
Polysaccharides and Their Derivatives as Potential Antiviral Molecules


 ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Commercial Antiviral Compounds
2.1. History of Antiviral Drugs
2.2. Development of Antiviral Drugs
2.3. Different Types of Antivirals
2.3.1. Cell Entry Inhibitors
2.3.2. Replication Inhibitors
2.3.3. Post-Replication Inhibitors
3. Polysaccharides with Antiviral Activities
3.1. Identification of the Main Polysaccharides with Antiviral Properties
3.1.1. Antiviral Activities of Polysaccharides That Have Animal Origins
3.1.2. Antiviral Activities of Polysaccharides from Seaweeds, Mushrooms, and Plants
3.1.3. Antiviral Activities of Microbial Polysaccharides
3.1.4. Antiviral Activities of Polysaccharide Derivatives
3.2. Structure–Activity Relationships
3.3. The Mechanisms of Antiviral Polysaccharides
3.3.1. Killing the Virus Directly
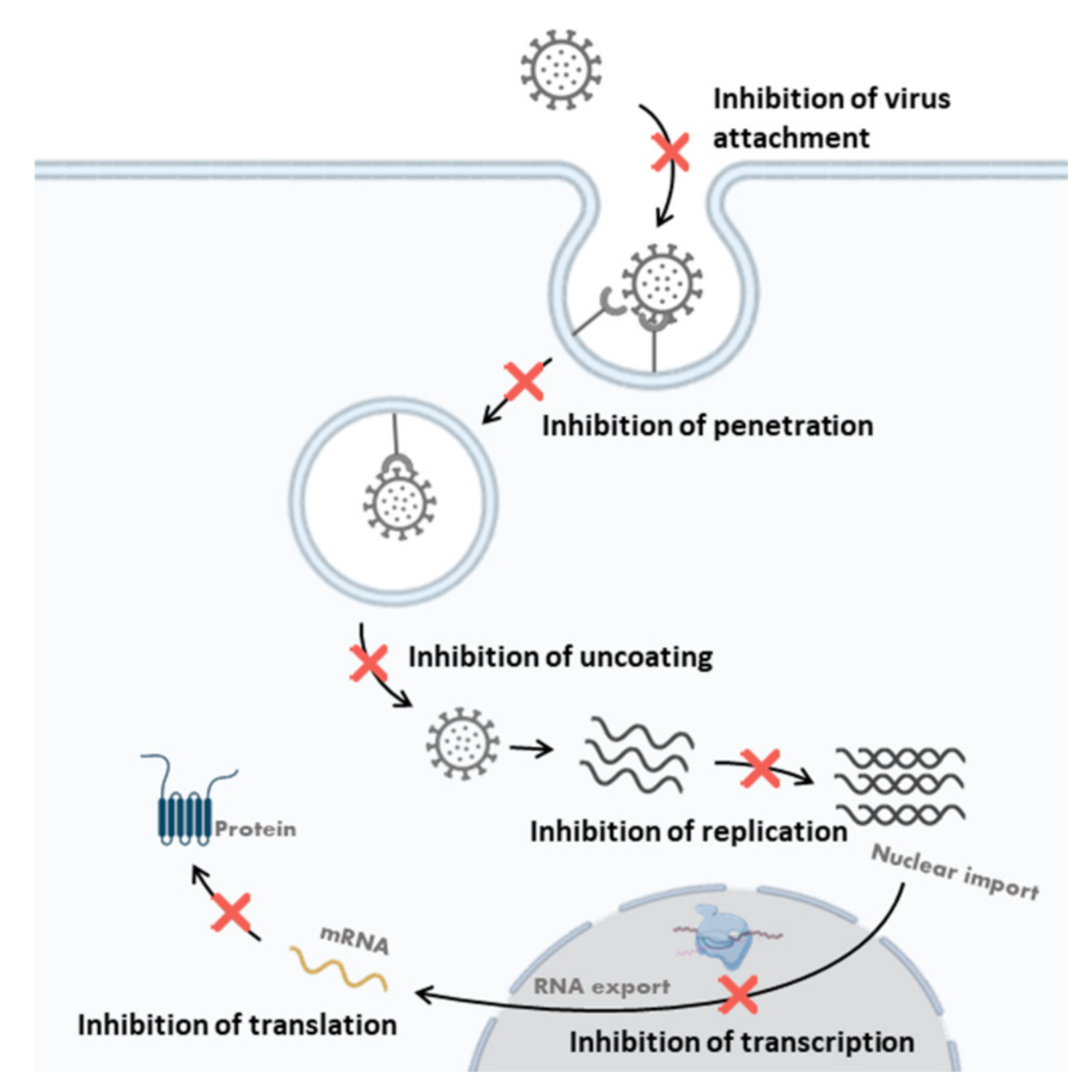
3.3.2. Inhibition of Virus Attachment
3.3.3. Inhibition of Penetration and Uncoating
3.3.4. Inhibition of Viral Transcription and Replication
4. Conclusions and Future Prospect(s)
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mazur-Marzec, H.; Cegłowska, M.; Konkel, R.; Pyrć, K. Antiviral Cyanometabolites—A Review. Biomolecules 2021, 11, 474. [Google Scholar] [CrossRef] [PubMed]
- Watson, D.G. Pharmaceutical Chemistry, International Edition E-Book; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Moelling, K.; Broecker, F. Viruses and Evolution—Viruses First? A Personal Perspective. Front. Microbiol. 2019, 10, 523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suttle, C.A. Viruses in the Sea. Nature 2005, 437, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Xu, T.; Wang, C.; Jia, T.; Zhang, Q. A Novel Picornavirus Discovered in White Leg Shrimp Penaeus vannamei. Viruses 2021, 13, 2381. [Google Scholar] [CrossRef]
- Sarker, S.; Bowden, T.R.; Boyle, D.B. Evidence of a Possible Viral Host Switch Event in an Avipoxvirus Isolated from an Endangered Northern Royal Albatross (Diomedea sanfordi). Viruses 2022, 14, 302. [Google Scholar] [CrossRef]
- Stokholm, I.; Fischer, N.; Baechlein, C.; Postel, A.; Galatius, A.; Kyhn, L.A.; Thøstesen, C.B.; Persson, S.; Siebert, U.; Olsen, M.T.; et al. In the Search of Marine Pestiviruses: First Case of Phocoena pestivirus in a Belt Sea Harbour Porpoise. Viruses 2022, 14, 161. [Google Scholar] [CrossRef]
- Liu, W.; Zheng, X.; Dai, X.; Zhang, Z.; Zhang, W.; Xiao, T.; Huang, L. Isolation and Characterization of the First Temperate Virus Infecting Psychrobacillus from Marine Sediments. Viruses 2022, 14, 108. [Google Scholar] [CrossRef]
- Akashi, M.; Takemura, M. Co-Isolation and Characterization of Two Pandoraviruses and a Mimivirus from a Riverbank in Japan. Viruses 2019, 11, 1123. [Google Scholar] [CrossRef] [Green Version]
- Furmaga, J.; Kowalczyk, M.; Furmaga, O.; Rokos, C.A.; Zapolski, T.; Krakowski, L.; Jakubczak, A.; Rudzki, S. Molecular Epidemiology and Variation of the BK Polyomavirus in the Population of Central and Eastern Europe Based on the Example of Poland. Viruses 2022, 14, 209. [Google Scholar] [CrossRef]
- Banyard, A.C.; Lean, F.Z.X.; Robinson, C.; Howie, F.; Tyler, G.; Nisbet, C.; Seekings, J.; Meyer, S.; Whittard, E.; Ashpitel, H.F.; et al. Detection of Highly Pathogenic Avian Influenza Virus H5N1 Clade 2.3.4.4b in Great Skuas: A Species of Conservation Concern in Great Britain. Viruses 2022, 14, 212. [Google Scholar] [CrossRef]
- Hancková, M.; Betáková, T. Pandemics of the 21st Century: The Risk Factor for Obese People. Viruses 2021, 14, 25. [Google Scholar] [CrossRef] [PubMed]
- Tumpey, T.M.; Basler, C.F.; Aguilar, P.V.; Zeng, H.; Solórzano, A.; Swayne, D.E.; Cox, N.J.; Katz, J.M.; Taubenberger, J.K.; Palese, P.; et al. Characterization of the Reconstructed 1918 Spanish Influenza Pandemic Virus. Science 2005, 310, 77–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loo, Y.-M.; Gale, M. Fatal Immunity and the 1918 Virus. Nature 2007, 445, 267–268. [Google Scholar] [CrossRef] [PubMed]
- Kalinichenko, S.; Komkov, D.; Mazurov, D. HIV-1 and HTLV-1 Transmission Modes: Mechanisms and Importance for Virus Spread. Viruses 2022, 14, 152. [Google Scholar] [CrossRef] [PubMed]
- Alene, K.A.; Wangdi, K.; Clements, A.C.A. Impact of the COVID-19 Pandemic on Tuberculosis Control: An Overview. Trop. Med. Infect. Dis. 2020, 5, 123. [Google Scholar] [CrossRef]
- Timeline: Swine Flu. Nature 2009. Available online: https://www.nature.com/articles/news.2009.416#appendices (accessed on 10 June 2021).
- Rainwater-Lovett, K.; Rodriguez-Barraquer, I.; Moss, W.J. Viral Epidemiology. In Viral Pathogenesis; Elsevier: Amsterdam, The Netherlands, 2016; pp. 241–252. ISBN 978-0-12-800964-2. [Google Scholar]
- Bos, L. Beijerinck’s Work on Tobacco Mosaic Virus: Historical Context and Legacy. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1999, 354, 675–685. [Google Scholar] [CrossRef] [Green Version]
- Roingeard, P. Viral Detection by Electron Microscopy: Past, Present and Future. Biol. Cell 2008, 100, 491–501. [Google Scholar] [CrossRef]
- Goldsmith, C.S.; Miller, S.E. Modern Uses of Electron Microscopy for Detection of Viruses. Clin. Microbiol. Rev. 2009, 22, 552–563. [Google Scholar] [CrossRef] [Green Version]
- De Clercq, E. Milestones in the Discovery of Antiviral Agents: Nucleosides and Nucleotides. Acta Pharm. Sin. B 2012, 2, 535–548. [Google Scholar] [CrossRef] [Green Version]
- De Clercq, E.; Field, H.J. Antiviral Prodrugs—The Development of Successful Prodrug Strategies for Antiviral Chemotherapy: Antiviral Prodrugs. Br. J. Pharmacol. 2006, 147, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Clercq, E.D. Antivirals and Antiviral Strategies. Nat. Rev. Microbiol. 2004, 2, 704–720. [Google Scholar] [CrossRef] [PubMed]
- Banik, N.; Yang, S.-B.; Kang, T.-B.; Lim, J.-H.; Park, J. Heparin and Its Derivatives: Challenges and Advances in Therapeutic Biomolecules. Int. J. Mol. Sci. 2021, 22, 10524. [Google Scholar] [CrossRef] [PubMed]
- Lim, G.B. Discovery and Purification of Heparin. Nat. Rev. Cardiol. 2017. Available online: https://www.nature.com/articles/nrcardio.2017.171#citeas (accessed on 10 June 2021).
- Andrew, M.; Jayaraman, G. Marine Sulfated Polysaccharides as Potential Antiviral Drug Candidates to Treat Corona Virus Disease (COVID-19). Carbohydr. Res. 2021, 505, 108326. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.J.A.; Olmo, L.M.B.D.; Benito, P.B. Antiviral Activities of Polysaccharides from Natural Sources. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2005; Volume 30, pp. 393–418. ISBN 978-0-444-51854-5. [Google Scholar]
- Chen, L.; Huang, G. The Antiviral Activity of Polysaccharides and Their Derivatives. Int. J. Biol. Macromol. 2018, 115, 77–82. [Google Scholar] [CrossRef]
- Meyer, H.; Ehmann, R.; Smith, G.L. Smallpox in the Post-Eradication Era. Viruses 2020, 12, 138. [Google Scholar] [CrossRef] [Green Version]
- Belongia, E.A.; Naleway, A.L. Smallpox Vaccine: The Good, the Bad, and the Ugly. Clin. Med. Res. 2003, 1, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Kausar, S.; Said Khan, F.; Ishaq Mujeeb Ur Rehman, M.; Akram, M.; Riaz, M.; Rasool, G.; Hamid Khan, A.; Saleem, I.; Shamim, S.; Malik, A. A Review: Mechanism of Action of Antiviral Drugs. Int. J. Immunopathol. Pharmacol. 2021, 35, 205873842110026. [Google Scholar] [CrossRef]
- Bray, M. Highly Pathogenic RNA Viral Infections: Challenges for Antiviral Research. Antivir. Res. 2008, 78, 1–8. [Google Scholar] [CrossRef]
- da Silva, M.; Marin, P.; Repolês, B.; Elias, M.; Machado, C. Analysis of DNA Exchange Using Thymidine Analogs (ADExTA) in Trypanosoma Cruzi. BIO-Protocol 2018, 8, e3125. [Google Scholar] [CrossRef]
- Prusoff, W.H. Synthesis and Biological Activities of Iododeoxyuridine, an Analog of Thymidine. Biochim. Biophys. Acta 1959, 32, 295–296. [Google Scholar] [CrossRef]
- Hamre, D.; Brownlee, K.A.; Donovick, R. Studies on the Chemotherapy of Vaccinia Virus. II. The Activity of Some Thiosemicarbazones. J. Immunol. 1951, 67, 305–312. [Google Scholar] [PubMed]
- Hamre, D.; Bernstein, J.; Donovick, R. Activity of P-Aminobenzaldehyde, 3-Thiosemicarbazone on Vaccinia Virus in the Chick Embryo and in the Mouse. Exp. Biol. Med. 1950, 73, 275–278. [Google Scholar] [CrossRef]
- Bauer, D.J.; Stvincent, L.; Kempe, C.H.; Downie, A.W. Prophylactic treatment of small pox contacts with n-methylisatin beta-thiosemicarbazone (compound 33t57, marboran). Lancet Lond. Engl. 1963, 2, 494–496. [Google Scholar] [CrossRef]
- Elion, G.B.; Furman, P.A.; Fyfe, J.A.; Miranda, P.D.; Beauchamp, L.; Schaeffer, H.J. Selectivity of Action of an Antiherpetic Agent, 9-(2-Hydroxyethoxymethyl)Guanine. Proc. Natl. Acad. Sci. USA 1977, 74, 5716–5720. [Google Scholar] [CrossRef] [Green Version]
- Derse, D.; Cheng, Y.C.; Furman, P.A.; St Clair, M.H.; Elion, G.B. Inhibition of Purified Human and Herpes Simplex Virus-Induced DNA Polymerases by 9-(2-Hydroxyethoxymethyl)Guanine Triphosphate. Effects on Primer-Template Function. J. Biol. Chem. 1981, 256, 11447–11451. [Google Scholar] [CrossRef]
- Clercq, E.D. Looking Back in 2009 at the Dawning of Antiviral Therapy Now 50 Years Ago. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2009; Volume 73, Chapter 1; pp. 1–53. ISBN 978-0-12-374786-0. [Google Scholar]
- Gayle, H.D.; Hill, G.L. Global Impact of Human Immunodeficiency Virus and AIDS. Clin. Microbiol. Rev. 2001, 14, 327–335. [Google Scholar] [CrossRef] [Green Version]
- Li, A.P. Screening for Human ADME/Tox Drug Properties in Drug Discovery. Drug Discov. Today 2001, 6, 357–366. [Google Scholar] [CrossRef]
- Vidal, L.; Msellati, P.; Abidjan, C.D.I. Sida Dans Les Pays En Développement. Actual. Doss. St. Publique 2002, 40, 37–42. [Google Scholar]
- Dolin, R.; Morgan, J. Antiviral Agents: General Principles. In Mandell, Douglas, and Bensnett’s Principles and Practice of Infec-Tious Diseases; Elsevier: Amsterdam, The Netherlands, 2015; pp. 528–530. [Google Scholar]
- Popping, S.; Cento, V.; Seguin-Devaux, C.; Boucher, C.; de Salazar, A.; Heger, E.; Mor, O.; Sayan, M.; Salmon-Ceron, D.; Weis, N.; et al. The European Prevalence of Resistance Associated Substitutions among Direct Acting Antiviral Failures. Viruses 2021, 14, 16. [Google Scholar] [CrossRef]
- Colpitts, C.; Baumert, T. Addressing the Challenges of Hepatitis C Virus Resistance and Treatment Failure. Viruses 2016, 8, 226. [Google Scholar] [CrossRef] [Green Version]
- Strasfeld, L.; Chou, S. Antiviral Drug Resistance: Mechanisms and Clinical Implications. Infect. Dis. Clin. N. Am. 2010, 24, 413–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agut, H.; Burrel, S.; Bonnafous, P.; Boutolleau, D. Où en est la recherche sur les antiviraux? Rev. Prat. 2016, 66, 1007–1014. [Google Scholar] [PubMed]
- Salvaggio, M.R.; Gnann, J.W. Drugs for Herpesvirus Infections. In Infectious Diseases; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1309–1317.e1. ISBN 978-0-7020-6285-8. [Google Scholar]
- De Clercq, E. Strategies in the Design of Antiviral Drugs. Nat. Rev. Drug Discov. 2002, 1, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Passman, A.M.; Strauss, R.P.; McSpadden, S.B.; Finch-Edmondson, M.; Andrewartha, N.; Woo, K.H.; Diepeveen, L.A.; Zhao, W.; Fernández-Irigoyen, J.; Santamaría, E.; et al. Maraviroc Prevents HCC Development by Suppressing Macrophages and the Liver Progenitor Cell Response in a Murine Chronic Liver Disease Model. Cancers 2021, 13, 4935. [Google Scholar] [CrossRef] [PubMed]
- Lalezari, J.P.; Henry, K.; O’Hearn, M.; Montaner, J.S.; Piliero, P.J.; Trottier, B.; Walmsley, S.; Cohen, C.; Kuritzkes, D.R.; Eron, J.J., Jr.; et al. Enfuvirtide, an HIV-1 Fusion Inhibitor, for Drug-Resistant HIV Infection in North and South America. N. Engl. J. Med. 2003, 348, 2175–2185. [Google Scholar] [CrossRef] [Green Version]
- Ratcliff, A.N.; Shi, W.; Arts, E.J. HIV-1 Resistance to Maraviroc Conferred by a CD4 Binding Site Mutation in the Envelope Glycoprotein Gp120. J. Virol. 2013, 87, 923–934. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, M.L. Resistance to Enfuvirtide, the First HIV Fusion Inhibitor. J. Antimicrob. Chemother. 2004, 54, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Lüscher-Mattii, M. Polyanions—A Lost Chance in the Fight against HIV and Other Virus Diseases? Antivir. Chem. Chemother. 2000, 11, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Gammeltoft, K.A.; Galli, A.; Offersgaard, A.; Fahnøe, U.; Ramirez, S.; Bukh, J.; Gottwein, J.M. Efficacy of Ion-Channel Inhibitors Amantadine, Memantine and Rimantadine for the Treatment of SARS-CoV-2 In Vitro. Viruses 2021, 13, 2082. [Google Scholar] [CrossRef]
- Al-Badr, A.A.; Ajarim, T.D.S. Ganciclovir. In Profiles of Drug Substances, Excipients and Related Methodology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 43, pp. 1–208. ISBN 978-0-12-815125-9. [Google Scholar]
- Kłysik, K.; Pietraszek, A.; Karewicz, A.; Nowakowska, M. Acyclovir in the Treatment of Herpes Viruses—A Review. Curr. Med. Chem. 2020, 27, 4118–4137. [Google Scholar] [CrossRef]
- Li, G.; Wang, Y.; De Clercq, E. Approved HIV Reverse Transcriptase Inhibitors in the Past Decade. Acta Pharm. Sin. B 2021, S2211383521004433. [Google Scholar] [CrossRef]
- Furman, P.A.; Barry, D.W. Spectrum of Antiviral Activity and Mechanism of Action of Zidovudine. An Overview. Am. J. Med. 1988, 85, 176–181. [Google Scholar] [PubMed]
- Havens, J.P.; Podany, A.T.; Scarsi, K.K.; Fletcher, C.V. Clinical Pharmacokinetics and Pharmacodynamics of Etravirine: An Updated Review. Clin. Pharmacokinet. 2020, 59, 137–154. [Google Scholar] [CrossRef] [PubMed]
- Bardsley-Elliot, A.; Perry, C.M. Nevirapine: A Review of Its Use in the Prevention and Treatment of Paediatric HIV Infection. Paediatr. Drugs 2000, 2, 373–407. [Google Scholar] [CrossRef]
- De Clercq, E. The Role of Non-Nucleoside Reverse Transcriptase Inhibitors (NNRTIs) in the Therapy of HIV-1 Infection. Antivir. Res. 1998, 38, 153–179. [Google Scholar] [CrossRef]
- Chemaly, R.F.; Ullmann, A.J.; Stoelben, S.; Richard, M.P.; Bornhäuser, M.; Groth, C.; Einsele, H.; Silverman, M.; Mullane, K.M.; Brown, J.; et al. Letermovir for Cytomegalovirus Prophylaxis in Hematopoietic-Cell Transplantation. N. Engl. J. Med. 2014, 370, 1781–1789. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.; Schiffer, C.; Lee, S.-K.; Swanstrom, R. Viral Protease Inhibitors. In Antiviral Strategies; Kräusslich, H.-G., Bartenschlager, R., Eds.; Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2009; Volume 189, pp. 85–110. ISBN 978-3-540-79085-3. [Google Scholar]
- Flexner, C. HIV-Protease Inhibitors. N. Engl. J. Med. 1998, 338, 1281–1293. [Google Scholar] [CrossRef]
- Varghese, J.N.; McKimm-Breschkin, J.L.; Caldwell, J.B.; Kortt, A.A.; Colman, P.M. The Structure of the Complex between Influenza Virus Neuraminidase and Sialic Acid, the Viral Receptor. Proteins Struct. Funct. Genet. 1992, 14, 327–332. [Google Scholar] [CrossRef]
- Moscona, A. Neuraminidase Inhibitors for Influenza. N. Engl. J. Med. 2005, 353, 1363–1373. [Google Scholar] [CrossRef] [Green Version]
- De Clercq, E. Clinical Potential of the Acyclic Nucleoside Phosphonates Cidofovir, Adefovir, and Tenofovir in Treatment of DNA Virus and Retrovirus Infections. Clin. Microbiol. Rev. 2003, 16, 569–596. [Google Scholar] [CrossRef] [Green Version]
- Maga, G.; Radi, M.; Gerard, M.-A.; Botta, M.; Ennifar, E. HIV-1 RT Inhibitors with a Novel Mechanism of Action: NNRTIs That Compete with the Nucleotide Substrate. Viruses 2010, 2, 880–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eggleton, J.S.; Nagalli, S. Highly Active Antiretroviral Therapy (HAART); StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Yu, Y.; Shen, M.; Song, Q.; Xie, J. Biological Activities and Pharmaceutical Applications of Polysaccharide from Natural Resources: A Review. Carbohydr. Polym. 2018, 183, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Chen, X.; Gong, P. Classification, Structure and Mechanism of Antiviral Polysaccharides Derived from Edible and Medicinal Fungus. Int. J. Biol. Macromol. 2021, 183, 1753–1773. [Google Scholar] [CrossRef]
- Wang, W.; Wang, S.-X.; Guan, H.-S. The Antiviral Activities and Mechanisms of Marine Polysaccharides: An Overview. Mar. Drugs 2012, 10, 2795–2816. [Google Scholar] [CrossRef]
- Witvrouw, M.; De Clercq, E. Sulfated Polysaccharides Extracted from Sea Algae as Potential Antiviral Drugs. Gen. Pharmacol. Vasc. Syst. 1997, 29, 497–511. [Google Scholar] [CrossRef]
- Halpern, V.; Ogunsola, F.; Obunge, O.; Wang, C.-H.; Onyejepu, N.; Oduyebo, O.; Taylor, D.; McNeil, L.; Mehta, N.; Umo-Otong, J.; et al. Effectiveness of Cellulose Sulfate Vaginal Gel for the Prevention of HIV Infection: Results of a Phase III Trial in Nigeria. PLoS ONE 2008, 3, e3784. [Google Scholar] [CrossRef] [Green Version]
- Kwon, P.S.; Oh, H.; Kwon, S.-J.; Jin, W.; Zhang, F.; Fraser, K.; Hong, J.J.; Linhardt, R.J.; Dordick, J.S. Sulfated Polysaccharides Effectively Inhibit SARS-CoV-2 in Vitro. Cell Discov. 2020, 6, 50. [Google Scholar] [CrossRef]
- Neyts, J.; Snoeck, R.; Schols, D.; Balzarini, J.; Esko, J.D.; Van Schepdael, A.; De Clercq, E. Sulfated Polymers Inhibit the Interaction of Human Cytomegalovirus with Cell Surface Heparan Sulfate. Virology 1992, 189, 48–58. [Google Scholar] [CrossRef]
- Tandon, R.; Sharp, J.S.; Zhang, F.; Pomin, V.H.; Ashpole, N.M.; Mitra, D.; McCandless, M.G.; Jin, W.; Liu, H.; Sharma, P.; et al. Effective Inhibition of SARS-CoV-2 Entry by Heparin and Enoxaparin Derivatives. J. Virol. 2021, 95. [Google Scholar] [CrossRef]
- Jinno-Oue, A.; Tanaka, A.; Shimizu, N.; Mori, T.; Sugiura, N.; Kimata, K.; Isomura, H.; Hoshino, H. Inhibitory Effect of Chondroitin Sulfate Type E on the Binding Step of Human T-Cell Leukemia Virus Type 1. AIDS Res. Hum. Retrovir. 2013, 29, 621–629. [Google Scholar] [CrossRef]
- Song, S.; Peng, H.; Wang, Q.; Liu, Z.; Dong, X.; Wen, C.; Ai, C.; Zhang, Y.; Wang, Z.; Zhu, B. Inhibitory Activities of Marine Sulfated Polysaccharides against SARS-CoV-2. Food Funct. 2020, 11, 7415–7420. [Google Scholar] [CrossRef] [PubMed]
- Bergefall, K.; Trybala, E.; Johansson, M.; Uyama, T.; Naito, S.; Yamada, S.; Kitagawa, H.; Sugahara, K.; Bergström, T. Chondroitin Sulfate Characterized by the E-Disaccharide Unit Is a Potent Inhibitor of Herpes Simplex Virus Infectivity and Provides the Virus Binding Sites on Gro2C Cells. J. Biol. Chem. 2005, 280, 32193–32199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, D.; Era, S.; Watanabe, I.; Arihara, M.; Sugiura, N.; Kimata, K.; Suzuki, Y.; Morita, K.; Hidari, K.I.P.J.; Suzuki, T. Antiviral Activity of Chondroitin Sulphate E Targeting Dengue Virus Envelope Protein. Antivir. Res. 2010, 88, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Kulikov, S.N.; Chirkov, S.N.; Il’ina, A.V.; Lopatin, S.A.; Varlamov, V.P. Effect of the molecular weight of chitosan on its antiviral activity in plants. Prikl. Biokhim. Mikrobiol. 2006, 42, 224–228. [Google Scholar] [CrossRef]
- Safarzadeh, M.; Sadeghi, S.; Azizi, M.; Rastegari-Pouyani, M.; Pouriran, R.; Haji Molla Hoseini, M. Chitin and Chitosan as Tools to Combat COVID-19: A Triple Approach. Int. J. Biol. Macromol. 2021, 183, 235–244. [Google Scholar] [CrossRef]
- Su, J.; Jiang, L.; Wu, J.; Liu, Z.; Wu, Y. Anti-Tumor and Anti-Virus Activity of Polysaccharides Extracted from Sipunculus Nudus (SNP) on Hepg2.2.15. Int. J. Biol. Macromol. 2016, 87, 597–602. [Google Scholar] [CrossRef]
- Woo, E.-R.; Kim, W.S.; Kim, Y.S. Virus-Cell Fusion Inhibitory Activity for the Polysaccharides from Various Korean Edible Clams. Arch. Pharm. Res. 2001, 24, 514–517. [Google Scholar] [CrossRef]
- Queiroz, K.C.S.; Medeiros, V.P.; Queiroz, L.S.; Abreu, L.R.D.; Rocha, H.A.O.; Ferreira, C.V.; Jucá, M.B.; Aoyama, H.; Leite, E.L. Inhibition of Reverse Transcriptase Activity of HIV by Polysaccharides of Brown Algae. Biomed. Pharmacother. 2008, 62, 303–307. [Google Scholar] [CrossRef]
- Mandal, P.; Mateu, C.G.; Chattopadhyay, K.; Pujol, C.A.; Damonte, E.B.; Ray, B. Structural Features and Antiviral Activity of Sulphated Fucans from the Brown Seaweed Cystoseira indica. Antivir. Chem. Chemother. 2007, 18, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, U.; Mateu, C.G.; Chattopadhyay, K.; Pujol, C.A.; Damonte, E.B.; Ray, B. Structure and Antiviral Activity of Sulfated Fucans from Stoechospermum marginatum. Phytochemistry 2006, 67, 2474–2482. [Google Scholar] [CrossRef]
- Ahmadi, A.; Zorofchian Moghadamtousi, S.; Abubakar, S.; Zandi, K. Antiviral Potential of Algae Polysaccharides Isolated from Marine Sources: A Review. BioMed Res. Int. 2015, 2015, 825203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sano, Y. Antiviral Activity of Alginate against Infection by Tobacco Mosaic Virus. Carbohydr. Polym. 1999, 38, 183–186. [Google Scholar] [CrossRef]
- Hui, B.; Xia, W.; Li, J.; Wang, L.; Ai, J.; Geng, M. Sulfated Polymannuroguluronate, a Novel Anti-Acquired Immune Deficiency Syndrome Drug Candidate, Blocks Neuroinflammatory Signalling by Targeting the Transactivator of Transcription (Tat) Protein: Characterization of the Binding Mode of SPMG and Tat. J. Neurochem. 2006, 97, 334–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buck, C.B.; Thompson, C.D.; Roberts, J.N.; Müller, M.; Lowy, D.R.; Schiller, J.T. Carrageenan Is a Potent Inhibitor of Papillomavirus Infection. PLoS Pathog. 2006, 2, e69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talarico, L.B.; Damonte, E.B. Interference in Dengue Virus Adsorption and Uncoating by Carrageenans. Virology 2007, 363, 473–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frediansyah, A. The Antiviral Activity of Iota-, Kappa-, and Lambda-Carrageenan against COVID-19: A Critical Review. Clin. Epidemiol. Glob. Health 2021, 12, 100826. [Google Scholar] [CrossRef]
- Morokutti-Kurz, M.; Fröba, M.; Graf, P.; Große, M.; Grassauer, A.; Auth, J.; Schubert, U.; Prieschl-Grassauer, E. Iota-Carrageenan Neutralizes SARS-CoV-2 and Inhibits Viral Replication in Vitro. PLoS ONE 2021, 16, e0237480. [Google Scholar] [CrossRef] [PubMed]
- Chahla, R.E.; Ruiz, L.M.; Ortega, E.S.; Morales, M.F.; Barreiro, F.; George, A.; Mansilla, C.; D’Amato, S.P.; Barrenechea, G.; Goroso, G.D. Intensive Treatment with Ivermectin and Iota-Carrageenan as Pre-Exposure Prophylaxis for COVID-19 in Health Care Workers from Tucuman, Argentina. Am. J. Ther. 2021, 28, e601–e604. [Google Scholar] [CrossRef]
- Stathis, C.; Victoria, N.; Loomis, K.; Nguyen, S.A.; Eggers, M.; Septimus, E.; Safdar, N. Review of the Use of Nasal and Oral Antiseptics during a Global Pandemic. Future Microbiol. 2021, 16, 119–130. [Google Scholar] [CrossRef]
- Schütz, D.; Conzelmann, C.; Fois, G.; Groß, R.; Weil, T.; Wettstein, L.; Stenger, S.; Zelikin, A.; Hoffmann, T.K.; Frick, M.; et al. Carrageenan-Containing over-the-Counter Nasal and Oral Sprays Inhibit SARS-CoV-2 Infection of Airway Epithelial Cultures. Am. J. Physiol. Lung Cell. Mol. Physiol. 2021, 320, L750–L756. [Google Scholar] [CrossRef]
- Carlucci, M.; Scolaro, L.; Noseda, M.; Cerezo, A.; Damonte, E. Protective Effect of a Natural Carrageenan on Genital Herpes Simplex Virus Infection in Mice. Antivir. Res. 2004, 64, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Nagorskaia, V.P.; Reunov, A.V.; Lapshina, L.A.; Ermak, I.M.; Barabanova, A.O. Influence of kappa/beta-carrageenan from red alga Tichocarpus crinitus on development of local infection induced by tobacco mosaic virus in Xanthi-nc tobacco leaves. Izv. Akad. Nauk. Ser. Biol. 2008, 3, 360–364. [Google Scholar]
- Ponce, M.; Zuasti, E.; Anguís, V.; Fernández-Díaz, C. Effects of the Sulfated Polysaccharide Ulvan from Ulva Ohnoi on the Modulation of the Immune Response in Senegalese Sole (Solea senegalensis). Fish Shellfish. Immunol. 2020, 100, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Shefer, S.; Robin, A.; Chemodanov, A.; Lebendiker, M.; Bostwick, R.; Rasmussen, L.; Lishner, M.; Gozin, M.; Golberg, A. Fighting SARS-CoV-2 with Green Seaweed Ulva Sp. Extract: Extraction Protocol Predetermines Crude Ulvan Extract Anti-SARS-CoV-2 Inhibition Properties in in Vitro Vero-E6 Cells Assay. PeerJ 2021, 9, e12398. [Google Scholar] [CrossRef]
- Chi, Y.; Zhang, M.; Wang, X.; Fu, X.; Guan, H.; Wang, P. Ulvan Lyase Assisted Structural Characterization of Ulvan from Ulva pertusa and Its Antiviral Activity against Vesicular Stomatitis Virus. Int. J. Biol. Macromol. 2020, 157, 75–82. [Google Scholar] [CrossRef]
- Hans, N.; Malik, A.; Naik, S. Antiviral Activity of Sulfated Polysaccharides from Marine Algae and Its Application in Combating COVID-19: Mini Review. Bioresour. Technol. Rep. 2021, 13, 100623. [Google Scholar] [CrossRef]
- Chiu, Y.-H.; Chan, Y.-L.; Li, T.-L.; Wu, C.-J. Inhibition of Japanese Encephalitis Virus Infection by the Sulfated Polysaccharide Extracts from Ulva lactuca. Mar. Biotechnol. 2012, 14, 468–478. [Google Scholar] [CrossRef]
- Lee, J.-B.; Hayashi, K.; Hirata, M.; Kuroda, E.; Suzuki, E.; Kubo, Y.; Hayashi, T. Antiviral Sulfated Polysaccharide from Navicula Directa, a Diatom Collected from Deep-Sea Water in Toyama Bay. Biol. Pharm. Bull. 2006, 29, 2135–2139. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Yim, J.H.; Kim, S.-Y.; Kim, H.S.; Lee, W.G.; Kim, S.J.; Kang, P.-S.; Lee, C.-K. In Vitro Inhibition of Influenza a Virus Infection by Marine Microalga-Derived Sulfated Polysaccharide p-KG03. Antivir. Res. 2012, 93, 253–259. [Google Scholar] [CrossRef]
- Hasui, M.; Matsuda, M.; Okutani, K.; Shigeta, S. In Vitro Antiviral Activities of Sulfated Polysaccharides from a Marine Microalga (Cochlodinium polykrikoides) against Human Immunodeficiency Virus and Other Enveloped Viruses. Int. J. Biol. Macromol. 1995, 17, 293–297. [Google Scholar] [CrossRef]
- Yim, J.H.; Kim, S.J.; Ahn, S.H.; Lee, C.K.; Rhie, K.T.; Lee, H.K. Antiviral Effects of Sulfated Exopolysaccharide from the Marine Microalga Gyrodinium impudicum Strain KG03. Mar. Biotechnol. 2004, 6, 17–25. [Google Scholar] [CrossRef]
- Jabeen, M.; Dutot, M.; Fagon, R.; Verrier, B.; Monge, C. Seaweed Sulfated Polysaccharides against Respiratory Viral Infections. Pharmaceutics 2021, 13, 733. [Google Scholar] [CrossRef]
- Song, X.; Yin, Z.; Li, L.; Cheng, A.; Jia, R.; Xu, J.; Wang, Y.; Yao, X.; Lv, C.; Zhao, X. Antiviral Activity of Sulfated Chuanminshen violaceum Polysaccharide against Duck Enteritis Virus in Vitro. Antivir. Res. 2013, 98, 344–351. [Google Scholar] [CrossRef]
- Ray, B.; Ali, I.; Jana, S.; Mukherjee, S.; Pal, S.; Ray, S.; Schütz, M.; Marschall, M. Antiviral Strategies Using Natural Source-Derived Sulfated Polysaccharides in the Light of the COVID-19 Pandemic and Major Human Pathogenic Viruses. Viruses 2021, 14, 35. [Google Scholar] [CrossRef]
- Lee, J.-B.; Tanikawa, T.; Hayashi, K.; Asagi, M.; Kasahara, Y.; Hayashi, T. Characterization and Biological Effects of Two Polysaccharides Isolated from Acanthopanax sciadophylloides. Carbohydr. Polym. 2015, 116, 159–166. [Google Scholar] [CrossRef]
- Baek, S.-H.; Lee, J.G.; Park, S.Y.; Bae, O.N.; Kim, D.-H.; Park, J.H. Pectic Polysaccharides from Panax ginseng as the Antirotavirus principals in Ginseng. Biomacromolecules 2010, 11, 2044–2052. [Google Scholar] [CrossRef]
- Wang, C.R.; Ng, T.B.; Li, L.; Fang, J.C.; Jiang, Y.; Wen, T.Y.; Qiao, W.T.; Li, N.; Liu, F. Isolation of a Polysaccharide with Antiproliferative, Hypoglycemic, Antioxidant and HIV-1 Reverse Transcriptase Inhibitory Activities from the Fruiting Bodies of the Abalone Mushroom Pleurotus abalonus. J. Pharm. Pharmacol. 2011, 63, 825–832. [Google Scholar] [CrossRef]
- Ren, G.; Xu, L.; Lu, T.; Yin, J. Structural Characterization and Antiviral Activity of Lentinan from Lentinus Edodes Mycelia against Infectious Hematopoietic Necrosis Virus. Int. J. Biol. Macromol. 2018, 115, 1202–1210. [Google Scholar] [CrossRef]
- Ohta, Y.; Lee, J.-B.; Hayashi, K.; Fujita, A.; Park, D.K.; Hayashi, T. In Vivo Anti-Influenza Virus Activity of an Immunomodulatory Acidic Polysaccharide Isolated from Cordyceps militaris Grown on Germinated Soybeans. J. Agric. Food Chem. 2007, 55, 10194–10199. [Google Scholar] [CrossRef]
- Martinez, A.F.C.; Moraes, L.A.B. Liquid Chromatography-Tandem Mass Spectrometry Characterization of Five New Leucinostatins Produced by Paecilomyces lilacinus CG—189. J. Antibiot. 2015, 68, 178–184. [Google Scholar] [CrossRef]
- Mader, J.; Gallo, A.; Schommartz, T.; Handke, W.; Nagel, C.-H.; Günther, P.; Brune, W.; Reich, K. Calcium Spirulan Derived from Spirulina platensis Inhibits Herpes Simplex Virus 1 Attachment to Human Keratinocytes and Protects against Herpes Labialis. J. Allergy Clin. Immunol. 2016, 137, 197–203.e3. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, T.; Hayashi, K.; Maeda, M.; Kojima, I. Calcium Spirulan, an Inhibitor of Enveloped Virus Replication, from a Blue-Green Alga Spirulina platensis. J. Nat. Prod. 1996, 59, 83–87. [Google Scholar] [CrossRef]
- Kanekiyo, K.; Hayashi, K.; Takenaka, H.; Lee, J.-B.; Hayashi, T. Anti-Herpes Simplex Virus Target of an Acidic Polysaccharide, Nostoflan, from the Edible Blue-Green Alga Nostoc flagelliforme. Biol. Pharm. Bull. 2007, 30, 1573–1575. [Google Scholar] [CrossRef] [Green Version]
- Kanekiyo, K.; Lee, J.-B.; Hayashi, K.; Takenaka, H.; Hayakawa, Y.; Endo, S.; Hayashi, T. Isolation of an Antiviral Polysaccharide, Nostoflan, from a Terrestrial Cyanobacterium, Nostocf lagelliforme. J. Nat. Prod. 2005, 68, 1037–1041. [Google Scholar] [CrossRef]
- Rider, C.C.; Coombe, D.R.; Harrop, H.A.; Hounsell, E.F.; Bauer, C.; Feeney, J.; Mulloy, B.; Mahmood, N.; Hay, A.; Parish, C.R. Anti-HIV-1 Activity of Chemically Modified Heparins: Correlation between Binding to the V3 Loop of Gp120 and Inhibition of Cellular HIV-1 Infection in Vitro. Biochemistry 1994, 33, 6974–6980. [Google Scholar] [CrossRef]
- Kim, S.Y.; Jin, W.; Sood, A.; Montgomery, D.W.; Grant, O.C.; Fuster, M.M.; Fu, L.; Dordick, J.S.; Woods, R.J.; Zhang, F.; et al. Characterization of Heparin and Severe Acute Respiratory Syndrome-Related Coronavirus 2 (SARS-CoV-2) Spike Glycoprotein Binding Interactions. Antivir. Res. 2020, 181, 104873. [Google Scholar] [CrossRef]
- Fenice, M.; Gorrasi, S. Advances in Chitin and Chitosan Science. Molecules 2021, 26, 1805. [Google Scholar] [CrossRef]
- De Souza, A.F.; Galindo, H.M.; de Lima, M.A.B.; Ribeaux, D.R.; Rodríguez, D.M.; da Silva Andrade, R.F.; Gusmão, N.B.; de Campos-Takaki, G.M. Biotechnological Strategies for Chitosan Production by Mucoralean Strains and Dimorphism Using Renewable Substrates. Int. J. Mol. Sci. 2020, 21, 4286. [Google Scholar] [CrossRef]
- Mania, S.; Partyka, K.; Pilch, J.; Augustin, E.; Cieślik, M.; Ryl, J.; Jinn, J.-R.; Wang, Y.-J.; Michałowska, A.; Tylingo, R. Obtaining and Characterization of the PLA/Chitosan Foams with Antimicrobial Properties Achieved by the Emulsification Combined with the Dissolution of Chitosan by CO2 Saturation. Molecules 2019, 24, 4532. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.; Zhou, L.; Lin, L.; Cai, Y.; Sun, H.; Zhao, L.; Gao, N.; Yin, R.; Zhao, J. Physicochemical Characteristics and Anticoagulant Activities of the Polysaccharides from Sea Cucumber Pattalus mollis. Mar. Drugs 2019, 17, 198. [Google Scholar] [CrossRef] [Green Version]
- Thammakarn, C.; Tsujimura, M.; Satoh, K.; Hasegawa, T.; Tamura, M.; Kawamura, A.; Ishida, Y.; Suguro, A.; Hakim, H.; Ruenphet, S.; et al. Efficacy of Scallop Shell Powders and Slaked Lime for Inactivating Avian Influenza Virus under Harsh Conditions. Arch. Virol. 2015, 160, 2577–2581. [Google Scholar] [CrossRef]
- Tang, F.; Huang, G.; Lin, L.; Yin, H.; Shao, L.; Xu, R.; Cui, X. Anti-HBV Activities of Polysaccharides from Thais clavigera (Küster) by In Vitro and In Vivo Study. Mar. Drugs 2021, 19, 195. [Google Scholar] [CrossRef]
- Lee, J.-C.; Hou, M.-F.; Huang, H.-W.; Chang, F.-R.; Yeh, C.-C.; Tang, J.-Y.; Chang, H.-W. Marine Algal Natural Products with Anti-Oxidative, Anti-Inflammatory, and Anti-Cancer Properties. Cancer Cell Int. 2013, 13, 55. [Google Scholar] [CrossRef] [Green Version]
- Mabeau, S.; Kloareg, B. Isolation and Analysis of the Cell Walls of Brown Algae: Fucus spiralis, F. Ceranoides, F. Vesiculosus, F. Serratus, Bifurcaria bifurcata and Laminaria digitata. J. Exp. Bot. 1987, 38, 1573–1580. [Google Scholar] [CrossRef]
- Xin, X.L.; Geng, M.Y.; Guan, H.S.; Li, Z.L. Study on the Mechanism of Inhibitory Action of 911 on Replication of HIV-1 in Vitro. Chin. J. Mar. Drugs 2000, 19, 15–18. [Google Scholar]
- Muto, S.; Niimura, K.; Oohara, M.; Oguchi, Y.; Matsunaga, K.; Hirose, K.; Kakuchi, J.; Sugita, N.; Furusho, T.; Yoshikumi, C.; et al. Polysaccharides and Antiviral Drugs Containing the Same as Active Ingredient. U.S. Patent 5,089,481, 18 February 1992. [Google Scholar]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and Bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef] [Green Version]
- Krylova, N.V.; Ermakova, S.P.; Lavrov, V.F.; Leneva, I.A.; Kompanets, G.G.; Iunikhina, O.V.; Nosik, M.N.; Ebralidze, L.K.; Falynskova, I.N.; Silchenko, A.S.; et al. The Comparative Analysis of Antiviral Activity of Native and Modified Fucoidans from Brown Algae Fucus evanescens In Vitro and In Vivo. Mar. Drugs 2020, 18, 224. [Google Scholar] [CrossRef] [Green Version]
- Lahaye, M.; Robic, A. Structure and Functional Properties of Ulvan, a Polysaccharide from Green Seaweeds. Biomacromolecules 2007, 8, 1765–1774. [Google Scholar] [CrossRef]
- Yildiz, H.; Karatas, N. Microbial Exopolysaccharides: Resources and Bioactive Properties. Process Biochem. 2018, 72, 41–46. [Google Scholar] [CrossRef]
- Jang, Y.; Shin, H.; Lee, M.K.; Kwon, O.S.; Shin, J.S.; Kim, Y.; Kim, C.W.; Lee, H.-R.; Kim, M. Antiviral Activity of Lambda-Carrageenan against Influenza Viruses and Severe Acute Respiratory Syndrome Coronavirus 2. Sci. Rep. 2021, 11, 821. [Google Scholar] [CrossRef]
- Feng, S.; Ding, H.; Liu, L.; Peng, C.; Huang, Y.; Zhong, F.; Li, W.; Meng, T.; Li, J.; Wang, X.; et al. Astragalus Polysaccharide Enhances the Immune Function of RAW264.7 Macrophages via the NF-κB P65/MAPK Signaling Pathway. Exp. Ther. Med. 2020, 21, 20. [Google Scholar] [CrossRef]
- Vijayakumar, P.; Dhas, T.S.; Mani, R.; Raguraman, V. Antiviral Activity of Sulfated Polysaccharides from Sargassum ilicifolium against Fish Betanodavirus Infection. Aquac. Int. 2021, 29, 1049–1067. [Google Scholar] [CrossRef]
- Liang, Y.; Li, K.; Niu, F.; Li, Y.; Wei, H.; Dai, Y.; Wang, Y.; Zhou, C.; Wan, X. Salvia Plebeia, R. Br. Polysaccharides (SPP) against RSV (Respiratory Syncytial Virus) Infection: Antiviral Effect and Mechanisms of Action. Biomed. Pharmacother. 2021, 141, 111843. [Google Scholar] [CrossRef]
- Yang, J.; Fu, Y.-P.; Du, B.-X.; Yang, Y.; Rong, R. Protective effect of Asarum polysaccharide on H1N1 influenza virus infection and expression of inflammatory factors. Zhongguo Zhong Yao Za Zhi Zhongguo Zhongyao Zazhi China J. Chin. Mater. Medica 2021, 46, 412–419. [Google Scholar] [CrossRef]
- Wang, T.; Wang, X.; Zhuo, Y.; Si, C.; Yang, L.; Meng, L.; Zhu, B. Antiviral Activity of a Polysaccharide from Radix Isatidis (Isatis 998 Indigotica Fortune) against Hepatitis B Virus (HBV) in Vitro via Activation of JAK/STAT Signal Pathway. J. Ethnopharmacol. 2020, 15, 112782. [Google Scholar] [CrossRef]
- Ilyicheva, T.N.; Teplyakova, T.V.; Svyatchenko, S.V.; Asbaganov, S.V.; Zmitrovich, I.V.; Vlasenko, A.V. Antiviral Activity of Total 1001 Polysaccharide Fraction of Water and Ethanol Extracts of Pleurotus pulmonarius against the Influenza A Virus. J. Fungal Biol. 2020, 10, 224–235. [Google Scholar]
- Arena, A.; Gugliandolo, C.; Stassi, G.; Pavone, B.; Iannello, D.; Bisignano, G.; Maugeri, T.L. An Exopolysaccharide Produced by Geobacillus thermodenitrificans Strain B3-72: Antiviral Activity on Immunocompetent Cells. Immunol. Lett. 2009, 123, 132–137. [Google Scholar] [CrossRef]
- Lee, J.-B.; Hayashi, T.; Hayashi, K.; Sankawa, U. Structural Analysis of Calcium Spirulan (Ca−SP)-Derived Oligosaccharides Using Electrospray Ionization Mass Spectrometry. J. Nat. Prod. 2000, 63, 136–138. [Google Scholar] [CrossRef]
- Rechter, S.; König, T.; Auerochs, S.; Thulke, S.; Walter, H.; Dörnenburg, H.; Walter, C.; Marschall, M. Antiviral Activity of Arthrospira-Derived Spirulan-like Substances. Antivir. Res. 2006, 72, 197–206. [Google Scholar] [CrossRef]
- Liu, Y.; Su, P.; Xu, J.; Chen, S.; Zhang, J.; Zhou, S.; Wang, Y.; Tang, Q.; Wang, Y. Structural Characterization of a Bioactive Water-Soluble Heteropolysaccharide from Nostoc Sphaeroids Kütz. Carbohydr. Polym. 2018, 200, 552–559. [Google Scholar] [CrossRef]
- Callahan, L.N.; Phelan, M.; Mallinson, M.; Norcross, M.A. Dextran Sulfate Blocks Antibody Binding to the Principal Neutralizing Domain of Human Immunodeficiency Virus Type 1 without Interfering with Gp120-CD4 Interactions. J. Virol. 1991, 65, 1543–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neyts, J.; Reymen, D.; Letourneur, D.; Jozefonvicz, J.; Schols, D.; Este, J.; Andrei, G.; McKenna, P.; Witvrouw, M.; Ikeda, S.; et al. Differential Antiviral Activity of Derivatized Dextrans. Biochem. Pharmacol. 1995, 50, 743–751. [Google Scholar] [CrossRef]
- Nácher-Vázquez, M.; Ballesteros, N.; Canales, Á.; Rodríguez Saint-Jean, S.; Pérez-Prieto, S.I.; Prieto, A.; Aznar, R.; López, P. Dextrans Produced by Lactic Acid Bacteria Exhibit Antiviral and Immunomodulatory Activity against Salmonid Viruses. Carbohydr. Polym. 2015, 124, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Copeland, R.; Balasubramaniam, A.; Tiwari, V.; Zhang, F.; Bridges, A.; Linhardt, R.J.; Shukla, D.; Liu, J. Using a 3-O-Sulfated Heparin Octasaccharide to Inhibit the Entry of Herpes Simplex Virus Type 1. Biochemistry 2008, 47, 5774–5783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Sousa Cardozo, F.T.G.; Camelini, C.M.; Mascarello, A.; Rossi, M.J.; Nunes, R.J.; Barardi, C.R.M.; de Mendonça, M.M.; Simões, C.M.O. Antiherpetic Activity of a Sulfated Polysaccharide from Agaricus brasiliensis Mycelia. Antivir. Res. 2011, 92, 108–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, Y.; Pei, Y.; Qu, C.; Lai, Z.; Ren, Z.; Yang, K.; Xiong, S.; Zhang, Y.; Yang, C.; Wang, D.; et al. In Vitro Anti-Herpes Simplex Virus Activity of 1,2,4,6-Tetra-O-Galloyl-β- d -Glucose from Phyllanthus emblica L. (Euphorbiaceae): In vitro anti-herpes simplex virus activity of 1246TGG from P. emblica. Phytother. Res. 2011, 25, 975–982. [Google Scholar] [CrossRef]
- Nguyen, T.L.; Chen, J.; Hu, Y.; Wang, D.; Fan, Y.; Wang, J.; Abula, S.; Zhang, J.; Qin, T.; Chen, X.; et al. In Vitro Antiviral Activity of Sulfated Auricularia auricula Polysaccharides. Carbohydr. Polym. 2012, 90, 1254–1258. [Google Scholar] [CrossRef]
- Zhao, X.; Hu, Y.; Wang, D.; Guo, L.; Yang, S.; Fan, Y.; Zhao, B.; Wang, Y.; Abula, S. Optimization of Sulfated Modification Conditions of Tremella Polysaccharide and Effects of Modifiers on Cellular Infectivity of NDV. Int. J. Biol. Macromol. 2011, 49, 44–49. [Google Scholar] [CrossRef]
- Lopes, N.; Faccin-Galhardi, L.C.; Espada, S.F.; Pacheco, A.C.; Ricardo, N.M.P.S.; Linhares, R.E.C.; Nozawa, C. Sulfated Polysaccharide of Caesalpinia ferrea Inhibits Herpes Simplex Virus and Poliovirus. Int. J. Biol. Macromol. 2013, 60, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Zhang, Y.; Yin, Z.; Zhao, X.; Liang, X.; He, C.; Yin, L.; Lv, C.; Zhao, L.; Ye, G.; et al. Antiviral Effect of Sulfated Chuanmingshen violaceum Polysaccharide in Chickens Infected with Virulent Newcastle Disease Virus. Virology 2015, 476, 316–322. [Google Scholar] [CrossRef] [Green Version]
- Mandal, P.; Pujol, C.A.; Carlucci, M.J.; Chattopadhyay, K.; Damonte, E.B.; Ray, B. Anti-Herpetic Activity of a Sulfated Xylomannan from Scinaia hatei. Phytochemistry 2008, 69, 2193–2199. [Google Scholar] [CrossRef] [PubMed]
- Sosa, M.A.G.; Fazely, F.; Koch, J.A.; Vercellotti, S.V.; Ruprecht, R.M. N-Carboxymethylchitosan-N,O-Sulfate as an Anti-HIV-1 Agent. Biochem. Biophys. Res. Commun. 1991, 174, 489–496. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, P.; Yu, G.-L.; Li, C.-X.; Hao, C.; Qi, X.; Zhang, L.-J.; Guan, H.-S. Preparation and Anti-Influenza A Virus Activity of κ-Carrageenan Oligosaccharide and Its Sulphated Derivatives. Food Chem. 2012, 133, 880–888. [Google Scholar] [CrossRef]
- Ming, K.; Chen, Y.; Yao, F.; Shi, J.; Yang, J.; Du, H.; Wang, X.; Wang, Y.; Liu, J. Phosphorylated Codonopsis pilosula Polysaccharide Could Inhibit the Virulence of Duck Hepatitis A Virus Compared with Codonopsis pilosula Polysaccharide. Int. J. Biol. Macromol. 2017, 94, 28–35. [Google Scholar] [CrossRef]
- Ming, K.; Chen, Y.; Shi, J.; Yang, J.; Yao, F.; Du, H.; Zhang, W.; Bai, J.; Liu, J.; Wang, D.; et al. Effects of Chrysanthemum indicum Polysaccharide and Its Phosphate on Anti-Duck Hepatitis a Virus and Alleviating Hepatic Injury. Int. J. Biol. Macromol. 2017, 102, 813–821. [Google Scholar] [CrossRef]
- Feng, H.; Fan, J.; Yang, S.; Zhao, X.; Yi, X. Antiviral Activity of Phosphorylated Radix Cyathulae officinalis Polysaccharide against Canine Parvovirus in Vitro. Int. J. Biol. Macromol. 2017, 99, 511–518. [Google Scholar] [CrossRef]
- Wu, D.; Ensinas, A.; Verrier, B.; Primard, C.; Cuvillier, A.; Champier, G.; Paul, S.; Delair, T. Zinc-Stabilized Chitosan-Chondroitin Sulfate Nanocomplexes for HIV-1 Infection Inhibition Application. Mol. Pharm. 2016, 13, 3279–3291. [Google Scholar] [CrossRef]
- Artan, M.; Karadeniz, F.; Karagozlu, M.Z.; Kim, M.-M.; Kim, S.-K. Anti-HIV-1 Activity of Low Molecular Weight Sulfated Chitooligosaccharides. Carbohydr. Res. 2010, 345, 656–662. [Google Scholar] [CrossRef]
- Yamada, T.; Ogamo, A.; Saito, T.; Uchiyama, H.; Nakagawa, Y. Preparation of O-Acylated Low-Molecular-Weight Carrageenans with Potent Anti-HIV Activity and Low Anticoagulant Effect. Carbohydr. Polym. 2000, 41, 115–120. [Google Scholar] [CrossRef]
- Artan, M.; Karadeniz, F.; Kim, M.M.; Kim, S.K. Chitosan Derivatives as HIV-1 Inhibitors. J. Biotechnol. 2008, 136S, S527–S540. [Google Scholar] [CrossRef]
- Roskoski, R.; Lim, C.T.; Roskoski, L.M. Human Brain and Placental Choline Acetyltransferase: Purification and Properties. Biochemistry 1975, 14, 5105–5110. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-S.; Kim, Y.-M.; Lee, M.-S.; Ahn, C.-B.; Jung, W.-K.; Je, J.-Y. Synergistic Effects between Aminoethyl-Chitosans and β-Lactams against Methicillin-Resistant Staphylococcus aureus (MRSA). Bioorg. Med. Chem. Lett. 2010, 20, 975–978. [Google Scholar] [CrossRef] [PubMed]
- Liaqat, F.; Eltem, R. Chitooligosaccharides and Their Biological Activities: A Comprehensive Review. Carbohydr. Polym. 2018, 184, 243–259. [Google Scholar] [CrossRef] [PubMed]
- Damonte, E.; Matulewicz, M.; Cerezo, A. Sulfated Seaweed Polysaccharides as Antiviral Agents. Curr. Med. Chem. 2004, 11, 2399–2419. [Google Scholar] [CrossRef]
- Liu, H.; Geng, M.; Xin, X.; Li, F.; Zhang, Z.; Li, J.; Ding, J. Multiple and Multivalent Interactions of Novel Anti-AIDS Drug Candidates, Sulfated Polymannuronate (SPMG)-Derived Oligosaccharides, with Gp120 and Their Anti-HIV Activities. Glycobiology 2005, 15, 501–510. [Google Scholar] [CrossRef]
- Ghosh, T.; Chattopadhyay, K.; Marschall, M.; Karmakar, P.; Mandal, P.; Ray, B. Focus on Antivirally Active Sulfated Polysaccharides: From Structure–Activity Analysis to Clinical Evaluation. Glycobiology 2009, 19, 2–15. [Google Scholar] [CrossRef]
- Giroglou, T.; Florin, L.; Schäfer, F.; Streeck, R.E.; Sapp, M. Human Papillomavirus Infection Requires Cell Surface Heparan Sulfate. J. Virol. 2001, 75, 1565–1570. [Google Scholar] [CrossRef] [Green Version]
- Ji, J.; Wang, L.; Wu, H.; Luan, H.-M. Bio-Function Summary of Marine Oligosaccharides. Int. J. Biol. 2010, 3, 74. [Google Scholar] [CrossRef] [Green Version]
- Witvrouw, M.; Schols, D.; Andrei, G.; Snoeck, R.; Hosoya, M.; Pauwels, R.; Balzarini, J.; De Clercq, E. Antiviral Activity of Low-MW Dextran Sulphate (Derived from Dextran MW 1000) Compared to Dextran Sulphate Samples of Higher MW. Antivir. Chem. Chemother. 1991, 2, 171–179. [Google Scholar] [CrossRef]
- Ivanova, V.; Rouseva, R.; Kolarova, M.; Serkedjieva, J.; Rachev, R.; Manolova, N. Isolation of a Polysaccharide with Antiviral Effect 927 from Ulva Lactuca. Prep. Biochem. 1994, 24, 83–97. [Google Scholar]
- Yu, N.; Liu, S.; Han, J.J.; Sun, F.S. The Depressive Effect of Glycosaminoglycan from Scallop on Type-I Herpes Simplex Virus. Acta Acad. Med. Qingdao Univ. 2008, 2, 111–114. [Google Scholar]
- El Hadrami, A.; Adam, L.R.; El Hadrami, I.; Daayf, F. Chitosan in Plant Protection. Mar. Drugs 2010, 8, 968–987. [Google Scholar] [CrossRef] [PubMed]
- Davydova, V.N.; Nagorskaia, V.P.; Gorbach, V.I.; Kalitnik, A.A.; Reunov, A.V.; Solov’eva, T.F.; Ermak, I.M. Chitosan Antiviral 1075 Activity: Dependence on Structure and Depolymerization Method. Prikl. Biokhim. Mikrobiol. 2011, 47, 113–118. [Google Scholar] [PubMed]
- Carlucci, M.J.; Scolaro, L.A.; Damonte, E.B. Herpes Simplex Virus Type 1 Variants Arising after Selection with an Antiviral Carrageenan: Lack of Correlation between Drug Susceptibility and Syn Phenotype. J. Med. Virol. 2002, 68, 92–98. [Google Scholar] [CrossRef]
- Maginnis, M.S. Virus–Receptor Interactions: The Key to Cellular Invasion. J. Mol. Biol. 2018, 430, 2590–2611. [Google Scholar] [CrossRef]
- Grassauer, A.; Weinmuellner, R.; Meier, C.; Pretsch, A.; Prieschl-Grassauer, E.; Unger, H. Iota-Carrageenan Is a Potent Inhibitor of Rhinovirus Infection. Virol. J. 2008, 5, 107. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Han, W.; Wang, G.; Zhao, X. Application Prospect of Polysaccharides in the Development of Anti-Novel Coronavirus Drugs and Vaccines. Int. J. Biol. Macromol. 2020, 164, 331–343. [Google Scholar] [CrossRef]
- Guo, C.; Zhu, Z.; Yu, P.; Zhang, X.; Dong, W.; Wang, X.; Chen, Y.; Liu, X. Inhibitory Effect of Iota-Carrageenan on Porcine Reproductive and Respiratory Syndrome Virus in Vitro. Antivir. Ther. 2019, 24, 261–270. [Google Scholar] [CrossRef]
- González, M.E.; Alarcón, B.; Carrasco, L. Polysaccharides as Antiviral Agents: Antiviral Activity of Carrageenan. Antimicrob. Agents Chemother. 1987, 31, 1388–1393. [Google Scholar] [CrossRef] [Green Version]
- Talarico, L.B.; Noseda, M.D.; Ducatti, D.R.B.; Duarte, M.E.R.; Damonte, E.B. Differential Inhibition of Dengue Virus Infection in Mammalian and Mosquito Cells by Iota-Carrageenan. J. Gen. Virol. 2011, 92, 1332–1342. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, P.; Hao, C.; Zhang, X.-E.; Cui, Z.-Q.; Guan, H.-S. In Vitro Inhibitory Effect of Carrageenan Oligosaccharide on Influenza A H1N1 Virus. Antivir. Res. 2011, 92, 237–246. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Therapeutic Class | Mechanism | Type | Target Viruses | Antivirals | Reference |
|---|---|---|---|---|---|
| Cell entry inhibitors | Fusion inhibitors | Polyanions, nonspecific inhibitors | Enveloped viruses: HIV, HSV, CMV, RSV | Polyanions in development: cosalane derivatives, polyoxometalates, polycarboxylates polysulfates, polysulfonates, negatively charged albumins, sulphated polysaccharides | [56] |
| Receptor and co-receptor antagonist analogues | HIV-1 | Maraviroc | [52,54] | ||
| Fusion and penetration inhibitors | Fusion inhibitor | HIV-1 | Enfuvirtide | [53,55] | |
| Uncoating inhibitor | Influenza A | Amantadine, rimantadine | [57] | ||
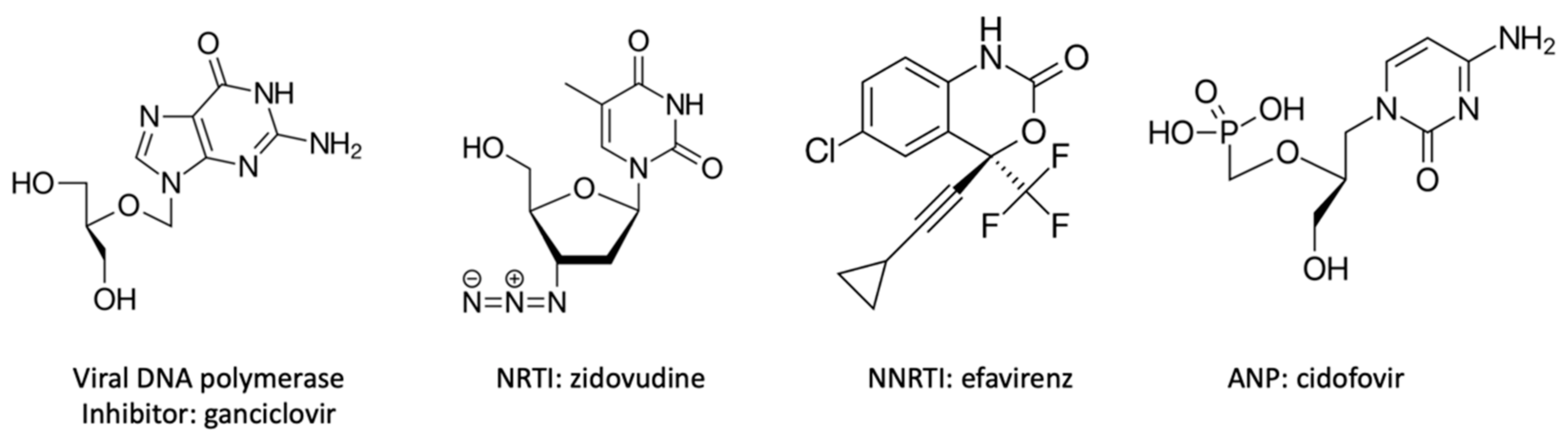
| Replication inhibitors | Nucleosides analogue inhibitors (activation by triphosphorylation) with competitive inhibition | Viral DNA polymerase inhibitors | Herpes viruses (HSV-1, HSV-2, VZV, CMV, EBV, HHV-6, HBV) | Foscarnet, acyclovir, famciclovir, brivudine, valganciclovir, penciclovir, ganciclovir, entecavir, telbuvine | [40,58,59] |
| Reverse transcriptase inhibitors | HIV | NRTIs: zidovudine, didanosine, zalcitabine, stavudine, abacavir, emtricitabine, lamivudine | [60,61] | ||
| Non-nucleoside inhibitors | NNRTIs: efavirenz, nevirapine, delavirdine, etravirine | [62,63,64] | |||
| Acyclic nucleoside phosphonates (activation by diphosphorylation) | Acyclic nucleoside phosphonates (ANP) | Herpes viruses (HSV-1, HSV-2, VZV, CMV, HHV-6, EBV, HBV), HIV-1, HIV-2 | Cidofovir, adefovir, tenofovir | [70] | |
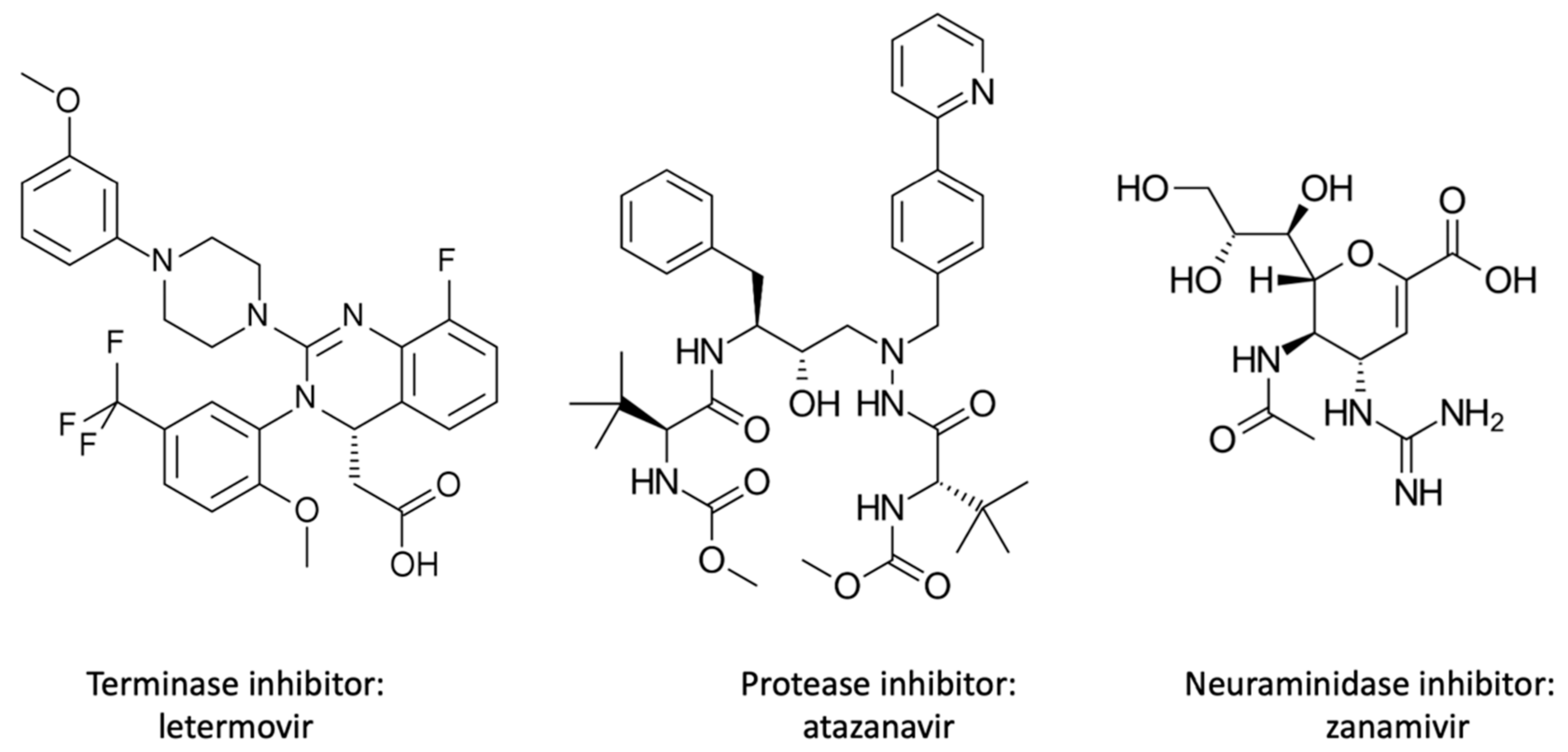
| Assembly, maturation, and release inhibitors | Packaging inhibitor | Viral terminase enzyme complex inhibitors | CMV | Letermovir | [65] |
| Maturation inhibitor | Viral protease inhibitors (polyprotein precursors cleavage) | HIV-1, HIV-2 | Amprenavir, fosamprenavir, atazanavir, indinavir, lopinavir, nelfinavir, ritonavir, saquinavir, darunavir, tipranavir, atazanavir | [66,67] | |
| Release inhibitors | Viral neuraminidase inhibitors | Influenza A and B | Zanamivir, oseltamivir, pramivir | [68,69] |
| Origin | Polysaccharides | Antiviral Effects | Mechanisms | References |
|---|---|---|---|---|
| ||||
| Bovine and porcine tissues | Heparin | Anti-CMV, HSV, HPV, DENV, JEV, YFV, ZIKV, HIV | Inhibition of adsorption | [78,79,80] |
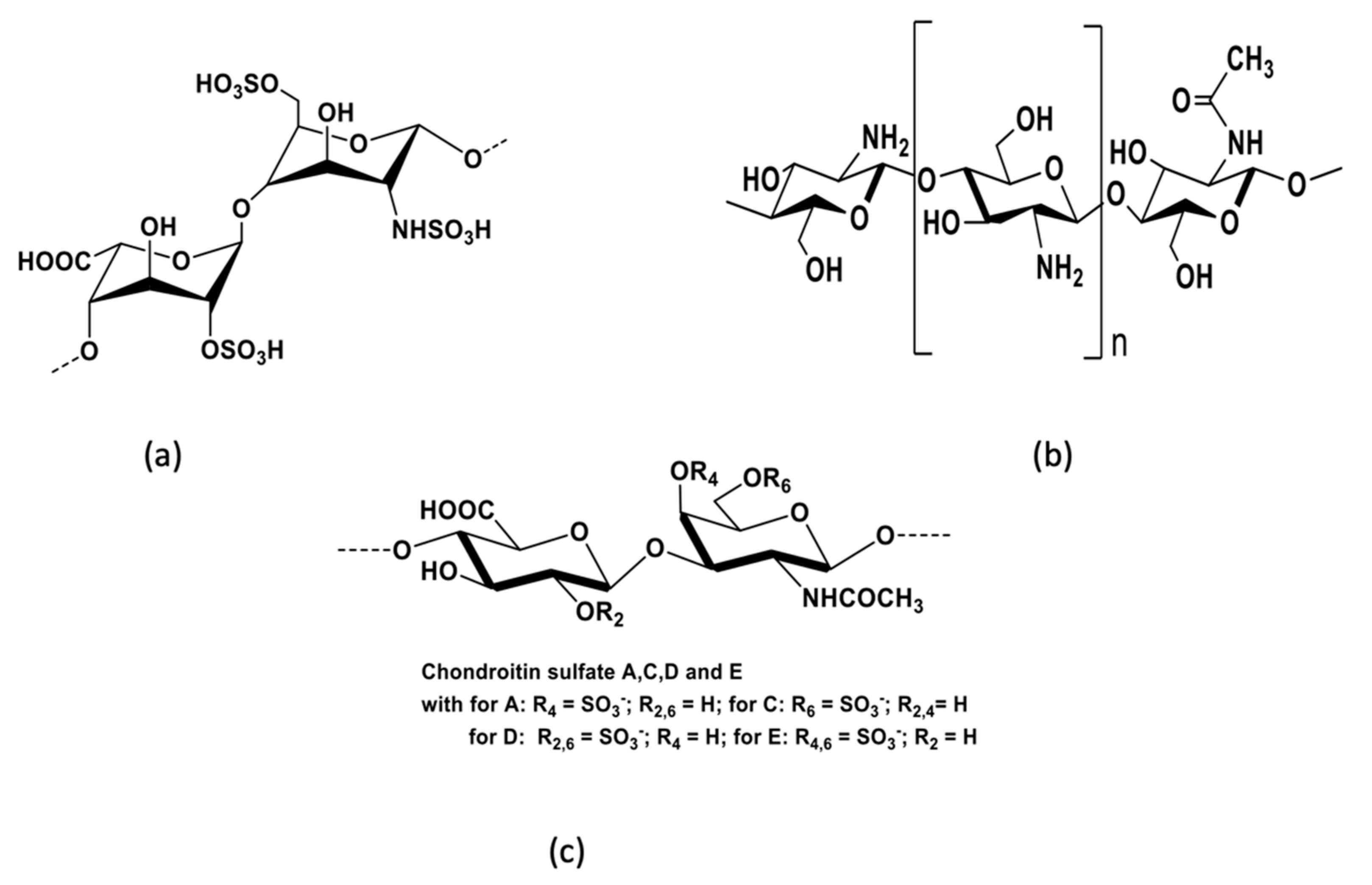
| Cartilage of animals | Chondroitin sulfate | Anti-HIV, HSV, HTLV-1, DENV | Inhibition of adsorption | [81,82,83,84] |
| Crustacean shell | Chitosan | Anti-bacteriophages, plant viruses, HIV, FCV-F9 | Direct virucidal effect | [85,86] |
| Shellfish | Shellfish polysaccharide | Anti-HBV, HIV-1, IFV, HSV-1 | Inhibition of adsorption | [87,88] |
| ||||
| Brown algae | Fucoidan | Anti-HIV, DENV, IAV, NDV, HSV | Inhibition of adsorption and transcription | [89,90,91] |
| Laminarin | Anti-HIV | Inhibition of adsorption and transcription | [92] | |
| Alginate | Anti-HIV, HBV, IAV, TMV | Inhibition of transcription | [93,94] | |
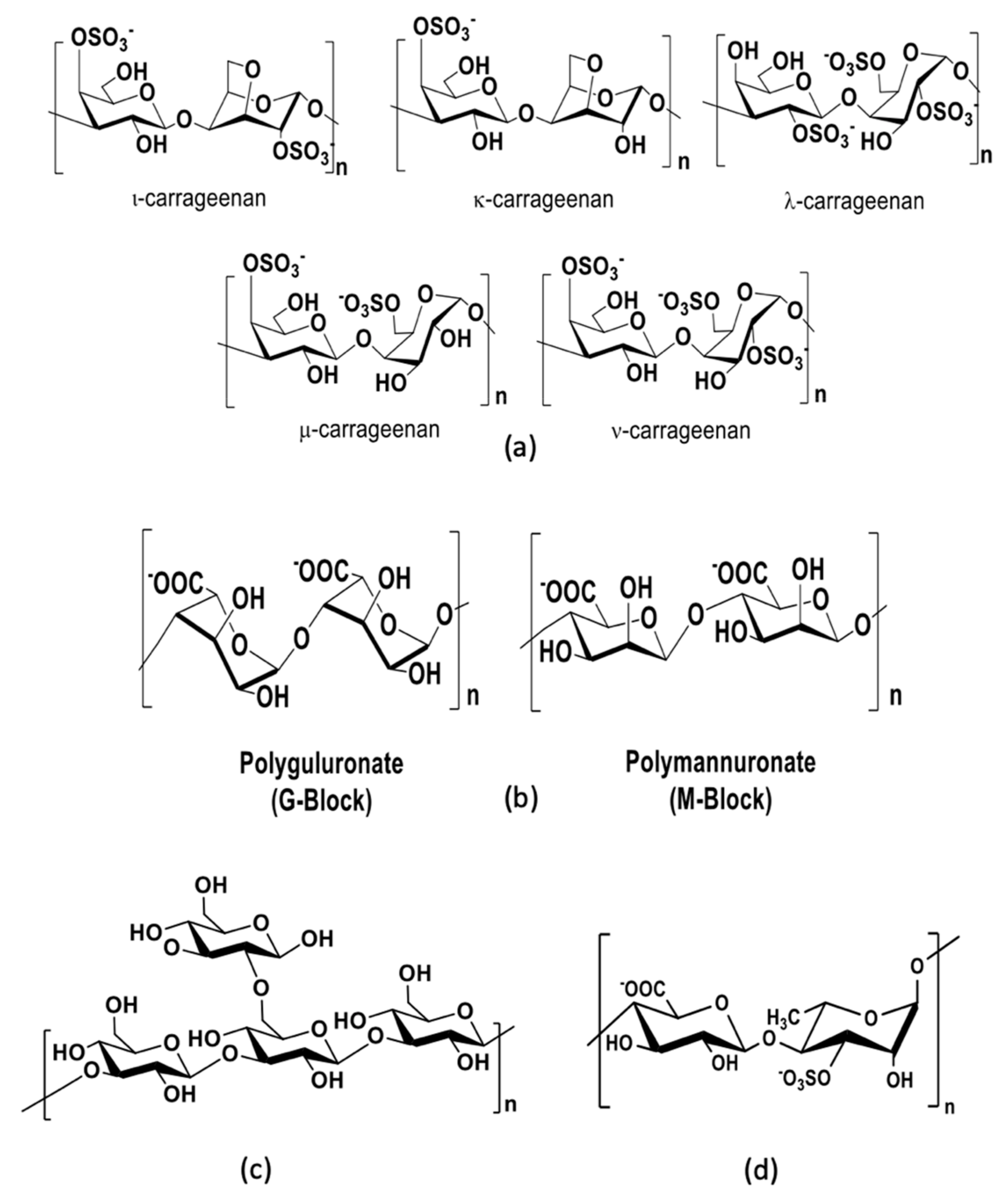
| Red algae | ι-carrageenan | Anti-DENV, HAV, HRV, IAV, HPV, ECMV, HSV, SFV, ASV, VV | Inhibition of adsorption, penetration, uncoating, replication and transcription | [82,92,95,96,97,98,99] |
| κ-carrageenan | Anti-enterovirus, IAV, HSV, CMV, Sindbis virus, VV, HIV, DENV, TMV | Inhibition of replication and transcription | [97,100] | |
| λ-carrageenan | Anti-DENV, HSV, HPV, HAV, CMV, HIV, Sindbis virus, VV, BoHV-1, SuHV-1, RABV | Direct virucidal effect, inhibition of adsorption | [95,97,101,102] | |
| β-carrageenan | TMV | - | [103] | |
| Green algae | Ulvan | Anti-IFV-A, JEV, NDV | Inhibition of penetration and uncoating | [104,105,106,107,108] |
| Diatom (microalga) | Naviculan | Anti-HIV, HSV-1, HSV-2, IFV | Inhibition of adsorption | [109] |
| Microalgae | Sulfated polysaccharide | Anti IAV, HIV-1, HSV-1, IFV-A, IFV-B, RSV-A, RSV-B, PIFV-2, EMCV | Inhibition of penetration and replication | [76,78,110,111,112,113] |
| Edible | Edible polysaccharide | Anti-DEV, HSV-1 | Inhibition of adsorption, penetration and uncoating | [114,115] |
| Medicinal plants | Medicinal plant polysaccharide | Anti-HSV-2, PV-1, PV-2, DHAV, BoHV-1, HBV, H1N1, H3N2, HIV, rotavirus | Inhibition of adsorption and replication | [116,117] |
| ||||
| Mushrooms | Mushroom polysaccharide | Anti-IFV-A *, PV-1, HIV-1, HBV, IHNV | Inhibition of transcription and replication | [118,119,120] |
| ||||
| Marine bacterias | EPS | Anti-HSV-2, HSV-1 | Inhibition of adsorption | [121] |
| Cyanobacterias | Calcium spirulan | Anti-HSV-1, HCMV, measles virus, mumps virus, IAV, HIV-1 | Inhibition of adsorption and replication | [122,123] |
| Nostoflan | Anti-HSV-1, HSV-2, IAV, HCMV | Inhibition of adsorption | [124,125] | |
| Source | Type of Extract | Virus Involved | Object of Study | Antiviral Activity | Results | Reference |
|---|---|---|---|---|---|---|
| S. ilicifolium (seaweed) | Water extraction | Betanodavirus | Fishes | Cytopathic effect reduction assay | Significant antiviral activity. | [144] |
| Salvia plebeia R. Br. | Alcohol precipitation | Respiratory syncytial virus (RSV) | mice | In vitro and in vivo antiviral | Fractions (Mw ≥ 10,000 Da) inhibit the RSV proliferation and reduce the lung lesions induced by RSV. | [145] |
| Red algae | Water extraction | Influenza viruses SARS-CoV-2 | Madin–Darby canine kidney cells, African green monkey kidney cells, human embryonic kidney (HEK) 293 T cells, mice. | In vitro and in vivo antiviral | λ-carrageenan reduced expression of viral proteins in cell lysates and suppressed progeny virus production. A dose-dependent effect was reported. | [142] |
| Asarum polysaccharides | Water extraction and alcohol precipitation | H1N1 influenza virus | Cell real-time monitoring system and Reed-Muench mice model | In vitro and in vivo antiviral | Good anti-influenza virus activity. | [146] |
| Isatidis radix | Ethanol extraction | Hepatitis B virus (HBV) | HepG2.2.15 cell | In vitro antiviral | No toxicity at <400 μg/mL. Doses of 50, 100, and 200 μg/mL significantly reduced extracellular and intracellular levels of HBsAg, HBeAg, and HBV DNA in HepG2.2.15 cells. | [147] |
| Pleurotus pulmonarius | Water and ethanol extraction | Influenza virus A California/07/09 (H1N1pdm) | The neutral red adsorption test was used in the study | In vitro antiviral | The ethanol extracts exhibit a more potent antiviral effect than that of water extracts. Weak toxicity was reported. | [148] |
| Modifications | Polysaccharides | Antiviral Effects | Mechanisms | References |
|---|---|---|---|---|
| Sulfatation | Dextran sulfate | Anti-ZIKV, IAV *, enveloped viruses | Inhibition of adsorption and replication | [153,154,155] |
| 3-O-Sulfated heparin | Anti-HSV-1 | Inhibition of adsorption | [156] | |
| MI-S, FR-S | Anti HSV-1 *, HSV-2 * | Inhibition of adsorption | [157,158] | |
| sAAPs, sTPS | Anti-NDV | - | [159,160] | |
| SPLCf | Anti HSV, PV-1 | Inhibition of adsorption and transcription | [161] | |
| sCVPS | Anti-NDV * | - | [162] | |
| S1F1, S2F1 | Anti HSV-1, HSV-2 | - | [163] | |
| NCMCS | Anti-HIV-1, RLV | Inhibition of virus adsorption | [164] | |
| κ-carrageenan | Anti-IAV *, HIV | Inhibition of replication | [165] | |
| Phosphorylation | pRCPS, pCIPS | Anti-DHAV | Inhibition of replication | [166,167] |
| pRCPS | Anti-CPV | - | [168] | |
| Complexes | Chitosan–chondroitin sulfate | Anti-HIV | Inhibition of replication | [169] |
| Enzymatic modification | Chitooligosaccharides | Anti-HIV-1 | Inhibition of adsorption | [170] |
| Acylation + sulfation | O-acylated carrageenans | Anti-HIV | - | [171] |
| Aminoderivatized | Aminoethyl-chitosan | Anti-HIV-1 | - | [172] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Claus-Desbonnet, H.; Nikly, E.; Nalbantova, V.; Karcheva-Bahchevanska, D.; Ivanova, S.; Pierre, G.; Benbassat, N.; Katsarov, P.; Michaud, P.; Lukova, P.; et al. Polysaccharides and Their Derivatives as Potential Antiviral Molecules. Viruses 2022, 14, 426. https://doi.org/10.3390/v14020426
Claus-Desbonnet H, Nikly E, Nalbantova V, Karcheva-Bahchevanska D, Ivanova S, Pierre G, Benbassat N, Katsarov P, Michaud P, Lukova P, et al. Polysaccharides and Their Derivatives as Potential Antiviral Molecules. Viruses. 2022; 14(2):426. https://doi.org/10.3390/v14020426
Chicago/Turabian StyleClaus-Desbonnet, Hadrien, Elsa Nikly, Vanya Nalbantova, Diana Karcheva-Bahchevanska, Stanislava Ivanova, Guillaume Pierre, Niko Benbassat, Plamen Katsarov, Philippe Michaud, Paolina Lukova, and et al. 2022. "Polysaccharides and Their Derivatives as Potential Antiviral Molecules" Viruses 14, no. 2: 426. https://doi.org/10.3390/v14020426
APA StyleClaus-Desbonnet, H., Nikly, E., Nalbantova, V., Karcheva-Bahchevanska, D., Ivanova, S., Pierre, G., Benbassat, N., Katsarov, P., Michaud, P., Lukova, P., & Delattre, C. (2022). Polysaccharides and Their Derivatives as Potential Antiviral Molecules. Viruses, 14(2), 426. https://doi.org/10.3390/v14020426