

Impact of Dextran-Sodium-Sulfate-Induced Enteritis on Murine Cytomegalovirus Reactivation

 , , ,
, , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice, Viral Strain and Stock
2.2. Experimental Design
2.3. Clinical Score
2.4. Measure of Fecal Lipocalin
2.5. Collection of Organ Samples
2.6. Histopathology
2.7. Immunohistochemistry
2.8. MCMV qPCR in Mouse Blood and Tissues
2.9. Statistics
2.10. Ethical Considerations
3. Results
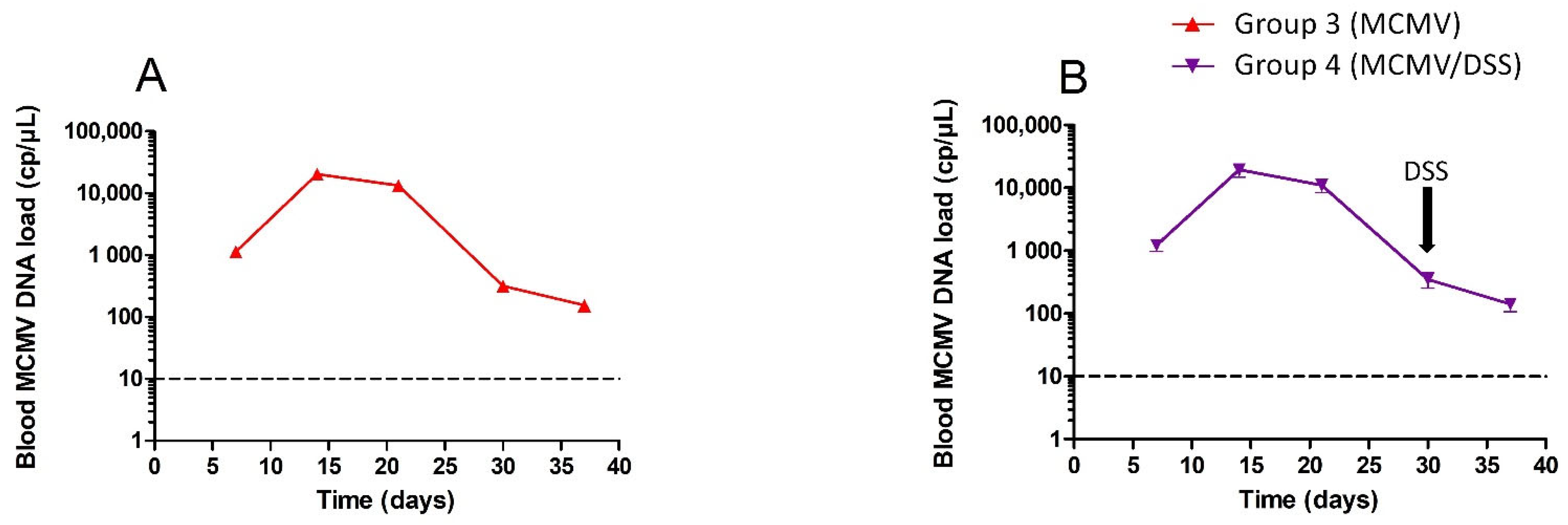
3.1. Monitoring of MCMV Systemic Infection
3.2. Clinical Assessment of DSS Administration
3.3. Severe Intestinal Inflammation Is Induced by DSS, Regardless of MCMV Infection Status
3.4. Absence of MCMV Reactivation in DSS Model of Gut Inflammation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Hibi, T.; Ogata, H. Novel Pathophysiological Concepts of Inflammatory Bowel Disease. J. Gastroenterol. 2006, 41, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, G.G. The Global Burden of IBD: From 2015 to 2025. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.C.Y.; Chan, F.K.L.; et al. Worldwide Incidence and Prevalence of Inflammatory Bowel Disease in the 21st Century: A Systematic Review of Population-Based Studies. Lancet 2017, 390, 2769–2778. [Google Scholar] [CrossRef] [PubMed]
- Mak, W.Y.; Zhao, M.; Ng, S.C.; Burisch, J. The Epidemiology of Inflammatory Bowel Disease: East Meets West. J. Gastroenterol. Hepatol. 2020, 35, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Ramos, G.P.; Papadakis, K.A. Mechanisms of Disease: Inflammatory Bowel Diseases. Mayo Clin. Proc. 2019, 94, 155–165. [Google Scholar] [CrossRef]
- Kucharzik, T.; Ellul, P.; Greuter, T.; Rahier, J.F.; Verstockt, B.; Abreu, C.; Albuquerque, A.; Allocca, M.; Esteve, M.; Farraye, F.A.; et al. ECCO Guidelines on the Prevention, Diagnosis, and Management of Infections in Inflammatory Bowel Disease. J. Crohns. Colitis 2021, 15, 879–913. [Google Scholar] [CrossRef] [PubMed]
- Zuhair, M.; Smit, G.S.A.; Wallis, G.; Jabbar, F.; Smith, C.; Devleesschauwer, B.; Griffiths, P. Estimation of the Worldwide Seroprevalence of Cytomegalovirus: A Systematic Review and Meta-Analysis. Rev. Med. Virol. 2019, 29, e2034. [Google Scholar] [CrossRef]
- Gugliesi, F.; Coscia, A.; Griffante, G.; Galitska, G.; Pasquero, S.; Albano, C.; Biolatti, M. Where Do We Stand after Decades of Studying Human Cytomegalovirus? Microorganisms 2020, 8, 685. [Google Scholar] [CrossRef]
- Collins-McMillen, D.; Buehler, J.; Peppenelli, M.; Goodrum, F. Molecular Determinants and the Regulation of Human Cytomegalovirus Latency and Reactivation. Viruses 2018, 10, 444. [Google Scholar] [CrossRef]
- Nakase, H.; Honzawa, Y.; Toyonaga, T.; Yamada, S.; Minami, N.; Yoshino, T.; Matsuura, M. Diagnosis and Treatment of Ulcerative Colitis with Cytomegalovirus Infection: Importance of Controlling Mucosal Inflammation to Prevent Cytomegalovirus Reactivation. Intest. Res. 2014, 12, 5–11. [Google Scholar] [CrossRef][Green Version]
- Jentzer, A.; Veyrard, P.; Roblin, X.; Saint-Sardos, P.; Rochereau, N.; Paul, S.; Bourlet, T.; Pozzetto, B.; Pillet, S. Cytomegalovirus and Inflammatory Bowel Diseases (IBD) with a Special Focus on the Link with Ulcerative Colitis (UC). Microorganisms 2020, 8, 1078. [Google Scholar] [CrossRef] [PubMed]
- Zagórowicz, E.; Bugajski, M.; Wieszczy, P.; Pietrzak, A.; Magdziak, A.; Mróz, A. Cytomegalovirus Infection in Ulcerative Colitis Is Related to Severe Inflammation and a High Count of Cytomegalovirus-Positive Cells in Biopsy Is a Risk Factor for Colectomy. J. Crohns. Colitis 2016, 10, 1205–1211. [Google Scholar] [CrossRef]
- Schenk, W.; Klugmann, T.; Borkenhagen, A.; Klecker, C.; Dietel, P.; Kirschner, R.; Schneider, E.; Bruns, T.; Stallmach, A.; Teich, N. The Detection of the Cytomegalovirus DNA in the Colonic Mucosa of Patients with Ulcerative Colitis Is Associated with Increased Long-Term Risk of Proctocolectomy: Results from an Outpatient IBD Clinic. Int. J. Colorectal. Dis. 2019, 34, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-S.; Park, S.H.; Kim, S.-H.; Kim, J.; Choi, J.; Lee, H.J.; Kim, W.S.; Lee, J.-M.; Kwak, M.S.; Hwang, S.W.; et al. Risk Factors and Clinical Outcomes Associated with Cytomegalovirus Colitis in Patients with Acute Severe Ulcerative Colitis. Inflamm. Bowel Dis. 2016, 22, 912–918. [Google Scholar] [CrossRef] [PubMed]
- Kishore, J.; Ghoshal, U.; Ghoshal, U.C.; Krishnani, N.; Kumar, S.; Singh, M.; Ayyagari, A. Infection with Cytomegalovirus in Patients with Inflammatory Bowel Disease: Prevalence, Clinical Significance and Outcome. J. Med. Microbiol. 2004, 53, 1155–1160. [Google Scholar] [CrossRef] [PubMed]
- Roblin, X.; Pillet, S.; Oussalah, A.; Berthelot, P.; Del Tedesco, E.; Phelip, J.-M.; Chambonnière, M.-L.; Garraud, O.; Peyrin-Biroulet, L.; Pozzetto, B. Cytomegalovirus Load in Inflamed Intestinal Tissue Is Predictive of Resistance to Immunosuppressive Therapy in Ulcerative Colitis. Am. J. Gastroenterol. 2011, 106, 2001–2008. [Google Scholar] [CrossRef]
- Pillet, S.; Pozzetto, B.; Jarlot, C.; Paul, S.; Roblin, X. Management of Cytomegalovirus Infection in Inflammatory Bowel Diseases. Dig. Liver Dis. 2012, 44, 541–548. [Google Scholar] [CrossRef]
- Pillet, S. Cytomegalovirus and Ulcerative Colitis: Place of Antiviral Therapy. World J. Gastroenterol. 2016, 22, 2030. [Google Scholar] [CrossRef]
- Onyeagocha, C.; Hossain, M.S.; Kumar, A.; Jones, R.M.; Roback, J.; Gewirtz, A.T. Latent Cytomegalovirus Infection Exacerbates Experimental Colitis. Am. J. Pathol. 2009, 175, 2034–2042. [Google Scholar] [CrossRef]
- Brunson, J.L.; Becker, F.; Stokes, K.Y. The Impact of Primary and Persistent Cytomegalovirus Infection on the Progression of Acute Colitis in a Murine Model. Pathophysiology 2015, 22, 31–37. [Google Scholar] [CrossRef]
- Eichele, D.D.; Kharbanda, K.K. Dextran Sodium Sulfate Colitis Murine Model: An Indispensable Tool for Advancing Our Understanding of Inflammatory Bowel Diseases Pathogenesis. World J. Gastroenterol. 2017, 23, 6016–6029. [Google Scholar] [CrossRef] [PubMed]
- Lutarewych, M.A.; Quirk, M.R.; Kringstad, B.A.; Li, W.; Verfaillie, C.M.; Jordan, M.C. Propagation and Titration of Murine Cytomegalovirus in a Continuous Bone Marrow-Derived Stromal Cell Line (M2-10B4). J. Virol. Methods 1997, 68, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Schippers, A.; Muschaweck, M.; Clahsen, T.; Tautorat, S.; Grieb, L.; Tenbrock, K.; Gaßler, N.; Wagner, N. Β7-Integrin Exacerbates Experimental DSS-Induced Colitis in Mice by Directing Inflammatory Monocytes into the Colon. Mucosal. Immunol. 2016, 9, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Chassaing, B.; Srinivasan, G.; Delgado, M.A.; Young, A.N.; Gewirtz, A.T.; Vijay-Kumar, M. Fecal Lipocalin 2, a Sensitive and Broadly Dynamic Non-Invasive Biomarker for Intestinal Inflammation. PLoS ONE 2012, 7, e44328. [Google Scholar] [CrossRef] [PubMed]
- Marchal-Bressenot, A.; Salleron, J.; Boulagnon-Rombi, C.; Bastien, C.; Cahn, V.; Cadiot, G.; Diebold, M.-D.; Danese, S.; Reinisch, W.; Schreiber, S.; et al. Development and Validation of the Nancy Histological Index for UC. Gut 2017, 66, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Rochereau, N.; Roblin, X.; Michaud, E.; Gayet, R.; Chanut, B.; Jospin, F.; Corthésy, B.; Paul, S. NOD2 Deficiency Increases Retrograde Transport of Secretory IgA Complexes in Crohn’s Disease. Nat. Commun. 2021, 12, 261. [Google Scholar] [CrossRef]
- Pillet, S.; Bourlet, T.; Pozzetto, B. Comparative Evaluation of the QIAsymphony RGQ System with the EasyMAG/R-Gene Combination for the Quantitation of Cytomegalovirus DNA Load in Whole Blood. Virol. J. 2012, 9, 231. [Google Scholar] [CrossRef]
- Khairallah, C.; Netzer, S.; Villacreces, A.; Juzan, M.; Rousseau, B.; Dulanto, S.; Giese, A.; Costet, P.; Praloran, V.; Moreau, J.-F.; et al. Γδ T Cells Confer Protection against Murine Cytomegalovirus (MCMV). PLoS Pathog. 2015, 11, e1004702. [Google Scholar] [CrossRef]
- Pillet, S.; Williet, N.; Pouvaret, A.; Del Tedesco, E.; Saint-Sardos, P.; Pozzetto, B.; Roblin, X. Distribution of Cytomegalovirus DNA Load in the Inflamed Colon of Ulcerative Colitis Patients. Am. J. Gastroenterol. 2016, 111, 439–441. [Google Scholar] [CrossRef]
- Forte, E.; Zhang, Z.; Thorp, E.B.; Hummel, M. Cytomegalovirus Latency and Reactivation: An Intricate Interplay with the Host Immune Response. Front. Cell. Infect. Microbiol. 2020, 10, 130. [Google Scholar] [CrossRef]
- Balthesen, M.; Messerle, M.; Reddehase, M.J. Lungs Are a Major Organ Site of Cytomegalovirus Latency and Recurrence. J. Virol. 1993, 67, 5360–5366. [Google Scholar] [CrossRef] [PubMed]
- Pomeroy, C.; Hilleren, P.J.; Jordan, M.C. Latent Murine Cytomegalovirus DNA in Splenic Stromal Cells of Mice. J. Virol. 1991, 65, 3330–3334. [Google Scholar] [CrossRef] [PubMed]
- Klotman, M.E.; Henry, S.C.; Greene, R.C.; Brazy, P.C.; Klotman, P.E.; Hamilton, J.D. Detection of Mouse Cytomegalovirus Nucleic Acid in Latently Infected Mice by in Vitro Enzymatic Amplification. J. Infect. Dis. 1990, 161, 220–225. [Google Scholar] [CrossRef]
- Pollock, J.L.; Virgin, H.W. Latency, without Persistence, of Murine Cytomegalovirus in the Spleen and Kidney. J. Virol. 1995, 69, 1762–1768. [Google Scholar] [CrossRef]
- Watanabe, H.; Numata, K.; Ito, T.; Takagi, K.; Matsukawa, A. Innate Immune Response in Th1- and Th2-Dominant Mouse Strains. Shock 2004, 22, 460–466. [Google Scholar] [CrossRef]
- Neurath, M.F. Targeting Immune Cell Circuits and Trafficking in Inflammatory Bowel Disease. Nat. Immunol. 2019, 20, 970–979. [Google Scholar] [CrossRef]
- Bhan, A.K.; Mizoguchi, E.; Smith, R.N.; Mizoguchi, A. Spontaneous Chronic Colitis in TCR Alpha-Mutant Mice; an Experimental Model of Human Ulcerative Colitis. Int. Rev. Immunol. 2000, 19, 123–138. [Google Scholar] [CrossRef]
- Matsumura, K.; Nakase, H.; Kosugi, I.; Honzawa, Y.; Yoshino, T.; Matsuura, M.; Kawasaki, H.; Arai, Y.; Iwashita, T.; Nagasawa, T.; et al. Establishment of a Novel Mouse Model of Ulcerative Colitis with Concomitant Cytomegalovirus Infection: In Vivo Identification of Cytomegalovirus Persistent Infected Cells. Inflamm. Bowel Dis. 2013, 19, 1951–1963. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Kolachala, V.; Dalmasso, G.; Nguyen, H.; Laroui, H.; Sitaraman, S.V.; Merlin, D. Temporal and Spatial Analysis of Clinical and Molecular Parameters in Dextran Sodium Sulfate Induced Colitis. PLoS ONE 2009, 4, e6073. [Google Scholar] [CrossRef]
- Mai, C.-T.; Wu, M.-M.; Wang, C.-L.; Su, Z.-R.; Cheng, Y.-Y.; Zhang, X.-J. Palmatine Attenuated Dextran Sulfate Sodium (DSS)-Induced Colitis via Promoting Mitophagy-Mediated NLRP3 Inflammasome Inactivation. Mol. Immunol. 2019, 105, 76–85. [Google Scholar] [CrossRef]
- Hummel, M.; Zhang, Z.; Yan, S.; DePlaen, I.; Golia, P.; Varghese, T.; Thomas, G.; Abecassis, M.I. Allogeneic Transplantation Induces Expression of Cytomegalovirus Immediate-Early Genes in Vivo: A Model for Reactivation from Latency. J. Virol. 2001, 75, 4814–4822. [Google Scholar] [CrossRef] [PubMed]
- Bekiaris, V.; Timoshenko, O.; Hou, T.Z.; Toellner, K.; Shakib, S.; Gaspal, F.; McConnell, F.M.; Parnell, S.M.; Withers, D.; Buckley, C.D.; et al. Ly49H+ NK Cells Migrate to and Protect Splenic White Pulp Stroma from Murine Cytomegalovirus Infection. J. Immunol. 2008, 180, 6768–6776. [Google Scholar] [CrossRef]
- Wirtz, S.; Popp, V.; Kindermann, M.; Gerlach, K.; Weigmann, B.; Fichtner-Feigl, S.; Neurath, M.F. Chemically Induced Mouse Models of Acute and Chronic Intestinal Inflammation. Nat. Protoc. 2017, 12, 1295–1309. [Google Scholar] [CrossRef]
- Kiesler, P.; Fuss, I.J.; Strober, W. Experimental Models of Inflammatory Bowel Diseases. Cell. Mol. Gastroenterol. Hepatol. 2015, 1, 154–170. [Google Scholar] [CrossRef]
- Heller, F.; Fuss, I.J.; Nieuwenhuis, E.E.; Blumberg, R.S.; Strober, W. Oxazolone Colitis, a Th2 Colitis Model Resembling Ulcerative Colitis, Is Mediated by IL-13-Producing NK-T Cells. Immunity 2002, 17, 629–638. [Google Scholar] [CrossRef]
- Krishna, B.A.; Lau, B.; Jackson, S.E.; Wills, M.R.; Sinclair, J.H.; Poole, E. Transient Activation of Human Cytomegalovirus Lytic Gene Expression during Latency Allows Cytotoxic T Cell Killing of Latently Infected Cells. Sci. Rep. 2016, 6, 24674. [Google Scholar] [CrossRef]
- Tang, Q.; Maul, G.G. Mouse Cytomegalovirus Immediate-Early Protein 1 Binds with Host Cell Repressors to Relieve Suppressive Effects on Viral Transcription and Replication during Lytic Infection. J. Virol. 2003, 77, 1357–1367. [Google Scholar] [CrossRef] [PubMed]
- Crawford, L.B.; Tempel, R.; Streblow, D.N.; Kreklywich, C.; Smith, P.; Picker, L.J.; Nelson, J.A.; Caposio, P. Human Cytomegalovirus Induces Cellular and Humoral Virus-Specific Immune Responses in Humanized BLT Mice. Sci. Rep. 2017, 7, 937. [Google Scholar] [CrossRef]
- Itell, H.L.; Kaur, A.; Deere, J.D.; Barry, P.A.; Permar, S.R. Rhesus Monkeys for a Nonhuman Primate Model of Cytomegalovirus Infections. Curr. Opin. Virol. 2017, 25, 126–133. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jentzer, A.; Fauteux-Daniel, S.; Verhoeven, P.; Cantais, A.; Novoa, M.Y.; Jospin, F.; Chanut, B.; Rochereau, N.; Bourlet, T.; Roblin, X.; et al. Impact of Dextran-Sodium-Sulfate-Induced Enteritis on Murine Cytomegalovirus Reactivation. Viruses 2022, 14, 2595. https://doi.org/10.3390/v14122595
Jentzer A, Fauteux-Daniel S, Verhoeven P, Cantais A, Novoa MY, Jospin F, Chanut B, Rochereau N, Bourlet T, Roblin X, et al. Impact of Dextran-Sodium-Sulfate-Induced Enteritis on Murine Cytomegalovirus Reactivation. Viruses. 2022; 14(12):2595. https://doi.org/10.3390/v14122595
Chicago/Turabian StyleJentzer, Alexandre, Sébastien Fauteux-Daniel, Paul Verhoeven, Aymeric Cantais, Melyssa Yaugel Novoa, Fabienne Jospin, Blandine Chanut, Nicolas Rochereau, Thomas Bourlet, Xavier Roblin, and et al. 2022. "Impact of Dextran-Sodium-Sulfate-Induced Enteritis on Murine Cytomegalovirus Reactivation" Viruses 14, no. 12: 2595. https://doi.org/10.3390/v14122595
APA StyleJentzer, A., Fauteux-Daniel, S., Verhoeven, P., Cantais, A., Novoa, M. Y., Jospin, F., Chanut, B., Rochereau, N., Bourlet, T., Roblin, X., Pozzetto, B., & Pillet, S. (2022). Impact of Dextran-Sodium-Sulfate-Induced Enteritis on Murine Cytomegalovirus Reactivation. Viruses, 14(12), 2595. https://doi.org/10.3390/v14122595