PubMed]
- Andrade, T.P.D.; Srisuvan, T.; Tang, K.F.J.; Lightner, D.V. Real-time reverse transcription polymerase chain reaction assay using TaqMan probe for detection and quantification of infectious myonecrosis virus (IMNV). Aquaculture 2007, 264, 9–15. [Google Scholar] [CrossRef]
- Senapin, S.; Phewsaiya, K.; Briggs, M.; Flegel, T.W. Outbreaks of infectious myonecrosis virus (IMNV) in Indonesia confirmed by genome sequencing and use of an alternative RT-PCR detection method. Aquaculture 2007, 266, 32–38. [Google Scholar] [CrossRef]
- Li, X.P.; Wan, X.Y.; Xu, T.T.; Huang, J.; Zhang, Q.L. Development and validation of a TaqMan RT-qPCR for the detection of convert mortality nodavirus (CMNV). J. Virol. Methods 2018, 262, 65–71. [Google Scholar] [CrossRef]
- Poulos, B.T.; Lightner, D.V. Detection of infectious myonecrosis virus (IMNV) of penaeid shrimp by reverse-transcriptase polymerase chain reaction (RT-PCR). Dis. Aquat. Org. 2006, 73, 69–72. [Google Scholar] [CrossRef]
- Zhang, Q.L.; Liu, Q.; Liu, S.; Yang, H.L.; Liu, S.; Zhu, L.L.; Yang, B.; Jin, J.T.; Ding, L.X.; Wang, X.H.; et al. A new nodavirus is associated with covert mortality disease of shrimp. J. Gen. Virol. 2014, 95, 2700–2709. [Google Scholar] [CrossRef]
- Navarro, S.A.; Tang, K.F.J.; Lightner, D.V. An improved Taura syndrome virus (TSV) RT-PCR using newly designed primers. Aquaculture 2009, 293, 290–292. [Google Scholar] [CrossRef]
- Tang, K.F.J.; Wang, J.; Lightner, D.V. Quantitation of Taura syndrome virus by real-time RT-PCR with a TaqMan assay. J. Virol. Methods 2004, 115, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Chen, X.; Guo, X.M.; Gao, W.; Huang, J. A TaqMan probe based real-time PCR for the detection of Decapod iridescent virus 1. J. Invertebr. Pathol. 2020, 173, 107367. [Google Scholar] [CrossRef]
- Cowley, J.A.; Cadogan, L.C.; Wongteerasupaya, C.; Hodgson, R.; Boonsaeng, V.; Walker, P.J. Multiplex RT-nested PCR differentiation of gill-associated virus (Australia) from yellow head virus (Thailand) of Penaeus monodon. J. Virol. Methods 2004, 117, 49–59. [Google Scholar] [CrossRef]
- Li, C.; Ren, Y.B.; Dong, X.; Wang, C.M.; Huang, J. Extraction of assembling complexes of viral capsomers from shrimp tissue infected with yellow head virus genotype 8 (YHV-8). J. Fish Dis. 2019, 42, 613–616. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Koichiro, T.; Glen, S.; Sudhir, K. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Dhar, A.K.; Cowley, J.A.; Hasson, K.W.; Walker, P.J. Genomic organization, biology, and diagnosis of Taura syndrome virus and yellow head virus of penaeid shrimp. Adv. Virus Res. 2004, 63, 353–421. [Google Scholar] [CrossRef]
- Ossiander, F.J.; Wedemeyer, G. Computer program for sample sizes required to determine disease incidence in fish populations. J. Fish. Res. Board Can. 1973, 30, 1383–1384. [Google Scholar] [CrossRef]
- Song, Z.L.; Dong, X.; Zhao, R.H.; Wang, X.H.; Wu, H.Y.; Yu, D.H.; Xie, G.S.; Huang, J. Evaluation on the detection of Enterocytozoon hepatopenaei in pooled DNA samples of Litopenaeus vannamei based on TaqMan qPCR. Prog. Fish. Sci. 2019, 40, 122–132. [Google Scholar] [CrossRef]
- Wang, W.J.; Lyu, D.; Wang, M.S.; Liu, K.F.; Kong, J.; Shan, X.J.; Jin, X.S. Research in migration route of hatchery released Chinese shrimp (Fenneropenaeus chinensis) in the Bohai Bay using method of SSR marker. Acta Oceanol. Sin. 2020, 39, 76–81. [Google Scholar] [CrossRef]
- Li, P.F.; Zhang, H.; Zhang, X.M.; Gao, T.X.; Han, Z.Q. Study on the genetic variability of the hatchery-released and wild populations of Chinese white shrimp Fenneropenaeus chinensis in the Yellow Sea and Bohai Sea. Aquac. Int. 2017, 25, 2117–2126. [Google Scholar] [CrossRef]
- Spann, K.M.; Cowley, J.A.; Walker, P.J.; Lester, R.J.G. A yellow-head-like virus from Penaeus monodon cultured in Australia. Dis. Aquat. Org. 1997, 31, 169–179. [Google Scholar] [CrossRef]
- Teixeira-Lopes, M.A.; Vieira-Girão, P.R.N.; Freire, J.E.D.C.; Rocha, Í.R.C.B.; Costa, F.H.F.; Rádis-Baptista, G. Natural co-infection with infectious hypodermal and hematopoietic necrosis virus (IHHNV) and infectious myonecrosis virus (IMNV) in Litopenaeus vannamei in Brazil. Aquaculture 2011, 312, 212–216. [Google Scholar] [CrossRef]
- Aranguren, L.F.; Tang, K.F.J.; Lightner, D.V. Protection from yellow head virus (YHV) infection in Penaeus vannamei pre-infected with Taura syndrome virus (TSV). Dis. Aquat. Org. 2012, 98, 185–192. [Google Scholar] [CrossRef] [PubMed]
- WOAH. Disease Card: Infection with Decapod Iridescent Virus 1 (A DIV1) Disease Card. World Organisation for Aquatic Animal Health. Available online: https://www.woah.org/app/uploads/2021/03/a-div1-disease-card.pdf (accessed on 8 January 2023).
- Yang, B.; Song, X.L.; Huang, J.; Shi, C.Y.; Liu, L. Evidence of existence of infectious hypodermal and hematopoietic necrosis virus in penaeid shrimp cultured in China. Vet. Microbiol. 2007, 120, 63–70. [Google Scholar] [CrossRef]
- Dong, X.; Li, F.J.; Xie, G.S.; Yang, B.; Zhang, Q.L.; Huang, J. The analysis of infectious hypodermal and hematopoietic necrosis in 2019. In 2020 Analysis of Major Aquatic Animal Diseases in China; BOF and NFTEC; China Agriculture Press: Beijing, China, 2020; pp. 150–165. ISBN 978-7-109-27064-0. [Google Scholar]
- Browdy, C.L. Recent developments in penaeid broodstock and seed production technologies: Improving the outlook for superior captive stocks. Aquaculture 1998, 164, 3–21. [Google Scholar] [CrossRef]
- Bondad-Reantaso, M.G.; Fejzic, N.; Mackinnon, B.; Huchzermeyer, D.; Seric-Haracic, S.; Mardones, F.O.; Mohan, C.V.; Taylor, N.; Jansen, M.D.; Tavornpanich, S.; et al. A 12-point checklist for surveillance of diseases of aquatic organisms: A novel approach to assist multidisciplinary teams in developing countries. Rev. Aquac. 2021, 13, 1469–1487. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′-3′) | Source |
|---|---|---|
| WSS-1011F | TGGTCCCGTCCTCATCTCAG | [35] |
| WSS-1079R | GCTGCCTTGCCGGAAATTA | |
| WSS-Probe | AGCCATGAAGAATGCCGTCTATCACACA | |
| WSSV-146F1 | ACTACTAACTTCAGCCTATCTAG | [34] |
| WSSV-146R1 | TAATGCGGGTGTAATGTTCTTACGA | |
| WSSV-146F2 | GTAACTGCCCCTTCCATCTCCA | |
| WSSV-146R2 | TACGGCAGCTGCTGCACCTTGT | |
| DIV1-142F | AATCCATGCAAGGTTCCTCAGG | [47] |
| DIV1-142R | CAATCAACATGTCGCGG GAAC | |
| DIV1-Probe | CCATAGTGCTCGCTCGGCTTCGG | |
| DIV1-F1 | GGGCGGGAGATGGTGTTAGAT | [31] |
| DIV1-R1 | TCGTTTCGGTACGAAGATGTA | |
| DIV1-F2 | CGGGAAACGATTCGTATTGGG | |
| DIV1-R2 | TTGCTTGATCGGCATCCTTGA | |
| IHHNV-1608F | TACTCCGGACACCCAACCA | [36] |
| IHHNV-1688R | GGCTCTGGCAGCAAAGGTAA | |
| IHHNV-Probe | ACCAGACATAGAGCTACAATCCTCGCCTATTTG | |
| IHHNV-389F | CGGAACACAACCCGACTTTA | [38] |
| IHHNV-389R | GGCCAAGACCAAAATACGAA | |
| EHP-157F | AGTAAACTATGCCGACAA | [37] |
| EHP-157R | AATTAAGCAGCACAATCC | |
| EHP-Probe | TCCTGGTAGTGTCCTTCCGT | |
| EHP-SWP-1F | TTGCAGAGTGTTGTTAAGGGTTT | [39] |
| EHP- SWP-1R | CACGATGTGTCTTTGCAATTTTC | |
| EHP-SWP-2F | TTGGCGGCACAATTCTCAAACA | |
| EHP-SWP-2R | GCTGTTTGTCTCCAACTGTATTTGA | |
| CMNV-TAQ-F | CGAGCTAATCCAAGCACTTC | [42] |
| CMNV-TAQ-R | ACCTGTTAGGTACGCTACCA | |
| CMNV-TAQ-Probe | CGCTCACGGCTTTGGAT ACCTT | |
| CMNV-7F1 | AAATACGGCGATGACG | [44] |
| CMNV-7R1 | ACGAAGTGCCCACAGAC | |
| CMNV-7F2 | CACAACCGAGTCAAACC | |
| CMNV-7R2 | GCGTAAACAGCGAAGG | |
| IMNV-412F | GGACCTATCATACATAGCGTTGCA | [40] |
| IMNV-545R | AACCCATATCTATTGTCGCTGGAT | |
| IMNV-Probe | CCACCTTTACTTTCAATACTACATCATCCCCGG | |
| IMNV-4587F | CGACGCTGCTAACCATACAA | [43] |
| IMNV-4914R | ACTCGGCTGTTCGATCAAGT | |
| IMNV-4725NF | GGCACATGCTCAGAGACA | |
| IMNV-4863NR | AGCGCTGAGTCCAGTCTTG | |
| TSV-1004F | TTGGGCACCAAACGACATT | [46] |
| TSV-1075R | GGGAGCTTAAACTGGACACACTGT | |
| TSV-Probe | CAGCACTGACGCACAATATTCGAGCATC | |
| TSV-7171F | CGACAGTTGGACATCTAGTG | [45] |
| TSV-7511R | GAGCTTCAGACTGCAACTTC | |
| OWV1-S1-F | CCGACATGGATGCGTTCA | [24] |
| OWV1-S1-R | CAAGGCTGCAACAATGACCTT | |
| OWV1-S1-Probe | CGCAGACATCCAGTTCCAGGGCTTT | |
| OWV1-F1 | CAAAGACGGGTGTTGTAGTGT | In this study |
| OWV1-R1 | AGCTTGCCTCACCGTTCTC | |
| OWV1-F2 | ATTCGTCTCACTCCTGGCTG | |
| OWV1-R2 | CGCCTTCCATGTACTCGCTA | |
| OWV1-F3 | CCACCACCTCAAGTCCACA | |
| OWV1-R3 | TCATCAGAGTATGGGGACAGGT | |
| OWV1-F4 | TGAAGAGGAGGGAGGTGCAT | |
| OWV1-R4 | ACAAAGACCGGGTGTGTTCTAA | |
| GY1 | GACATCACTCCAGACAACATCTG | [48] |
| GY2 | CATCTGTCCAGAAGGCGTCTATGA | |
| GY4 | GTGAAGTCCATGTGTGTGAGACG | |
| GY5 | GAGCTGGAATTCAGTGAGAGAACA | |
| Y3 | ACGCTCTGTGACAAGCATGAAGTT | |
| G6 | GTAGTAGAGACGAGTGACACCTAT | |
| YH30-F1m | TACCAYTCAAACATCATYAAYAAYCAYCA | [19] |
| YH30-R1m | GAGATGATYTGRTKCTTRAAYTTCTGRAA | |
| YH31-F2m | CTCARATCCATGCMATYTGGGARTCHTC | |
| YH31-R2m | AGTTTGGCRCGRATRTTRGTRAGRAT |
| Viruses | Species | Detection Method | Number of Samples | ||
|---|---|---|---|---|---|
| 20201124001-009 | 20211126001-008 | 20220309001-020 | |||
| DIV1 | P. chinensis | Nested -PCR | P | N | N |
| qPCR (cycles) | 31.3 ± 4.3 | N | N | ||
| IHHNV | P. chinensis | PCR | P | N | P |
| qPCR (cycles) | 36.7 ± 1.4 | N | 28.5 ± 0.2 | ||
| CMNV | P. chinensis | Nested RT-PCR | N | N | N |
| RT-qPCR (cycles) | N | N | N | ||
| EHP | P. chinensis | Nested -PCR | N | N | N |
| qPCR (cycles) | N | N | N | ||
| WSSV | P. chinensis | Nested -PCR | N | N | N |
| qPCR (cycles) | N | N | N | ||
| IMNV | P. chinensis | Nested RT-PCR | N | N | N |
| RT-qPCR (cycles) | N | N | N | ||
| TSV | P. chinensis | RT-PCR | N | N | N |
| RT-qPCR (cycles) | N | N | N | ||
| Number of Samples | Species | Viruses and Detection Method | |
|---|---|---|---|
| OWV1 RT-qPCR (Cycles) | YHVs Nested RT-PCR | ||
| 20201124001 | P. chinensis | 13.8 ± 0.1 | P |
| 20201124002 | P. chinensis | 25.2 ± 1.0 | P |
| 20201124003 | P. chinensis | 11.4 ± 0.1 | P |
| 20201124004 | P. chinensis | 12.3 ± 0.2 | P |
| 20201124005 | P. chinensis | 11.9 ± 0.3 | P |
| 20201124006 | P. chinensis | 13.2 ± 0.2 | P |
| 20201124007 | P. chinensis | 29.1 ± 0.1 | P |
| 20201124008 | P. chinensis | 26.6 ± 0.1 | P |
| 20201124009 | P. chinensis | 29.6 ± 0.1 | P |
| 20211126001 | P. chinensis | 26.2 ± 0.1 | P |
| 20211126002 | P. chinensis | 10.2 ± 0.2 | P |
| 20211126003 | P. chinensis | 19.6 ± 0.1 | P |
| 20211126004 | P. chinensis | 26.6 ± 0.1 | P |
| 20211126005 | P. chinensis | 11.3 ± 0.6 | P |
| 20211126006 | P. chinensis | 12.6 ± 0.4 | P |
| 20211126007 | P. chinensis | 9.2 ± 0.1 | P |
| 20211126008 | P. chinensis | 14.5 ± 0.2 | P |
| 20220309001 | P. chinensis | 20.2 ± 0.3 | P |
| 20220309002 | P. chinensis | 25.0 ± 0.8 | P |
| 20220309003 | P. chinensis | 26.2 ± 0.4 | P |
| 20220309004 | P. chinensis | 14.1 ± 0.5 | P |
| 20220309005 | P. chinensis | 19.1 ± 0.9 | P |
| 20220309006 | P. chinensis | 19.0 ± 1.1 | P |
| 20220309007 | P. chinensis | 32.5 ± 3.2 | P |
| 20220309008 | P. chinensis | 13.7 ± 1.5 | P |
| 20220309009 | P. chinensis | 28.6 ± 0.5 | P |
| 20220309010 | P. chinensis | 30.4 ± 0.0 | P |
| 20220309011 | P. chinensis | 27.3 ± 0.7 | P |
| 20220309012 | P. chinensis | 29.9 ± 1.1 | P |
| 20220309013 | P. chinensis | 24.2 ± 0.4 | P |
| 20220309014 | P. chinensis | 26.9 ± 0.5 | P |
| 20220309015 | P. chinensis | 31.2 ± 0.8 | P |
| 20220309016 | P. chinensis | 35.0 ± 1.7 | P |
| 20220309017 | P. chinensis | 10.0 ± 6.5 | P |
| 20220309018 | P. chinensis | 32.4 ± 3.5 | P |
| 20220309019 | P. chinensis | 29.1 ± 0.9 | P |
| 20220309020 | P. chinensis | 25.4 ± 0.1 | P |
| Source | Object of Comparison | Max Score | Query Cover | Percent Identity |
|---|---|---|---|---|
| 20201124002 | YHV-8 20120706 (NC_048215.1) | 1203 | 97% | 99.10% |
| 20201124003 | YHV-8 20120706 (NC_048215.1) | 1186 | 97% | 98.65% |
| 20201124004 | YHV-8 20120706 (NC_048215.1) | 1197 | 96% | 99.10% |
| 20211124005 | YHV-8 20120706 (NC_048215.1) | 1203 | 99% | 98.96% |
| 20201124006 | YHV-8 20120706 (NC_048215.1) | 1186 | 97% | 98.65% |
| 20201124008 | YHV-8 20120706 (NC_048215.1) | 1197 | 97% | 98.95% |
| 20201124009 | YHV-8 20120706 (NC_048215.1) | 1192 | 96% | 98.80% |
| 20211126001 | YHV-8 20120706 (NC_048215.1) | 1151 | 100% | 98.32% |
| 20211126002 | YHV-8 20120706 (NC_048215.1) | 1168 | 100% | 98.78% |
| 20211126003 | YHV-8 20120706 (NC_048215.1) | 1151 | 100% | 98.32% |
| 20211126004 | YHV-8 20120706 (NC_048215.1) | 1162 | 100% | 98.63% |
| 20211126005 | YHV-8 20120706 (NC_048215.1) | 1168 | 100% | 98.78% |
| 20211126006 | YHV-8 20120706 (NC_048215.1) | 1184 | 100% | 99.24% |
| 20211126007 | YHV-8 20120706 (NC_048215.1) | 1175 | 99% | 99.08% |
| 20211126008 | YHV-8 20120706 (NC_048215.1) | 1170 | 99% | 98.93% |
| 20220309001 | YHV-8 20120706 (NC_048215.1) | 1203 | 97% | 99.10% |
| 20220309002 | YHV-8 20120706 (NC_048215.1) | 1197 | 97% | 98.95% |
| Source | Object of Comparison | Max Score | Query Cover | Percent Identity |
|---|---|---|---|---|
| 20201124004 | OWV1 (QHW05228.1) | 1449 | 100% | 99.42% |
| 20201124005 | OWV1 (QHW05228.1) | 1451 | 99% | 99.57% |
| 20201124006 | OWV1 (QHW05228.1) | 1446 | 100% | 99.14% |
| 20211126001 | OWV1 (QHW05228.1) | 1449 | 100% | 99.42% |
| 20211126003 | OWV1 (QHW05228.1) | 1447 | 100% | 99.14% |
| 20211126008 | OWV1 (QHW05228.1) | 1449 | 100% | 99.42% |
| 20220309001 | OWV1 (QHW05228.1) | 1451 | 100% | 99.57% |
| 20220309002 | OWV1 (QHW05228.1) | 1449 | 100% | 99.42% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, J.; Meng, F.; Wang, G.; Chen, Y.; Zhang, F.; Li, C.; Dong, X.; Huang, J. Coinfection with Yellow Head Virus Genotype 8 (YHV-8) and Oriental Wenrivirus 1 (OWV1) in Wild Penaeus chinensis from the Yellow Sea. Viruses 2023, 15, 361. https://doi.org/10.3390/v15020361
Qin J, Meng F, Wang G, Chen Y, Zhang F, Li C, Dong X, Huang J. Coinfection with Yellow Head Virus Genotype 8 (YHV-8) and Oriental Wenrivirus 1 (OWV1) in Wild Penaeus chinensis from the Yellow Sea. Viruses. 2023; 15(2):361. https://doi.org/10.3390/v15020361
Chicago/Turabian StyleQin, Jiahao, Fanzeng Meng, Guohao Wang, Yujin Chen, Fan Zhang, Chen Li, Xuan Dong, and Jie Huang. 2023. "Coinfection with Yellow Head Virus Genotype 8 (YHV-8) and Oriental Wenrivirus 1 (OWV1) in Wild Penaeus chinensis from the Yellow Sea" Viruses 15, no. 2: 361. https://doi.org/10.3390/v15020361
APA StyleQin, J., Meng, F., Wang, G., Chen, Y., Zhang, F., Li, C., Dong, X., & Huang, J. (2023). Coinfection with Yellow Head Virus Genotype 8 (YHV-8) and Oriental Wenrivirus 1 (OWV1) in Wild Penaeus chinensis from the Yellow Sea. Viruses, 15(2), 361. https://doi.org/10.3390/v15020361