
Prevalence, Genetics and Evolutionary Properties of Eurasian Avian-like H1N1 Swine Influenza Viruses in Liaoning

Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Identification
2.2. Hemagglutination Inhibition Assay
2.3. Sequence Analyses
2.4. Selection Pressure and Positive Selection Sites Estimation
3. Results
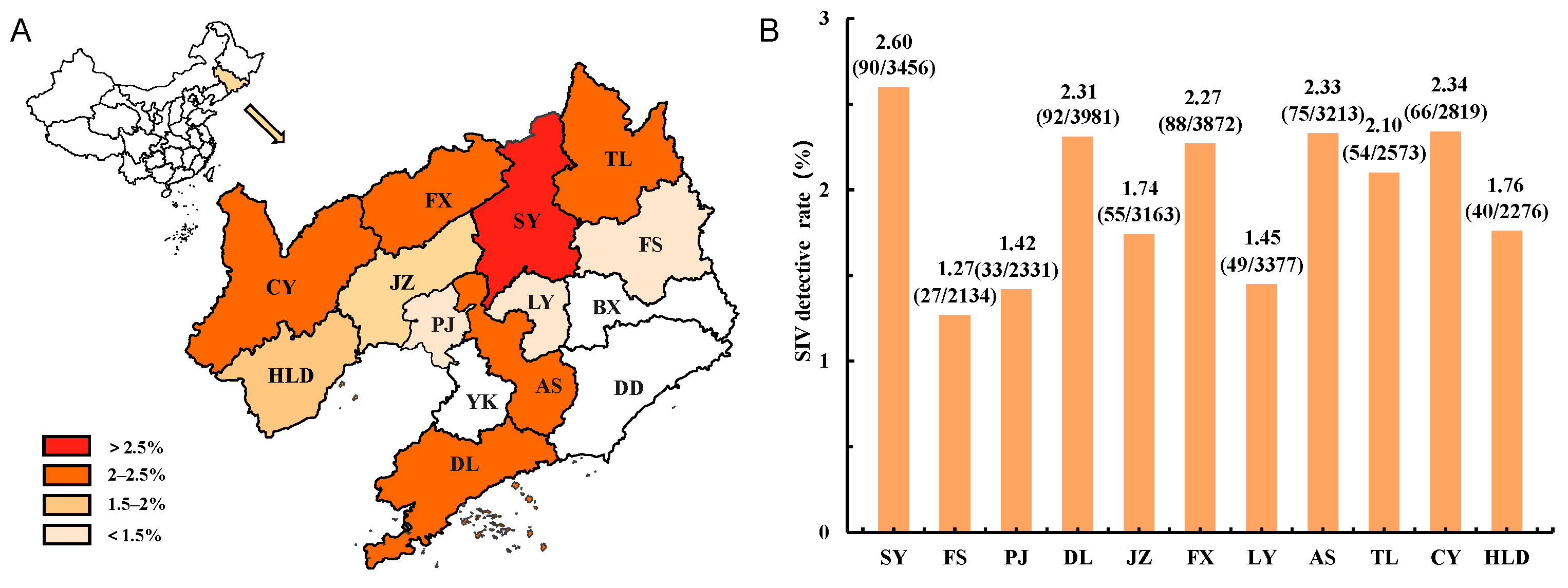
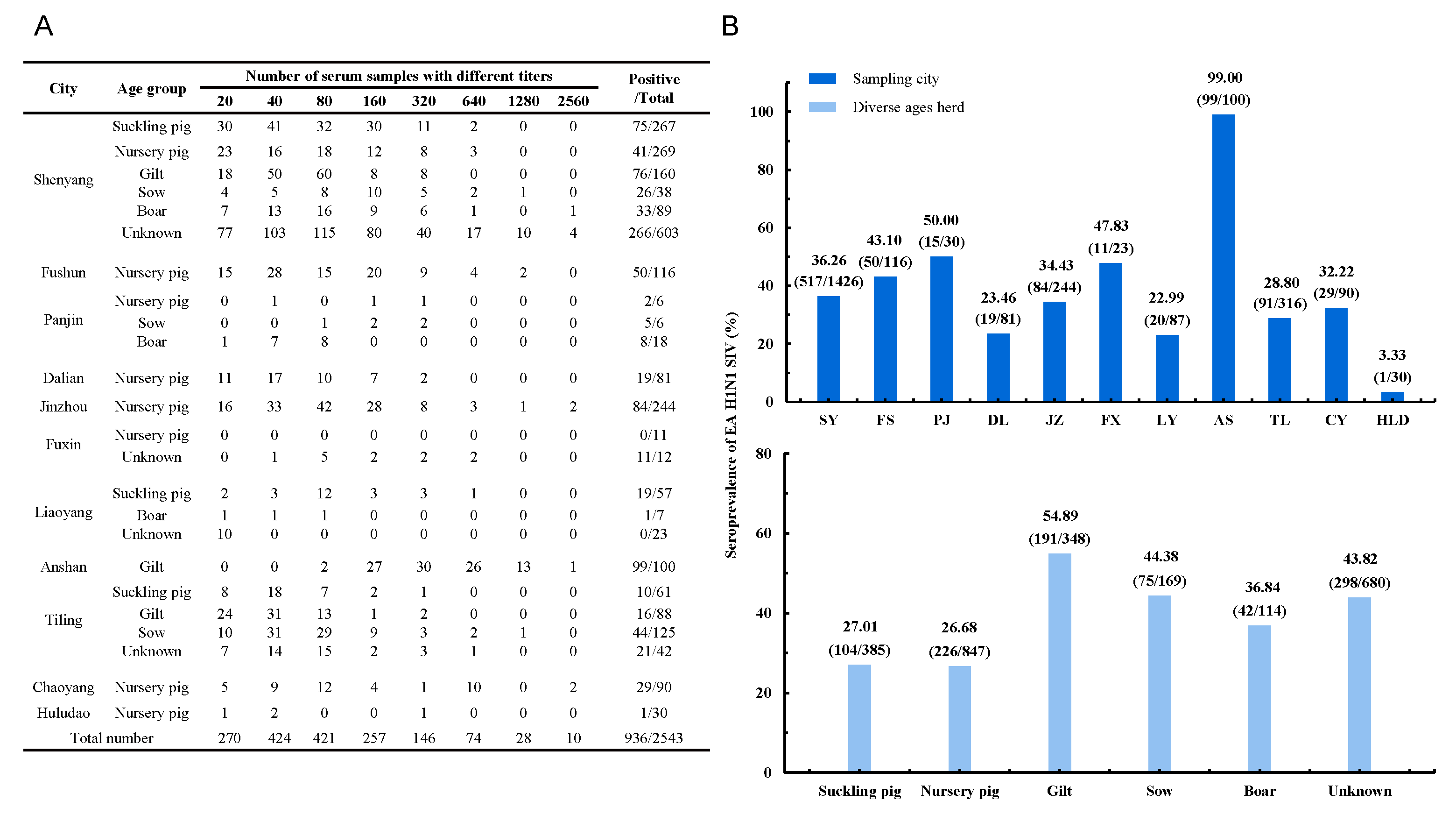
3.1. Epidemic of SIVs
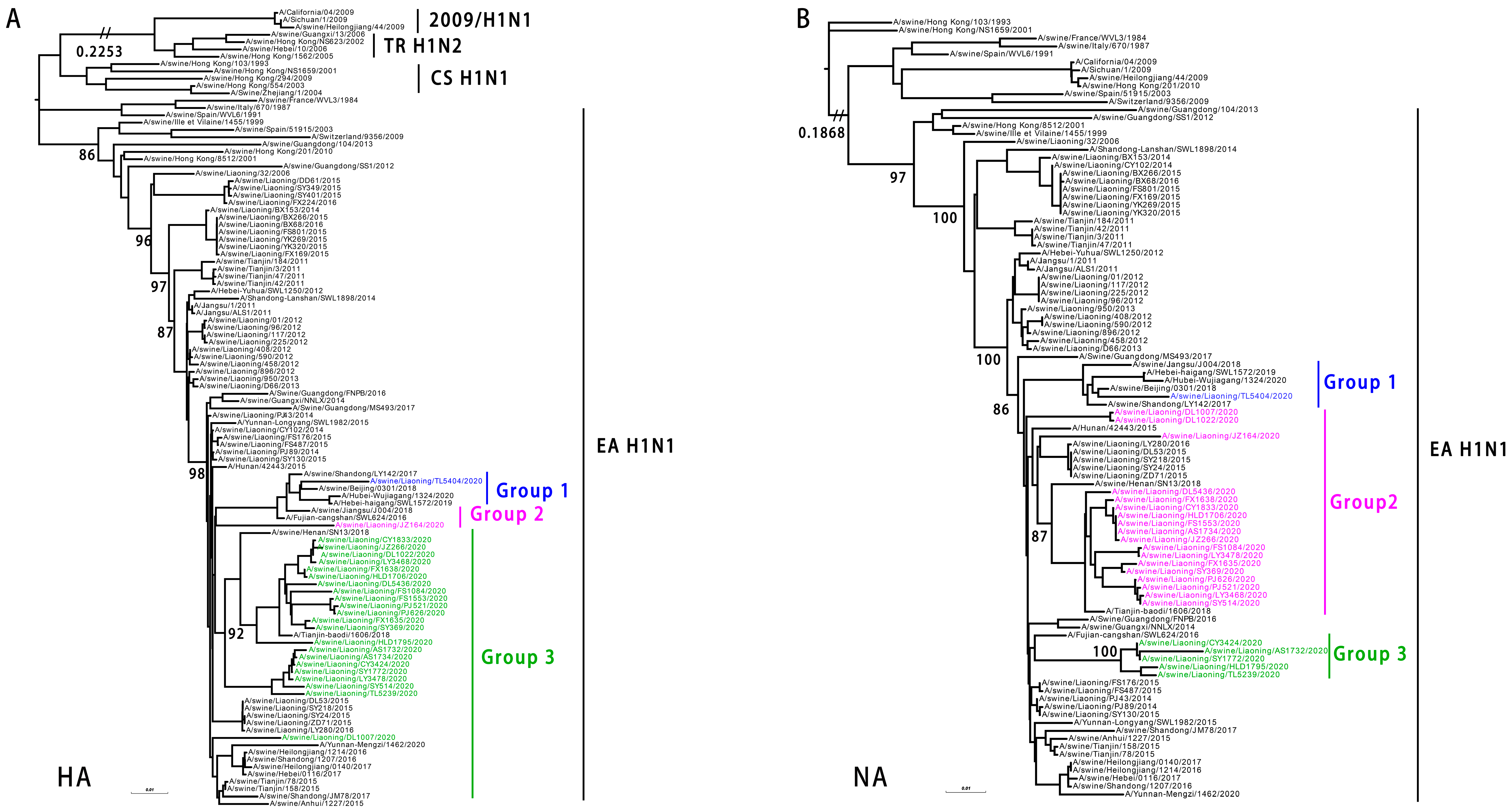
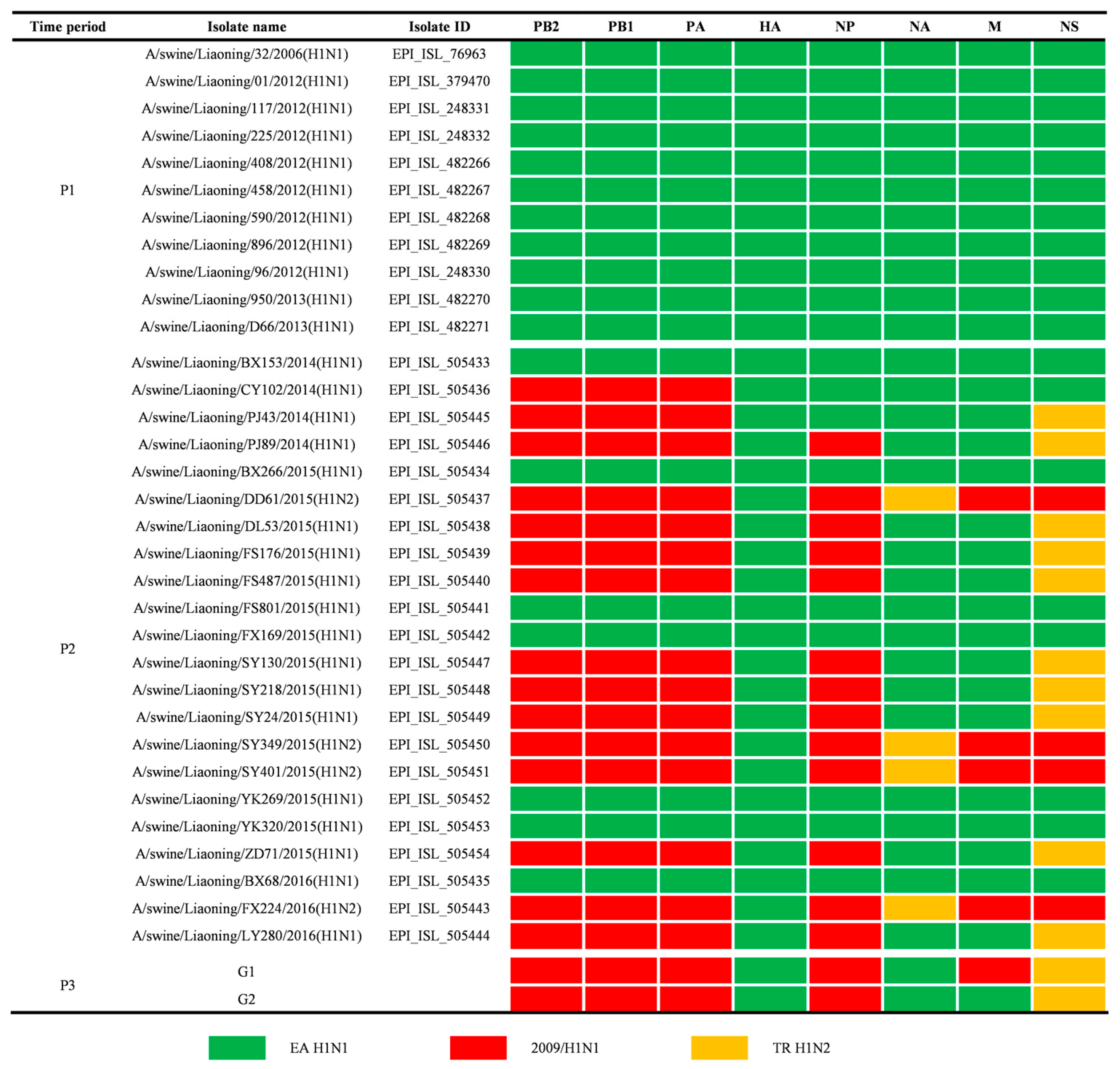
3.2. Genetics of EA H1N1 SIVs
3.3. Molecular Characters of EA H1N1 SIVs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Segments | Amino Acid Mutation | Liaoning 2020 EA H1N1 SIVs |
|---|---|---|
| PB2 | ||
| A combination of 271A, 590S, and 591R plays a critical role in SIV replication and virulence in mammalian host [31] | PB2-T271A-A590S-A591R | PB2-271A-590S-591R |
| Increased viral replication and pathogenicity of EA H1N1 SIVs [32] | PB2-R251K | PB2-251R/K b |
| Enhanced the virulence of EA H1N1 SIVs in mice [13] | PB2-T431M | PB2-431M |
| Enhanced 2009/H1N1 influenza virus virulence [33] | PB2-T588I | PB2-588I |
| Increased avian influenza virus adaptation in mammalian host [34,35] | PB2-E627K | PB2-627E |
| PB2-D701N | PB2-701D | |
| HA a | ||
| Change the antigenicity of EA H1N1 SIVs [30] | HA-E158G | HA-158E/G c |
| Increase the receptor binding affinity to human-type a-2,6-linked sialic acid receptors [29] | HA-E190D | HA-190D |
| Increase the transmissibility of EA H1N1 SIVs in guinea pigs [25] | HA-G225E | HA-225E |
| NP | ||
| A typical human signature marker, which enhances the pathogenicity of EA viruses in mice [36] | NP-Q357K | NP-357K |
| M2 | ||
| Resistance to adamantine derivatives [37] | M2-S31N | M2-31N |
| NS1 | ||
| The key amino acid in regulating the host IFN response and facilitating virus replication [38,39] | NS1-A42S | NS1-42S |
3.4. Evolutionary Properties of EA H1N1 SIVs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ito, T.; Couceiro, J.N.; Kelm, S.; Baum, L.G.; Krauss, S.; Castrucci, M.R.; Donatelli, I.; Kida, H.; Paulson, J.C.; Webster, R.G.; et al. Molecular basis for the generation in pigs of influenza A viruses with pandemic potential. J. Virol. 1998, 72, 7367–7373. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, H.; Wang, W.; Wang, Y.; Han, G.Z.; Chen, H.; Wang, X. A unique feature of swine ANP32A provides susceptibility to avian influenza virus infection in pigs. PLoS Pathog. 2020, 16, e1008330. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Bi, Y.; Qin, K.; Fu, G.; Yang, J.; Peng, J.; Ma, G.; Liu, Q.; Pu, J.; Tian, F. Emergence of European avian influenza virus-like H1N1 swine influenza A viruses in China. J. Clin. Microbiol. 2009, 47, 2643–2646. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Zhu, Q.; Yang, H.; Zhang, X.; Qiao, C.; Chen, Y.; Xin, X.; Chen, H. Two genotypes of H1N2 swine influenza viruses appeared among pigs in China. J. Clin. Virol. 2009, 46, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Ninomiya, A.; Takada, A.; Okazaki, K.; Shortridge, K.F.; Kida, H. Seroepidemiological evidence of avian H4, H5, and H9 influenza A virus transmission to pigs in southeastern China. Vet. Microbiol. 2002, 88, 107–114. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, J.; Qiao, C.; Yang, H.; Zhang, Y.; Xin, X.; Chen, H. Co-circulation of pandemic 2009 H1N1, classical swine H1N1 and avian-like swine H1N1 influenza viruses in pigs in China. Infect. Genet. Evol. 2013, 13, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Brown, I.H. The epidemiology and evolution of influenza viruses in pigs. Vet. Microbiol. 2000, 74, 29–46. [Google Scholar] [CrossRef]
- Webby, R.J.; Webster, R.G. Emergence of influenza A viruses. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2001, 356, 1817–1828. [Google Scholar] [CrossRef]
- Ma, W. Swine influenza virus: Current status and challenge. Virus Res. 2020, 288, 198118. [Google Scholar] [CrossRef]
- Sui, J.; Yang, D.; Qiao, C.; Xu, H.; Xu, B.; Wu, Y.; Yang, H.; Chen, Y.; Chen, H. Protective efficacy of an inactivated Eurasian avian-like H1N1 swine influenza vaccine against homologous H1N1 and heterologous H1N1 and H1N2 viruses in mice. Vaccine 2016, 34, 3757–3763. [Google Scholar] [CrossRef]
- Cao, Z.; Zeng, W.; Hao, X.; Huang, J.; Cai, M.; Zhou, P.; Zhang, G. Continuous evolution of influenza A viruses of swine from 2013 to 2015 in Guangdong, China. PLoS ONE 2019, 14, e0217607. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Xiao, Y.; Liu, J.; Wang, D.; Li, F.; Wang, C.; Li, C.; Zhu, J.; Song, J.; Sun, H.; et al. Prevalent Eurasian avian-like H1N1 swine influenza virus with 2009 pandemic viral genes facilitating human infection. Proc. Natl. Acad. Sci. USA 2020, 117, 17204–17210. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Xu, B.; Wu, Y.; Yang, S.; Jia, Y.; Liang, W.; Yang, D.; He, L.; Zhu, W.; Chen, Y.; et al. A Single Amino Acid at Position 431 of the PB2 Protein Determines the Virulence of H1N1 Swine Influenza Viruses in Mice. J. Virol. 2020, 94, e01930-19. [Google Scholar] [CrossRef] [PubMed]
- Wernike, K.; Beer, M.; Hoffmann, B. Rapid detection of foot-and-mouth disease virus, influenza A virus and classical swine fever virus by high-speed real-time RT-PCR. J. Virol. Methods 2013, 193, 50–54. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hoffmann, E.; Stech, J.; Guan, Y.; Webster, R.G.; Perez, D.R. Universal primer set for the full-length amplification of all influenza A viruses. Arch. Virol. 2001, 146, 2275–2289. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Yang, D.; Xu, B.; Liang, W.; Sui, J.; Chen, Y.; Yang, H.; Chen, H.; Wei, P.; Qiao, C. Immune efficacy of an adenoviral vector-based swine influenza vaccine against antigenically distinct H1N1 strains in mice. Antivir. Res. 2017, 147, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Rowe, T.; Abernathy, R.A.; Hu-Primmer, J.; Thompson, W.W.; Lu, X.; Lim, W.; Fukuda, K.; Cox, N.J.; Katz, J.M. Detection of antibody to avian influenza A (H5N1) virus in human serum by using a combination of serologic assays. J. Clin. Microbiol. 1999, 37, 937–943. [Google Scholar] [CrossRef]
- Karamendin, K.; Kydyrmanov, A.; Kasymbekov, Y.; Daulbayeva, K.; Khan, E.; Seidalina, A.; Sayatov, M. A Highly Pathogenic H5N1 Influenza A Virus Isolated from a Flamingo on the Caspian Sea Shore. Microbiol. Resour. Announc. 2020, 9, e00508-20. [Google Scholar] [CrossRef]
- Lin, Y.P.; Gregory, V.; Collins, P.; Kloess, J.; Wharton, S.; Cattle, N.; Lackenby, A.; Daniels, R.; Hay, A. Neuraminidase receptor binding variants of human influenza A(H3N2) viruses resulting from substitution of aspartic acid 151 in the catalytic site: A role in virus attachment? J. Virol. 2010, 84, 6769–6781. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Delport, W.; Poon, A.F.; Frost, S.D.; Kosakovsky Pond, S.L. Datamonkey 2010: A suite of phylogenetic analysis tools for evolutionary biology. Bioinformatics 2010, 26, 2455–2457. [Google Scholar] [CrossRef] [PubMed]
- Kosakovsky Pond, S.L.; Frost, S.D. Not so different after all: A comparison of methods for detecting amino acid sites under selection. Mol. Biol. Evol. 2005, 22, 1208–1222. [Google Scholar] [CrossRef] [PubMed]
- Murrell, B.; Wertheim, J.O.; Moola, S.; Weighill, T.; Scheffler, K.; Kosakovsky Pond, S.L. Detecting individual sites subject to episodic diversifying selection. PLoS Genet. 2012, 8, e1002764. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yang, H.; Chen, Y.; Tao, S.; Liu, L.; Kong, H.; Ma, S.; Meng, F.; Suzuki, Y.; Qiao, C.; et al. A Single-Amino-Acid Substitution at Position 225 in Hemagglutinin Alters the Transmissibility of Eurasian Avian-like H1N1 Swine Influenza Virus in Guinea Pigs. J. Virol. 2017, 91, e00800-17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Q.; Gao, Y.; He, X.; Kong, H.; Jiang, Y.; Guan, Y.; Xia, X.; Shu, Y.; Kawaoka, Y.; et al. Key molecular factors in hemagglutinin and PB2 contribute to efficient transmission of the 2009 H1N1 pandemic influenza virus. J. Virol. 2012, 86, 9666–9674. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, Y.; Shinya, K.; Deng, G.; Jiang, Y.; Li, Z.; Guan, Y.; Tian, G.; Li, Y.; Shi, J.; et al. Identification of amino acids in HA and PB2 critical for the transmission of H5N1 avian influenza viruses in a mammalian host. PLoS Pathog. 2009, 5, e1000709. [Google Scholar] [CrossRef]
- Yang, H.; Chen, Y.; Qiao, C.; He, X.; Zhou, H.; Sun, Y.; Yin, H.; Meng, S.; Liu, L.; Zhang, Q.; et al. Prevalence, genetics, and transmissibility in ferrets of Eurasian avian-like H1N1 swine influenza viruses. Proc. Natl. Acad. Sci. USA 2016, 113, 392–397. [Google Scholar] [CrossRef]
- Matrosovich, M.; Tuzikov, A.; Bovin, N.; Gambaryan, A.; Klimov, A.; Castrucci, M.R.; Donatelli, I.; Kawaoka, Y. Early alterations of the receptor-binding properties of H1, H2, and H3 avian influenza virus hemagglutinins after their introduction into mammals. J. Virol. 2000, 74, 8502–8512. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Y.; Chen, H.; Meng, F.; Tao, S.; Ma, S.; Qiao, C.; Chen, H.; Yang, H. A single amino acid at position 158 in haemagglutinin affects the antigenic property of Eurasian avian-like H1N1 swine influenza viruses. Transbound Emerg. Dis. 2021; in press. [Google Scholar] [CrossRef]
- Liu, Q.; Qiao, C.; Marjuki, H.; Bawa, B.; Ma, J.; Guillossou, S.; Webby, R.J.; Richt, J.A.; Ma, W. Combination of PB2 271A and SR polymorphism at positions 590/591 is critical for viral replication and virulence of swine influenza virus in cultured cells and in vivo. J. Virol. 2012, 86, 1233–1237. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Zhong, R.; Qin, C.; Yu, Z.; Wen, X.; Xian, J.; Chen, Y.; Cai, Y.; Yi, H.; Gong, L.; et al. The R251K Substitution in Viral Protein PB2 Increases Viral Replication and Pathogenicity of Eurasian Avian-like H1N1 Swine Influenza Viruses. Viruses 2020, 12, 52. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Yi, C.; Zhao, L.; Wang, S.; Zhou, L.; Hu, Y.; Zou, W.; Chen, H.; Jin, M. PB2-588I enhances 2009 H1N1 pandemic influenza virus virulence by increasing viral replication and exacerbating PB2 inhibition of beta interferon expression. J. Virol. 2014, 88, 2260–2267. [Google Scholar] [CrossRef] [PubMed]
- Hatta, M.; Gao, P.; Halfmann, P.; Kawaoka, Y. Molecular basis for high virulence of Hong Kong H5N1 influenza A viruses. Science 2001, 293, 1840–1842. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Chen, H.; Jiao, P.; Deng, G.; Tian, G.; Li, Y.; Hoffmann, E.; Webster, R.G.; Matsuoka, Y.; Yu, K. Molecular basis of replication of duck H5N1 influenza viruses in a mammalian mouse model. J. Virol. 2005, 79, 12058–12064. [Google Scholar] [CrossRef]
- Zhu, W.; Feng, Z.; Chen, Y.; Yang, L.; Liu, J.; Li, X.; Liu, S.; Zhou, L.; Wei, H.; Gao, R.; et al. Mammalian-adaptive mutation NP-Q357K in Eurasian H1N1 Swine Influenza viruses determines the virulence phenotype in mice. Emerg. Microbes Infect. 2019, 8, 989–999. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, Y.; Ma, C.; Fiorin, G.; Wang, J.; Pinto, L.H.; Lamb, R.A.; Klein, M.L.; Degrado, W.F. Structure and inhibition of the drug-resistant S31N mutant of the M2 ion channel of influenza A virus. Proc. Natl. Acad. Sci. USA 2013, 110, 1315–1320. [Google Scholar] [CrossRef]
- Cheng, J.; Zhang, C.; Tao, J.; Li, B.; Shi, Y.; Liu, H. Effects of the S42 residue of the H1N1 swine influenza virus NS1 protein on interferon responses and virus replication. Virol. J. 2018, 15, 57. [Google Scholar] [CrossRef]
- Jiao, P.; Tian, G.; Li, Y.; Deng, G.; Jiang, Y.; Liu, C.; Liu, W.; Bu, Z.; Kawaoka, Y.; Chen, H. A single-amino-acid substitution in the NS1 protein changes the pathogenicity of H5N1 avian influenza viruses in mice. J. Virol. 2008, 82, 1146–1154. [Google Scholar] [CrossRef]
- Li, X.; Shi, J.; Guo, J.; Deng, G.; Zhang, Q.; Wang, J.; He, X.; Wang, K.; Chen, J.; Li, Y.; et al. Genetics, receptor binding property, and transmissibility in mammals of naturally isolated H9N2 Avian Influenza viruses. PLoS Pathog. 2014, 10, e1004508. [Google Scholar] [CrossRef]
- Suzuki, Y.; Uchida, Y.; Tanikawa, T.; Maeda, N.; Takemae, N.; Saito, T. Amino acid substitutions in PB1 of avian influenza viruses influence pathogenicity and transmissibility in chickens. J. Virol. 2014, 88, 11130–11139. [Google Scholar] [CrossRef] [PubMed]
- Yamaji, R.; Yamada, S.; Le, M.Q.; Li, C.; Chen, H.; Qurnianingsih, E.; Nidom, C.A.; Ito, M.; Sakai-Tagawa, Y.; Kawaoka, Y. Identification of PB2 mutations responsible for the efficient replication of H5N1 influenza viruses in human lung epithelial cells. J. Virol. 2015, 89, 3947–3956. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.Y.; Li, O.T.; Tang, W.P.; Hu, C.; Wang, G.X.; Ngo, J.C.; Wan, D.C.; Poon, L.L.; Shaw, P.C. Identification of influenza polymerase inhibitors targeting C-terminal domain of PA through surface plasmon resonance screening. Sci. Rep. 2018, 8, 2280. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Riedel, K.; Lynch, P.; Chien, C.Y.; Montelione, G.T.; Krug, R.M. RNA binding by the novel helical domain of the influenza virus NS1 protein requires its dimer structure and a small number of specific basic amino acids. RNA 1999, 5, 195–205. [Google Scholar] [CrossRef]
- Qiao, C.; Liu, L.; Yang, H.; Chen, Y.; Xu, H.; Chen, H. Novel triple reassortant H1N2 influenza viruses bearing six internal genes of the pandemic 2009/H1N1 influenza virus were detected in pigs in China. J. Clin. Virol. 2014, 61, 529–534. [Google Scholar] [CrossRef]
- Yang, H.; Chen, Y.; Shi, J.; Guo, J.; Xin, X.; Zhang, J.; Wang, D.; Shu, Y.; Qiao, C.; Chen, H. Reassortant H1N1 influenza virus vaccines protect pigs against pandemic H1N1 influenza virus and H1N2 swine influenza virus challenge. Vet. Microbiol. 2011, 152, 229–234. [Google Scholar] [CrossRef]
- Vincent, A.; Awada, L.; Brown, I.; Chen, H.; Claes, F.; Dauphin, G.; Donis, R.; Culhane, M.; Hamilton, K.; Lewis, N.; et al. Review of influenza A virus in swine worldwide: A call for increased surveillance and research. Zoonoses Public Health 2014, 61, 4–17. [Google Scholar] [CrossRef]
- Yang, H.; Qiao, C.; Tang, X.; Chen, Y.; Xin, X.; Chen, H. Human infection from avian-like influenza A (H1N1) viruses in pigs, China. Emerg. Infect. Dis 2012, 18, 1144–1146. [Google Scholar] [CrossRef]
- Li, C.; Chen, H. Enhancement of influenza virus transmission by gene reassortment. Curr. Top. Microbiol. Immunol. 2014, 385, 185–204. [Google Scholar] [CrossRef]
- Jeffares, D.C.; Tomiczek, B.; Sojo, V.; dos Reis, M. A beginners guide to estimating the non-synonymous to synonymous rate ratio of all protein-coding genes in a genome. Methods Mol. Biol. 2015, 1201, 65–90. [Google Scholar] [CrossRef]





| Virus | Sample Information | |||
|---|---|---|---|---|
| No. | Full Name | Abbreviation | Collected Date | Location |
| 1 | A/swine/Liaoning/JZ164/2020 (H1N1) | JZ164 | 19 January 2020 | Jinzhou |
| 2 | A/swine/Liaoning/JZ266/2020 (H1N1) | JZ266 | 12 January 2020 | Jinzhou |
| 3 | A/swine/Liaoning/SY369/2020 (H1N1) | SY369 | 14 February 2020 | Shenyang |
| 4 | A/swine/Liaoning/SY514/2020 (H1N1) | SY514 | 11 September 2020 | Shenyang |
| 5 | A/swine/Liaoning/PJ521/2020 (H1N1) | PJ521 | 25 March 2020 | Panjin |
| 6 | A/swine/Liaoning/PJ626/2020 (H1N1) | PJ626 | 20 March 2020 | Panjin |
| 7 | A/swine/Liaoning/DL1007/2020 (H1N1) | DL1007 | 21 April 2020 | Dalian |
| 8 | A/swine/Liaoning/DL1022/2020 (H1N1) | DL1022 | 15 January 2020 | Dalian |
| 9 | A/swine/Liaoning/FS1553/2020 (H1N1) | FS1553 | 17 May 2020 | Fushun |
| 10 | A/swine/Liaoning/FS1084/2020 (H1N1) | FS1084 | 10 May 2020 | Fushun |
| 11 | A/swine/Liaoning/FX1635/2020 (H1N1) | FX1635 | 11 June 2020 | Fuxin |
| 12 | A/swine/Liaoning/FX1638/2020 (H1N1) | FX1638 | 18 June 2020 | Fuxin |
| 13 | A/swine/Liaoning/HLD1706/2020 (H1N1) | HLD1706 | 27 July 2020 | Huludao |
| 14 | A/swine/Liaoning/AS1732/2020 (H1N1) | AS1732 | 27 July 2020 | Anshan |
| 15 | A/swine/Liaoning/AS1734/2020 (H1N1) | AS1734 | 9 August 2020 | Anshan |
| 16 | A/swine/Liaoning/SY1772/2020 (H1N1) | SY1772 | 11 August 2020 | Shenyang |
| 17 | A/swine/Liaoning/CY1833/2020 (H1N1) | CY1833 | 15 September 2020 | Chaoyang |
| 18 | A/swine/Liaoning/HLD1795/2020 (H1N1) | HLD1795 | 15 September 2020 | Huludao |
| 19 | A/swine/Liaoning/CY3424/2020 (H1N1) | CY3424 | 8 October 2020 | Chaoyang |
| 20 | A/swine/Liaoning/LY3468/2020 (H1N1) | LY3468 | 3 August 2020 | Liaoyang |
| 21 | A/swine/Liaoning/TL5239/2020 (H1N1) | TL5239 | 12 November 2020 | Tieling |
| 22 | A/swine/Liaoning/LY3478/2020 (H1N1) | LY3478 | 12 November 2020 | Liaoyang |
| 23 | A/swine/Liaoning/TL5404/2020 (H1N1) | TL5404 | 14 December 2020 | Tieling |
| 24 | A/swine/Liaoning/DL5436/2020 (H1N1) | DL5436 | 14 December 2020 | Dalian |
| Gene Segment | Positive Selection Sites of 2020 EA H1N1 SIVs | Selection Pressure (dN/dS) | ||
|---|---|---|---|---|
| 2020 a | 2014–2016 | 2006–2013 | ||
| HA b | 60, 219, 361 | 0.193 | 0.160 | 0.215 |
| NA c | - | 0.209 | 0.164 | 0.190 |
| PB2 | 391, 661 | 0.125 | - | - |
| PB1 | 14, 364, 607, 673, 683 | 0.135 | - | - |
| PA | 436, 532 | 0.163 | - | - |
| NP | 113 | 0.121 | - | - |
| M1 | - | 0.243 | - | - |
| M2 | - | 0.491 | - | - |
| NS1 | 4, 51 | 0.539 | - | - |
| NS2 | - | 0.253 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Leng, H.; Tang, S.; Su, C.; Xu, Y.; Wang, Y.; Lv, J.; Zhang, S.; Feng, Y.; Song, S.; et al. Prevalence, Genetics and Evolutionary Properties of Eurasian Avian-like H1N1 Swine Influenza Viruses in Liaoning. Viruses 2022, 14, 643. https://doi.org/10.3390/v14030643
Li H, Leng H, Tang S, Su C, Xu Y, Wang Y, Lv J, Zhang S, Feng Y, Song S, et al. Prevalence, Genetics and Evolutionary Properties of Eurasian Avian-like H1N1 Swine Influenza Viruses in Liaoning. Viruses. 2022; 14(3):643. https://doi.org/10.3390/v14030643
Chicago/Turabian StyleLi, Hailing, Haoyu Leng, Siqi Tang, Chaofan Su, Yina Xu, Yongtao Wang, Jiaming Lv, Shiwei Zhang, Yali Feng, Shaokang Song, and et al. 2022. "Prevalence, Genetics and Evolutionary Properties of Eurasian Avian-like H1N1 Swine Influenza Viruses in Liaoning" Viruses 14, no. 3: 643. https://doi.org/10.3390/v14030643
APA StyleLi, H., Leng, H., Tang, S., Su, C., Xu, Y., Wang, Y., Lv, J., Zhang, S., Feng, Y., Song, S., & Zhang, Y. (2022). Prevalence, Genetics and Evolutionary Properties of Eurasian Avian-like H1N1 Swine Influenza Viruses in Liaoning. Viruses, 14(3), 643. https://doi.org/10.3390/v14030643