
Improved Neutralisation of the SARS-CoV-2 Omicron Variant following a Booster Dose of Pfizer-BioNTech (BNT162b2) COVID-19 Vaccine

 , ,
, , 
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. SARS-CoV-2 Culture
2.2. SARS-CoV-2 Viral Load Quantitation by RT-qPCR
2.3. SARS-CoV-2 Sequencing
2.4. Bioinformatic Analysis
2.5. Post Pfizer-BioNTech (BNT162b2) Vaccine Sera
2.6. Determination of 50% Tissue Culture Infective Dose (TCID50)
2.7. Micro-neutralisation Assay
2.8. Statistical Analysis
3. Results
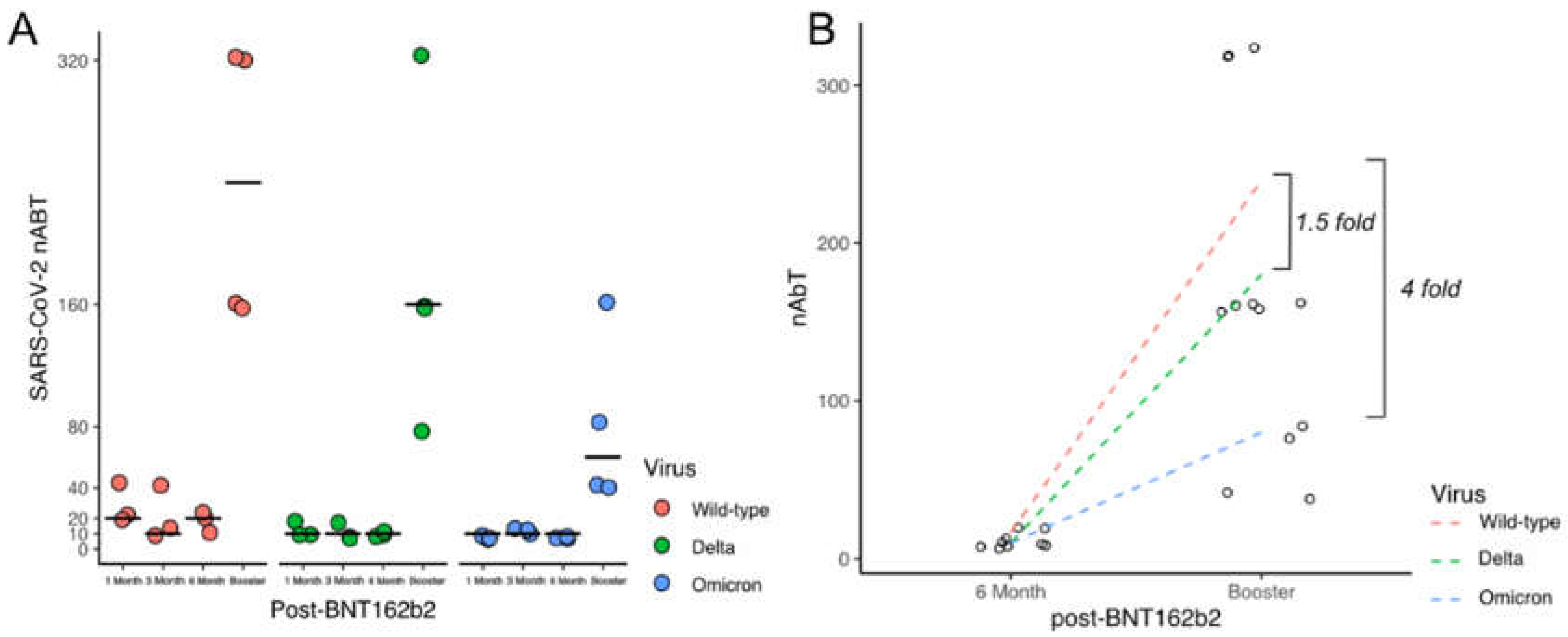
3.1. Levels of Neutralising Antibodies against Different SARS-CoV-2 Lineages
3.2. Different in Vitro Infection Kinetics between SARS-CoV-2 Lineages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Network for Genomic Surveillance in South Africa (NGS-SA) SARS-CoV-2 Sequencing Update 1 December 2021. Network for Genomic Surveillance in South Africa (NGS-SA). 2021. Available online: https://www.nicd.ac.za/wp-content/uploads/2021/12/update-of-sa-sequencing-data-from-gisaid-1-dec-final.pdf (accessed on 6 December 2021).
- World Health Organization Statement—Classification of Omicron (B.1.1.529): SARS-CoV-2 Variant of Concern 26 November 2021. Available online: https://www.who.int/news/item/26-11-2021-classification-of-omicron-(b.1.1.529)-sars-cov-2-variant-of-concern (accessed on 6 December 2021).
- The World Health Organization—Tracking SARS-CoV-2 Variants. Available online: https://Www.Who.Int/Activities/Tracking-SARS-CoV-2-Variants (accessed on 4 August 2022).
- The World Health Organization—COVID-19 Vaccines with WHO Emergency Use Listing. Available online: https://Extranet.Who.Int/Pqweb/Vaccines/Vaccinescovid-19-Vaccine-Eul-Issued (accessed on 4 August 2022).
- Liu, Y.; Liu, J.; Xia, H.; Zhang, X.; Fontes-Garfias, C.R.; Swanson, K.A.; Cai, H.; Sarkar, R.; Chen, W.; Cutler, M.; et al. Neutralizing Activity of BNT162b2-Elicited Serum. N. Engl. J. Med. 2021, 384, 1466–1468. [Google Scholar] [CrossRef] [PubMed]
- Lopez Bernal, J.; Andrews, N.; Gower, C.; Gallagher, E.; Simmons, R.; Thelwall, S.; Stowe, J.; Tessier, E.; Groves, N.; Dabrera, G.; et al. Effectiveness of Covid-19 Vaccines against the B.1.617.2 (Delta) Variant. N. Engl. J. Med. 2021, 385, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Chemaitelly, H.; Yassine, H.M.; Benslimane, F.M.; al Khatib, H.A.; Tang, P.; Hasan, M.R.; Malek, J.A.; Coyle, P.; Ayoub, H.H.; al Kanaani, Z.; et al. MRNA-1273 COVID-19 Vaccine Effectiveness against the B.1.1.7 and B.1.351 Variants and Severe COVID-19 Disease in Qatar. Nat. Med. 2021, 27, 1614–1621. [Google Scholar] [CrossRef] [PubMed]
- Graham, M.S.; Sudre, C.H.; May, A.; Antonelli, M.; Murray, B.; Varsavsky, T.; Kläser, K.; Canas, L.S.; Molteni, E.; Modat, M.; et al. Changes in Symptomatology, Reinfection, and Transmissibility Associated with the SARS-CoV-2 Variant B.1.1.7: An Ecological Study. Lancet Public Health 2021, 6, e335–e345. [Google Scholar] [CrossRef]
- Jangra, S.; Ye, C.; Rathnasinghe, R.; Stadlbauer, D.; Alshammary, H.; Amoako, A.A.; Awawda, M.H.; Beach, K.F.; Bermúdez-González, M.C.; Chernet, R.L.; et al. SARS-CoV-2 Spike E484K Mutation Reduces Antibody Neutralisation. Lancet Microbe 2021, 2, e283–e284. [Google Scholar] [CrossRef]
- Xie, X.; Liu, Y.; Liu, J.; Zhang, X.; Zou, J.; Fontes-Garfias, C.R.; Xia, H.; Swanson, K.A.; Cutler, M.; Cooper, D.; et al. Neutralization of SARS-CoV-2 Spike 69/70 Deletion, E484K and N501Y Variants by BNT162b2 Vaccine-Elicited Sera. Nat. Med. 2021, 27, 620–621. [Google Scholar] [CrossRef]
- Willett, B.J.; Grove, J.; MacLean, O.A.; Wilkie, C.; de Lorenzo, G.; Furnon, W.; Cantoni, D.; Scott, S.; Logan, N.; Ashraf, S.; et al. SARS-CoV-2 Omicron Is an Immune Escape Variant with an Altered Cell Entry Pathway. Nat. Microbiol. 2022, 7, 1161–1179. [Google Scholar] [CrossRef]
- VanBlargan, L.A.; Errico, J.M.; Halfmann, P.J.; Zost, S.J.; Crowe, J.E.; Purcell, L.A.; Kawaoka, Y.; Corti, D.; Fremont, D.H.; Diamond, M.S. An Infectious SARS-CoV-2 B.1.1.529 Omicron Virus Escapes Neutralization by Therapeutic Monoclonal Antibodies. Nat. Med. 2022, 28, 490–495. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, S.; Wu, B.; Yang, Q.; Chen, A.; Li, Y.; Zhang, Y.; Pan, T.; Zhang, H.; He, X. SARS-CoV-2 Omicron Strain Exhibits Potent Capabilities for Immune Evasion and Viral Entrance. Signal Transduct. Target. Ther. 2021, 6, 430. [Google Scholar] [CrossRef]
- Chaguza, C.; Coppi, A.; Earnest, R.; Ferguson, D.; Kerantzas, N.; Warner, F.; Young, H.P.; Breban, M.I.; Billig, K.; Koch, R.T.; et al. Rapid Emergence of SARS-CoV-2 Omicron Variant Is Associated with an Infection Advantage over Delta in Vaccinated Persons. Med 2022, 3, 325–334.e4. [Google Scholar] [CrossRef]
- Rahman, H.; Carter, I.; Basile, K.; Donovan, L.; Kumar, S.; Tran, T.; Ko, D.; Alderson, S.; Sivaruban, T.; Eden, J.-S.; et al. Interpret with Caution: An Evaluation of the Commercial AusDiagnostics versus in-House Developed Assays for the Detection of SARS-CoV-2 Virus. J. Clin. Virol. 2020, 127, 104374. [Google Scholar] [CrossRef] [PubMed]
- Basile, K.; McPhie, K.; Carter, I.; Alderson, S.; Rahman, H.; Donovan, L.; Kumar, S.; Tran, T.; Ko, D.; Sivaruban, T.; et al. Cell-Based Culture of SARS-CoV-2 Informs Infectivity and Safe de-Isolation Assessments during COVID-19. Clin. Infect. Dis. 2020, 73, e2952–e2959. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.A.; Xie, X.; Bailey, A.L.; Kalveram, B.; Lokugamage, K.G.; Muruato, A.; Zou, J.; Zhang, X.; Juelich, T.; Smith, J.K.; et al. Loss of Furin Cleavage Site Attenuates SARS-CoV-2 Pathogenesis. Nature 2021, 591, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Klimstra, W.B.; Tilston-Lunel, N.L.; Nambulli, S.; Boslett, J.; McMillen, C.M.; Gilliland, T.; Dunn, M.D.; Sun, C.; Wheeler, S.E.; Wells, A.; et al. SARS-CoV-2 Growth, Furin-Cleavage-Site Adaptation and Neutralization Using Serum from Acutely Infected Hospitalized COVID-19 Patients. J. Gen. Virol. 2020, 101, 1156–1169. [Google Scholar] [CrossRef]
- Peacock, T.P.; Goldhill, D.H.; Zhou, J.; Baillon, L.; Frise, R.; Swann, O.C.; Kugathasan, R.; Penn, R.; Brown, J.C.; Sanchez-David, R.Y.; et al. The Furin Cleavage Site in the SARS-CoV-2 Spike Protein Is Required for Transmission in Ferrets. Nat. Microbiol. 2021, 6, 899–909. [Google Scholar] [CrossRef]
- Lam, C.; Gray, K.; Gall, M.; Sadsad, R.; Arnott, A.; Johnson-Mackinnon, J.; Fong, W.; Basile, K.; Kok, J.; Dwyer, D.E.; et al. SARS-CoV-2 Genome Sequencing Methods Differ in Their Abilities To Detect Variants from Low-Viral-Load Samples. J. Clin. Microbiol. 2021, 59, e0104621. [Google Scholar] [CrossRef]
- Freed, N.E.; Vlková, M.; Faisal, M.B.; Silander, O.K. Rapid and Inexpensive Whole-Genome Sequencing of SARS-CoV-2 Using 1200 Bp Tiled Amplicons and Oxford Nanopore Rapid Barcoding. Biol. Methods Protoc. 2021, 5, bpaa014. [Google Scholar] [CrossRef]
- Rockett, R.J.; Arnott, A.; Lam, C.; Sadsad, R.; Timms, V.; Gray, K.-A.; Eden, J.-S.; Chang, S.; Gall, M.; Draper, J.; et al. Revealing COVID-19 Transmission in Australia by SARS-CoV-2 Genome Sequencing and Agent-Based Modeling. Nat. Med. 2020, 26, 1398–1404. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and Accurate Short Read Alignment with Burrows-Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Grubaugh, N.D.; Gangavarapu, K.; Quick, J.; Matteson, N.L.; De Jesus, J.G.; Main, B.J.; Tan, A.L.; Paul, L.M.; Brackney, D.E.; Grewal, S.; et al. An Amplicon-Based Sequencing Framework for Accurately Measuring Intrahost Virus Diversity Using PrimalSeq and Ivar. Genome Biol. 2019, 20, 8. [Google Scholar] [CrossRef] [PubMed]
- Turakhia, Y.; de Maio, N.; Thornlow, B.; Gozashti, L.; Lanfear, R.; Walker, C.R.; Hinrichs, A.S.; Fernandes, J.D.; Borges, R.; Slodkowicz, G.; et al. Stability of SARS-CoV-2 Phylogenies. PLoS Genet. 2020, 16, e1009175. [Google Scholar] [CrossRef]
- O’Toole, Á.; Scher, E.; Underwood, A.; Jackson, B.; Hill, V.; McCrone, J.T.; Colquhoun, R.; Ruis, C.; Abu-Dahab, K.; Taylor, B.; et al. Assignment of Epidemiological Lineages in an Emerging Pandemic Using the Pangolin Tool. Virus Evol. 2021, 7, veab064. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Holmes, E.C.; O’Toole, Á.; Hill, V.; McCrone, J.T.; Ruis, C.; du Plessis, L.; Pybus, O.G. A Dynamic Nomenclature Proposal for SARS-CoV-2 Lineages to Assist Genomic Epidemiology. Nat. Microbiol. 2020, 5, 1403–1407. [Google Scholar] [CrossRef] [PubMed]
- Hueston, L.; Kok, J.; Guibone, A.; McDonald, D.; Hone, G.; Goodwin, J.; Carter, I.; Basile, K.; Sandaradura, I.; Maddocks, S.; et al. The Antibody Response to SARS-CoV-2 Infection. Open Forum Infect. Dis. 2020, 7, ofaa387. [Google Scholar] [CrossRef]
- SARS-CoV-2 Inactivation Testing: Interim Report; 2020. High Containment Microbiology, NIS Laboratories, National Infection Service, Public Health England. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/898679/HCM-CoV2-023-v1_Triton_X-100_Serum.pdf (accessed on 1 August 2021).
- Cele, S.; Jackson, L.; Khoury, D.S.; Khan, K.; Moyo-Gwete, T.; Tegally, H.; San, J.E.; Cromer, D.; Scheepers, C.; Amoako, D.G.; et al. Omicron Extensively but Incompletely Escapes Pfizer BNT162b2 Neutralization. Nature 2022, 602, 654–656. [Google Scholar] [CrossRef] [PubMed]
- Arbel, R.; Hammerman, A.; Sergienko, R.; Friger, M.; Peretz, A.; Netzer, D.; Yaron, S. BNT162b2 Vaccine Booster and Mortality Due to Covid-19. N. Engl. J. Med. 2021, 385, 2413–2420. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basile, K.; Rockett, R.J.; McPhie, K.; Fennell, M.; Johnson-Mackinnon, J.; Agius, J.E.; Fong, W.; Rahman, H.; Ko, D.; Donavan, L.; et al. Improved Neutralisation of the SARS-CoV-2 Omicron Variant following a Booster Dose of Pfizer-BioNTech (BNT162b2) COVID-19 Vaccine. Viruses 2022, 14, 2023. https://doi.org/10.3390/v14092023
Basile K, Rockett RJ, McPhie K, Fennell M, Johnson-Mackinnon J, Agius JE, Fong W, Rahman H, Ko D, Donavan L, et al. Improved Neutralisation of the SARS-CoV-2 Omicron Variant following a Booster Dose of Pfizer-BioNTech (BNT162b2) COVID-19 Vaccine. Viruses. 2022; 14(9):2023. https://doi.org/10.3390/v14092023
Chicago/Turabian StyleBasile, Kerri, Rebecca J. Rockett, Kenneth McPhie, Michael Fennell, Jessica Johnson-Mackinnon, Jessica E. Agius, Winkie Fong, Hossinur Rahman, Danny Ko, Linda Donavan, and et al. 2022. "Improved Neutralisation of the SARS-CoV-2 Omicron Variant following a Booster Dose of Pfizer-BioNTech (BNT162b2) COVID-19 Vaccine" Viruses 14, no. 9: 2023. https://doi.org/10.3390/v14092023
APA StyleBasile, K., Rockett, R. J., McPhie, K., Fennell, M., Johnson-Mackinnon, J., Agius, J. E., Fong, W., Rahman, H., Ko, D., Donavan, L., Hueston, L., Lam, C., Arnott, A., Chen, S. C.-A., Maddocks, S., O’Sullivan, M. V., Dwyer, D. E., Sintchenko, V., & Kok, J. (2022). Improved Neutralisation of the SARS-CoV-2 Omicron Variant following a Booster Dose of Pfizer-BioNTech (BNT162b2) COVID-19 Vaccine. Viruses, 14(9), 2023. https://doi.org/10.3390/v14092023