
Infection with IBV DMV/1639 at a Young Age Leads to Increased Incidence of Cystic Oviduct Formation Associated with False Layer Syndrome


Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses
2.2. Experimental Design
2.3. RNA Extraction and qRT-PCR
2.4. Histopathology
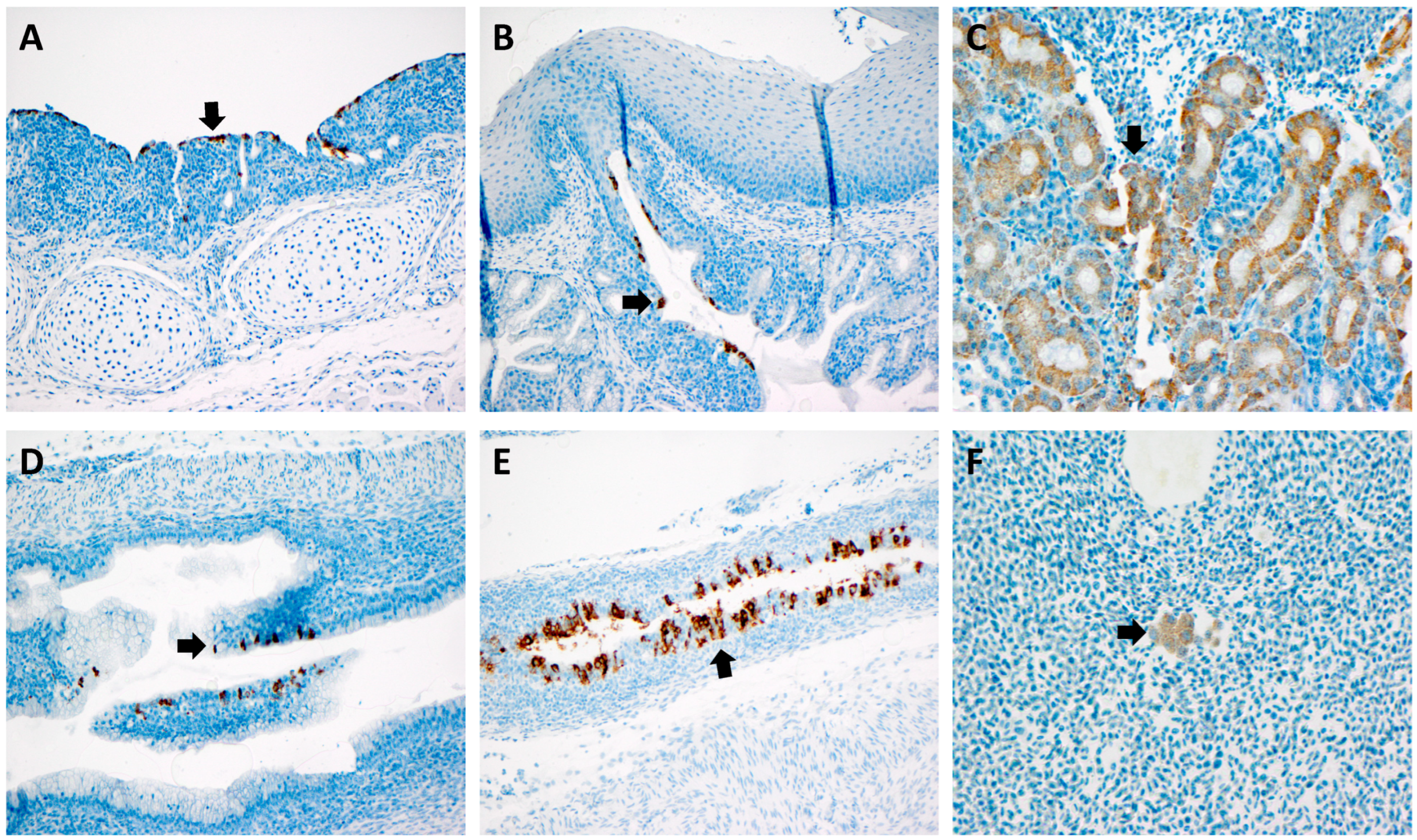
2.5. Riboprobe RNAScope Analysis
3. Results
3.1. qRT-PCR
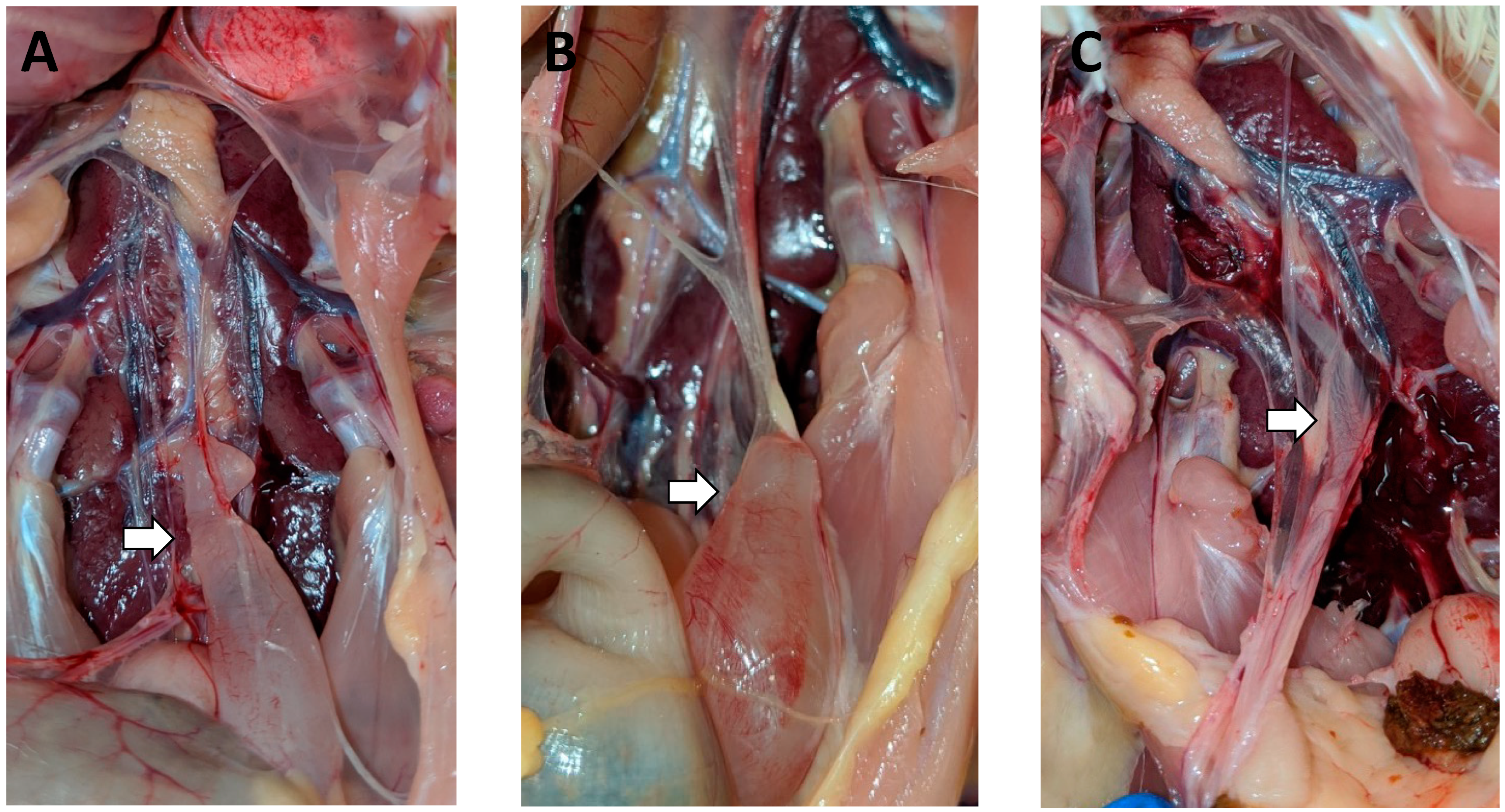
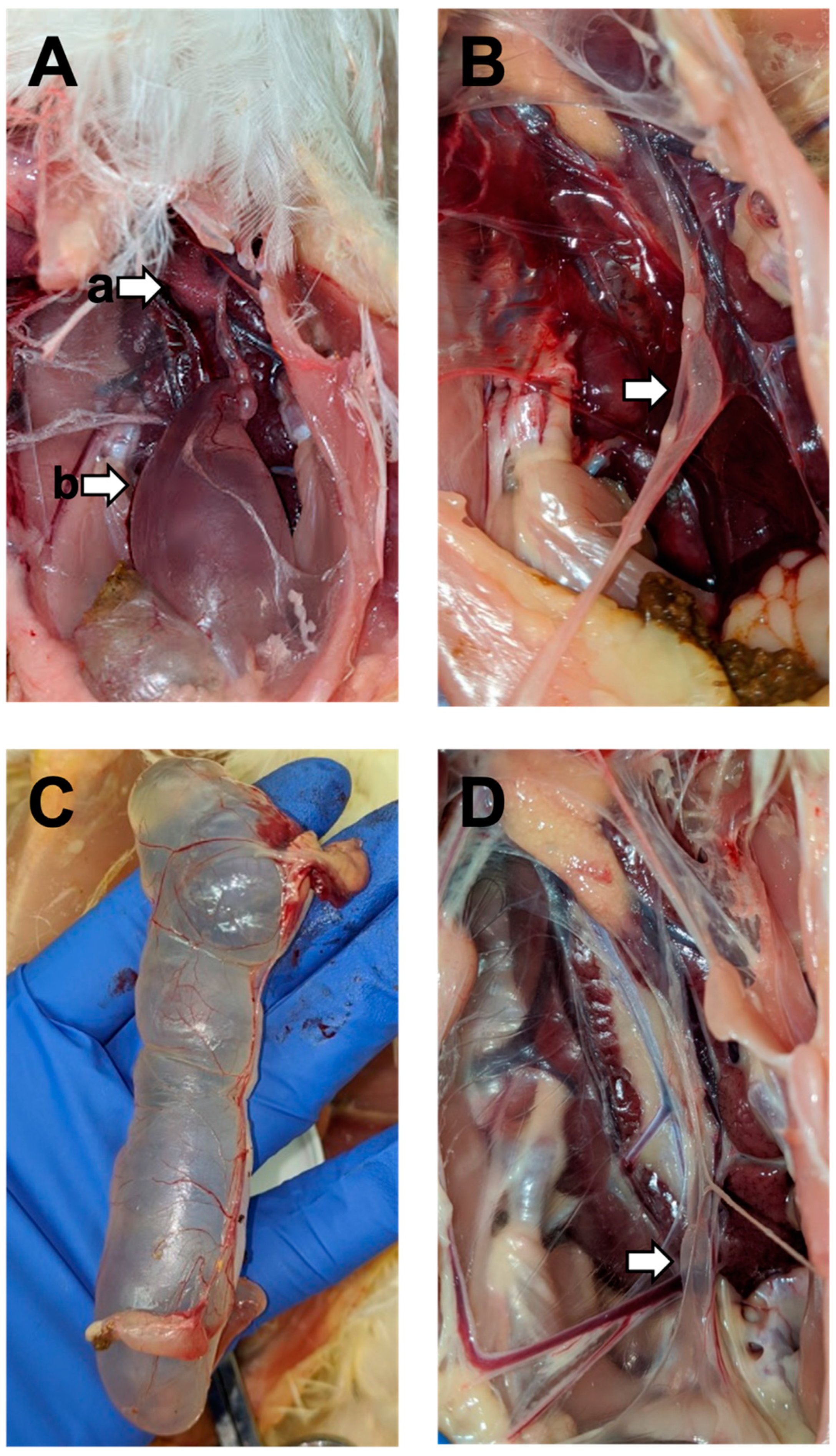
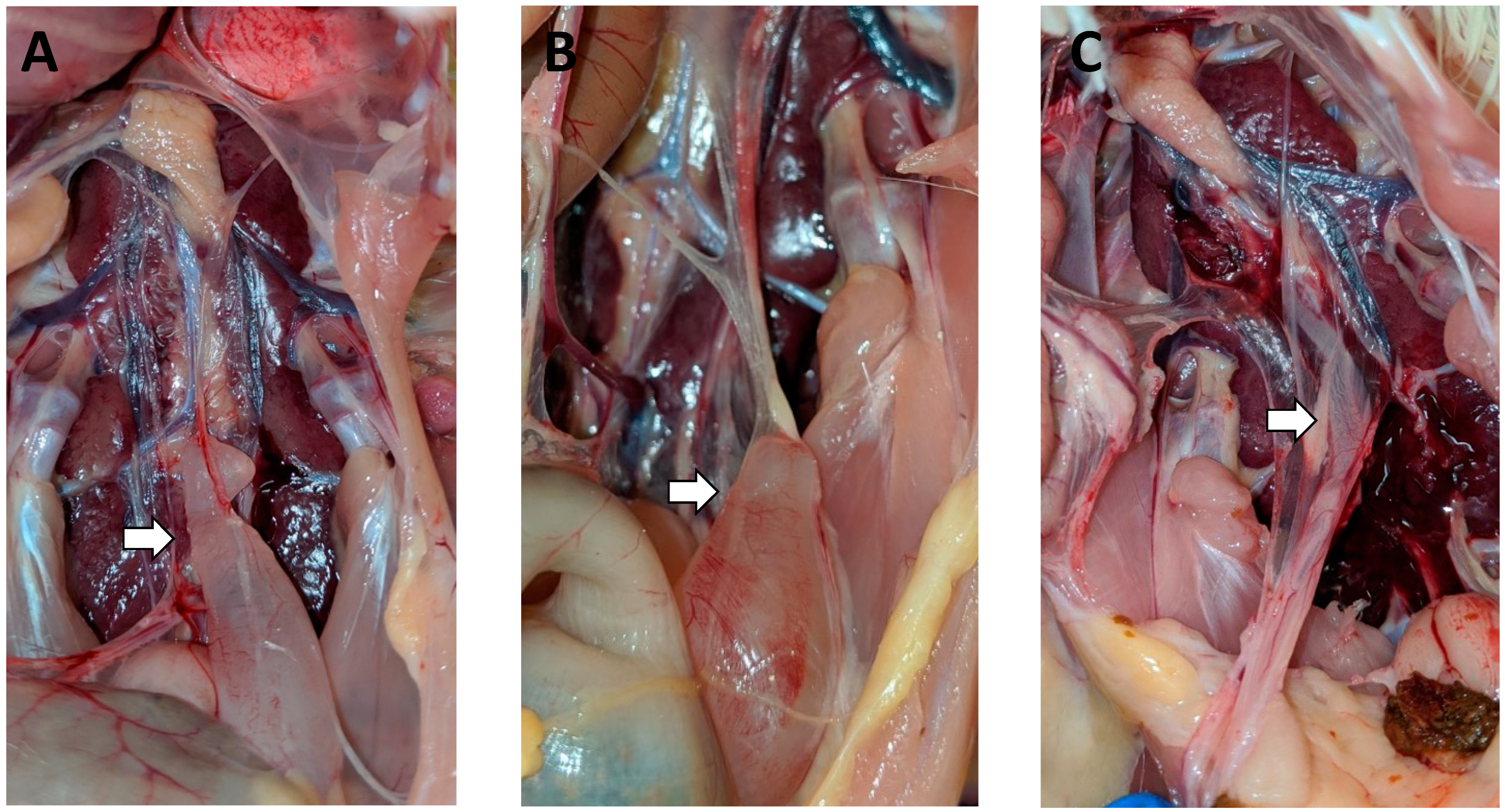
3.2. Gross Lesions
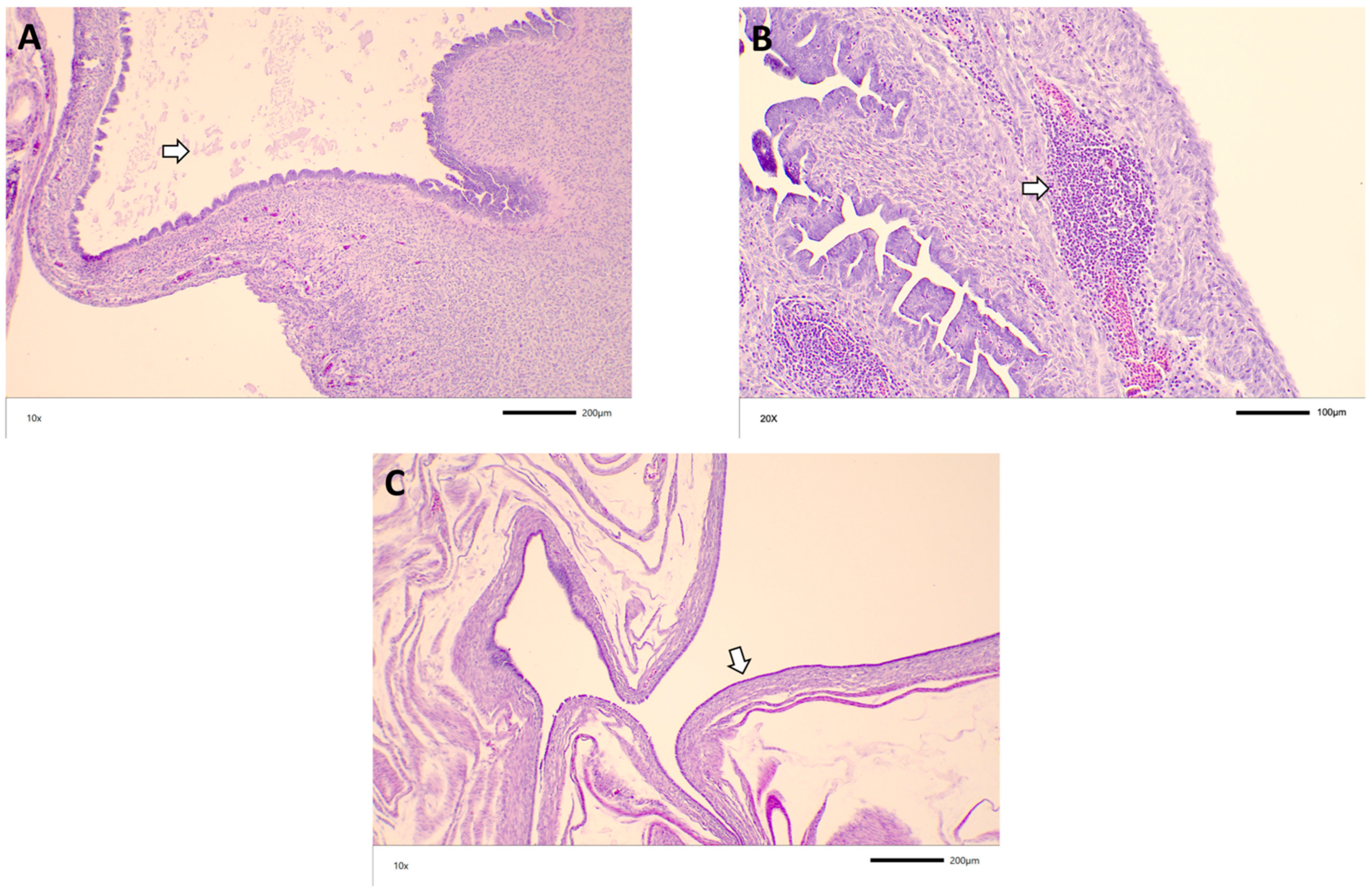
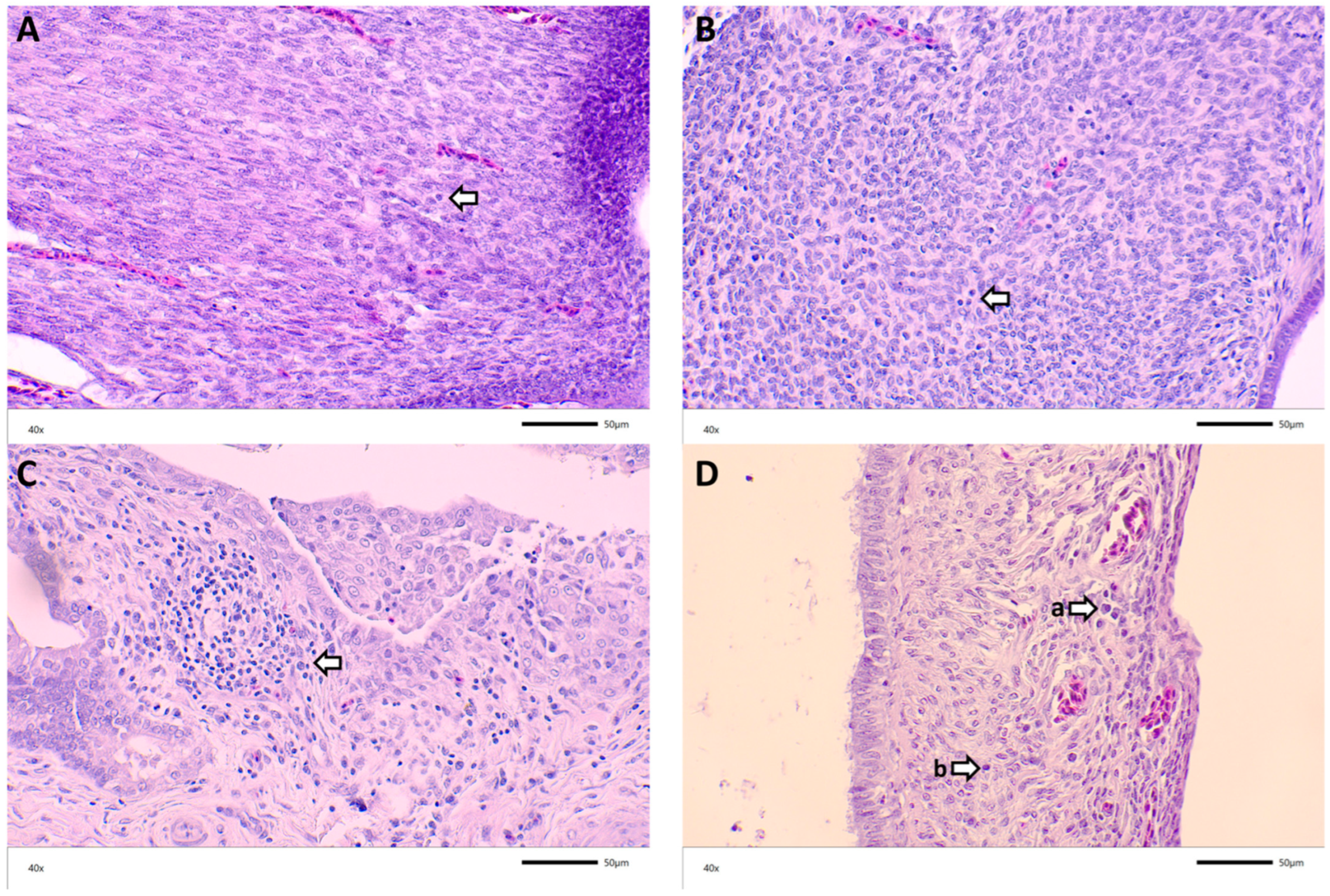
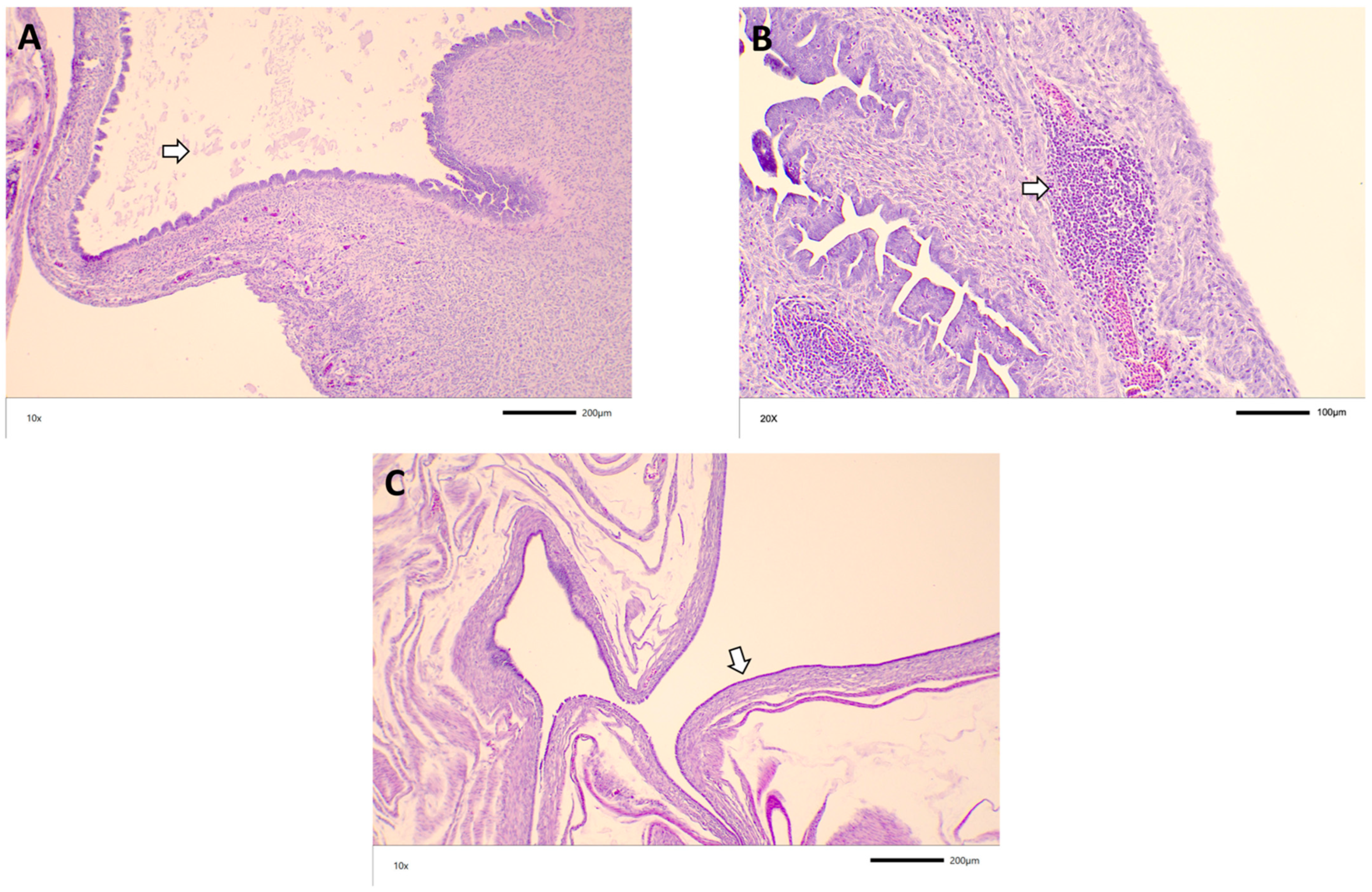
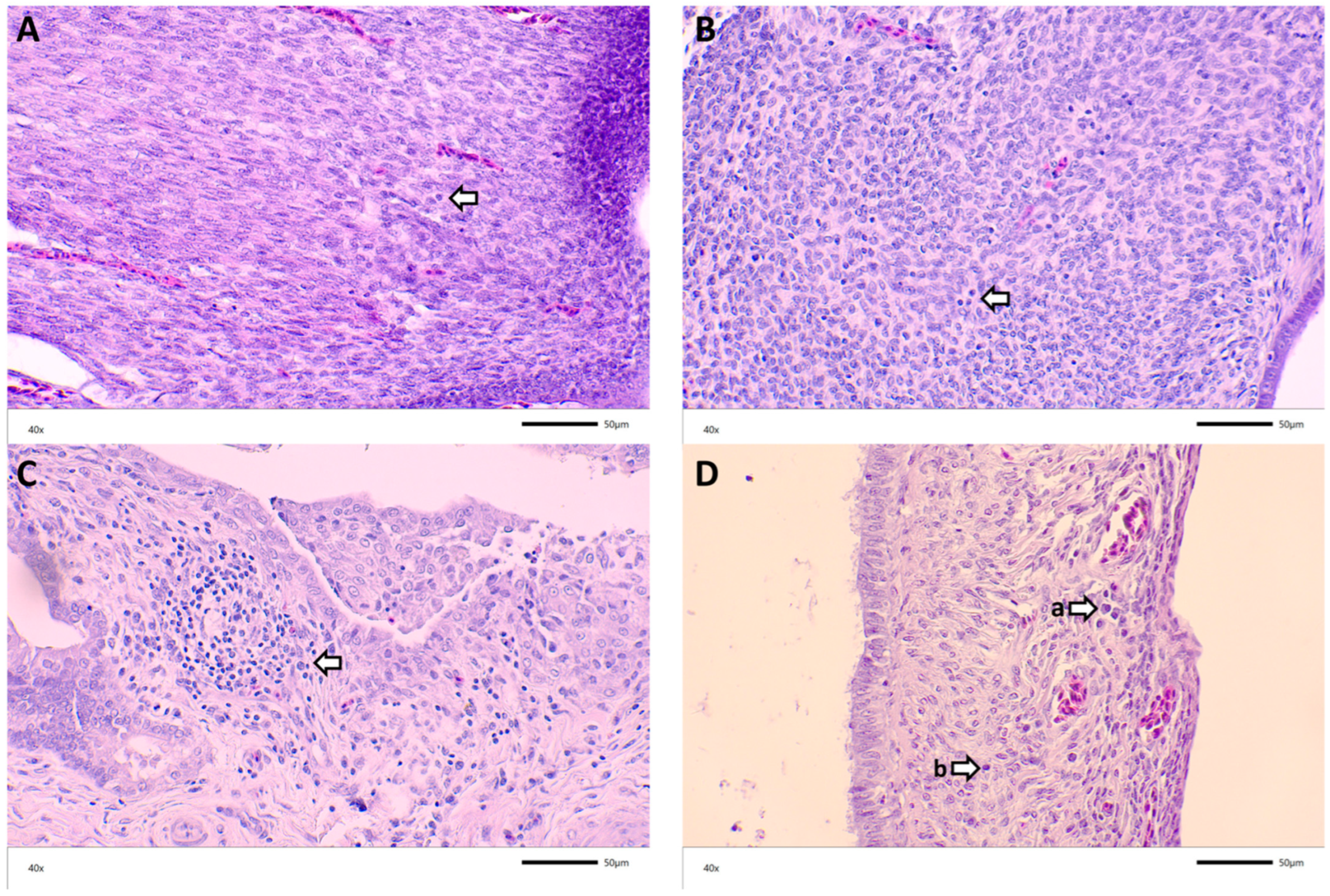
3.3. Histopathology
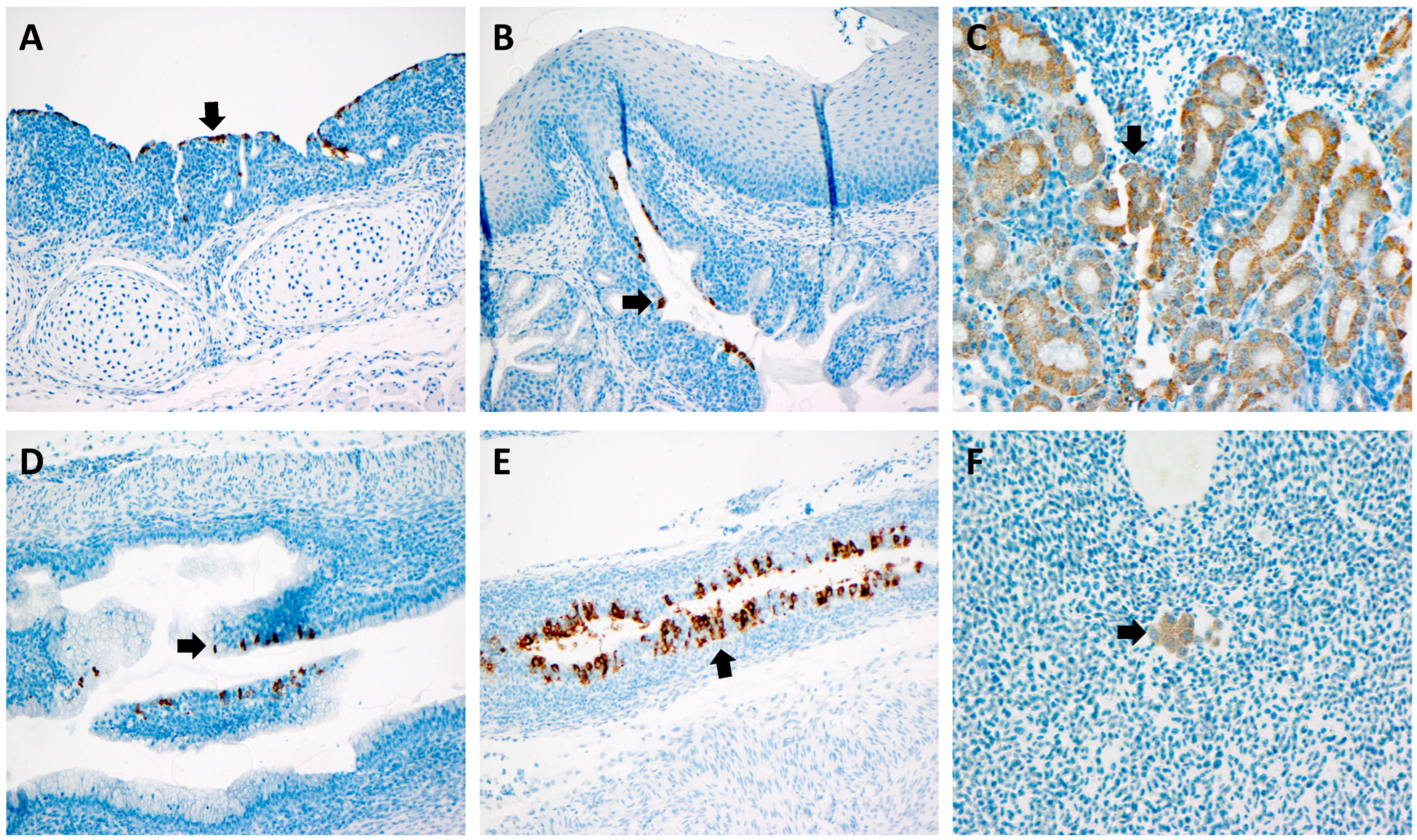
3.4. RNAScope
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Awad, F.; Chhabra, R.; Baylis, M.; Ganapathy, K. An overview of infectious bronchitis virus in chickens. World’s Poult. Sci. J. 2014, 70, 375–384. [Google Scholar] [CrossRef]
- Broadfoot, D.I.; Pomeroy, B.S.; Smith, W.M. Effects of Infectious Bronchitis in baby chicks. Poult. Sci. 1956, 35, 757–762. [Google Scholar] [CrossRef]
- Sevoian, M.; Levine, P.P. Effects of Infectious Bronchitis on the reproductive tracts, egg production, and egg quality of laying chickens. Avian Dis. 1957, 1, 136–164. [Google Scholar] [CrossRef]
- Muneer, M.A.; Halvorson, D.A.; Sivanandan, V.; Newman, J.A.; Coon, C.N. Effects of Infectious Bronchitis Virus (Arkansas Strain) on Laying Chickens. Avian Dis. 1986, 30, 644–647. [Google Scholar] [CrossRef]
- Ignjatovic, J.; Sapats, S. Avian Infectious Bronchitis Virus. Rev. Sci. Tech. 2000, 19, 493–508. [Google Scholar] [CrossRef]
- De Wit, J.J.S.; Cook, J.K.A.; van der Heijden, H.M.J.F. Infectious bronchitis virus variants: A review of the history, current situation and control measures. Avian Pathol. 1972, 40, 223–235. [Google Scholar] [CrossRef]
- Landman, W.J.M. High incidence of false layers in (re)production hens supposedly attributed to a uvenile infectious bronchitis virus infection. Proc. West. Poult. Dis. Conf. 2005, 54, 105–116. [Google Scholar]
- Crinion, R.A.P.; Ball, R.A.; Hofstad, M.S. Abnormalities in laying chickens following exposure to infectious bronchitis virus at one day old. Avian Dis. 1971, 15, 42–48. [Google Scholar] [CrossRef]
- Duff, R.H.; Macdonald, J.W.; McMartin, D.A.; Ross, J.G. Infection of day-old chicks with Infectious Bronchitis (IB) virus and subsequent anatomical abnormalities. Vet. Rec. 1971, 88, 315. [Google Scholar] [CrossRef]
- Zhong, Q.; Hu, Z.; Jin, J.; Zhao, Y.; Zhao, J.; Zhang, G. Pathogenicity of virulent infectious bronchitis virus isolate YN on hen ovary and oviduct. Vet. Microbiol. 2016, 193, 100–105. [Google Scholar] [CrossRef]
- Benyeda, Z.; Szeredi, L.; Mato, T.; Suveges, T.; Balka, G.; Abonyi-Toth, Z.; Rusvai, M.; Palya, V. Comparative Histopathology and Immunohistochemistry of QX-like, Massachusetts and 793/B Serotypes of Infectious Bronchitis Virus Infection in Chickens. J. Comp. Pathol. 2010, 143, 276–283. [Google Scholar] [CrossRef] [PubMed]
- McMartin, D.A.; Macleod, H. Abnormal oviducts in laying hens following natural infection with Infectious Bronchitis at an early age. Br. Vet. J. 1972, 128, XIX–XXI. [Google Scholar] [PubMed]
- Roh, H.J.; Hilt, D.A.; Jackwood, M.W. Detection of infectious bronchitis virus with the use of real-time quantitative reverse transcriptase-PCR and correlation with virus detection in embryonated eggs. Avian Dis. 2014, 58, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Jordan, B. American Association of Avian Pathologists. In Proceedings of the Annual Report of the AAAP Respiratory Disease Committee, Jacksonville, FL, USA, 2–5 August 2019. [Google Scholar]
- Mo, J.; Angelichio, M.; Gow, L.; Leathers, V.; Jackwood, M.W. Validation of specific quantitiative real-time RT-PCR assay panel for Infectious Bronchitis using synthetic DNA standards and clinical specimens. J. Virol. Methods 2020, 276, 113773. [Google Scholar] [CrossRef]
- Wang, F.; Flanagan, J.; Su, N.; Wang, L.C.; Bui, S.; Nielson, A.; Wu, X.; Vo, H.T.; Ma, X.J.; Luo, Y. RNAscope: A novel in situ RNA analysis platform for formalin-fixed, araffin-embedded tissues. J. Mol. Diagn. 2012, 14, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Hassan, M.S.H.; Ali, A.; Buharideen, S.M.; Goldsmith, D.; Coffin, C.S.; Cork, S.C.; van der Meer, F.; Boulianna, M.; Abdul-Careem, M. Pathogenicity of the Canadian Delmarva (DMV/1639) Infectious Bronchitis Virus (IBV) on Female Reproductive Tract of Chickens. Viruses 2021, 13, 2488. [Google Scholar] [CrossRef]
- Crinion, R.A.P.; Hofstad, M.S. Pathogenicity of Four Serotypes of Avian Infectious Bronchitis Virus for the Oviduct of Young Chickens of Various Ages. Avian Dis. 1972, 16, 351–363. [Google Scholar] [CrossRef]
- Bozorgmehri-Fard, M.H.; Charkhkar, S.; Hosseini, H. Detection of the Chinese Genotype of Infectious Bronchitis Virus (QX-type) in Iran. Iran. J. Virol. 2013, 7, 57–60. [Google Scholar] [CrossRef]
- Jones, R.C.; Jordan, F.T.W. Persistence of virus in the tissues and development of the oviduct in the fowl following infection at day old with Infectious Bronchitis Virus. Res. Vet. Sci. 1972, 13, 52–60. [Google Scholar] [CrossRef]
- Crinion, R.A.P. Egg quality and production following Infectious Bronchitis Virus exposure at one day old. Poult. Sci. 1972, 51, 582–585. [Google Scholar] [CrossRef]
- Petrik, M. Personal communication, Zoom Virtual Meeting. 2021.
- Hoerr, F.J. The Pathology of Infectious Bronchitis. Avian Dis. 2021, 65, 600–611. [Google Scholar] [CrossRef] [PubMed]
- Kapczynski, D.R.; Sellers, H.S.; Rowland, G.N.; Jackwood, M.W. Detection of in ovo-inoculated infectious bronchitis virus by immunohistochemistry and in situ hybridization with a riboprobe in epithelial cells of the lung and cloacal bursa. Avian Dis. 2002, 46, 679–685. [Google Scholar] [CrossRef]
- Franca, M.; Woolcock, P.R.; Yu, M.; Jackwood, M.W.; Shivaprasad, H.L. Nephritis associated with infectious bronchitis virus Cal99 variant in game chickens. Avian Dis. 2011, 55, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Braune, M.O.; Gentry, R.F. Standardization of the fluorescent antibody technique for the detection of avian respiratory viruses. Avian Dis. 1965, 9, 535–545. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Challenge | Age at Challenge |
|---|---|---|
| 1 | M41 | 3 days |
| 2 | M41 | 7 days |
| 3 | M41 | 14 days |
| 4 | DMV | 3 days |
| 5 | DMV | 7 days |
| 6 | DMV | 14 days |
| Group/Challenge | Age at Challenge | Mean CT Value ± SEM 7 Days Post-Challenge | % Positive Challenged |
|---|---|---|---|
| 1/M41 | 3 days | 25.19 ± 2.23 | 100% |
| 2/M41 | 7 days | 25.85 ± 1.73 | 99.1% |
| 3/M41 | 14 days | 27.38 ± 2.39 | 100% |
| 4/DMV | 3 days | 24.171 ± 2.83 | 100% |
| 5/DMV | 7 days | 22.47 ± 2.47 | 100% |
| 6/DMV | 14 Days | 25.90 ± 2.08 | 100% |
| Group/Challenge | Age at Challenge | Total % Cystic Oviduct | Total % Affected (including Cystic Oviduct) |
|---|---|---|---|
| 1/M41 | 3 days | 14.78% | 20.87% |
| 2/M41 | 7 days | 32% | 34% |
| 3/M41 | 14 days | 9.91% | 9.91% |
| 4/DMV | 3 days | 24.56% | 28.07% |
| 5/DMV | 7 days | 12.28% | 13.16% |
| 6/DMV | 14 days | 8.55% | 9.40% |
| Score 1 | Inflammation − | Inflammation + | Exudate − | Exudate + | Nodules − | Nodules + |
|---|---|---|---|---|---|---|
| Group 1 | 43.48% | 56.52% | 52.17% | 47.83% | 71.74% | 28.26% |
| Group 2 | 37.5% | 62.50% | 68.75% | 31.25% | 64.58% | 35.42% |
| Group 3 | 38.78% | 61.22% | 53.06% | 46.94% | 71.43% | 28.57% |
| Group 4 | 51.02% | 48.98% | 46.94% | 53.06% | 65.31% | 34.69% |
| Group 5 | 50% | 50% | 70.45% | 29.55% | 68.18% | 31.82% |
| Group 6 | 31.91% | 68.09% | 65.96% | 34.04% | 48.94% | 51.06% |
| Score 1 | Total − | Total + | Total ++ | Total +++ |
|---|---|---|---|---|
| Group 1 | 23.91% | 39.13% | 17.39% | 19.57% |
| Group 2 | 16.67% | 39.58% | 41.67% | 2.08% |
| Group 3 | 16.33% | 42.86% | 28.57% | 12.24% |
| Group 4 | 12.24% | 34.69% | 30.61% | 22.45% |
| Group 5 | 34.09% | 29.55% | 27.27% | 9.09% |
| Group 6 | 14.89% | 40.43% | 27.66% | 17.02% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mueller Slay, A.; Franca, M.; Jackwood, M.; Jordan, B. Infection with IBV DMV/1639 at a Young Age Leads to Increased Incidence of Cystic Oviduct Formation Associated with False Layer Syndrome. Viruses 2022, 14, 852. https://doi.org/10.3390/v14050852
Mueller Slay A, Franca M, Jackwood M, Jordan B. Infection with IBV DMV/1639 at a Young Age Leads to Increased Incidence of Cystic Oviduct Formation Associated with False Layer Syndrome. Viruses. 2022; 14(5):852. https://doi.org/10.3390/v14050852
Chicago/Turabian StyleMueller Slay, Adrea, Monique Franca, Mark Jackwood, and Brian Jordan. 2022. "Infection with IBV DMV/1639 at a Young Age Leads to Increased Incidence of Cystic Oviduct Formation Associated with False Layer Syndrome" Viruses 14, no. 5: 852. https://doi.org/10.3390/v14050852
APA StyleMueller Slay, A., Franca, M., Jackwood, M., & Jordan, B. (2022). Infection with IBV DMV/1639 at a Young Age Leads to Increased Incidence of Cystic Oviduct Formation Associated with False Layer Syndrome. Viruses, 14(5), 852. https://doi.org/10.3390/v14050852