

Expression of SARS-Cov-2 Entry Factors in Patients with Chronic Hepatitis

 , , , ,
, , , ,  , ,
, ,  , , , ,
, , , ,  , , ,
, , ,  ,
,  add
Show full author list
add
Show full author list
Abstract
1. Introduction
2. Materials and Methods
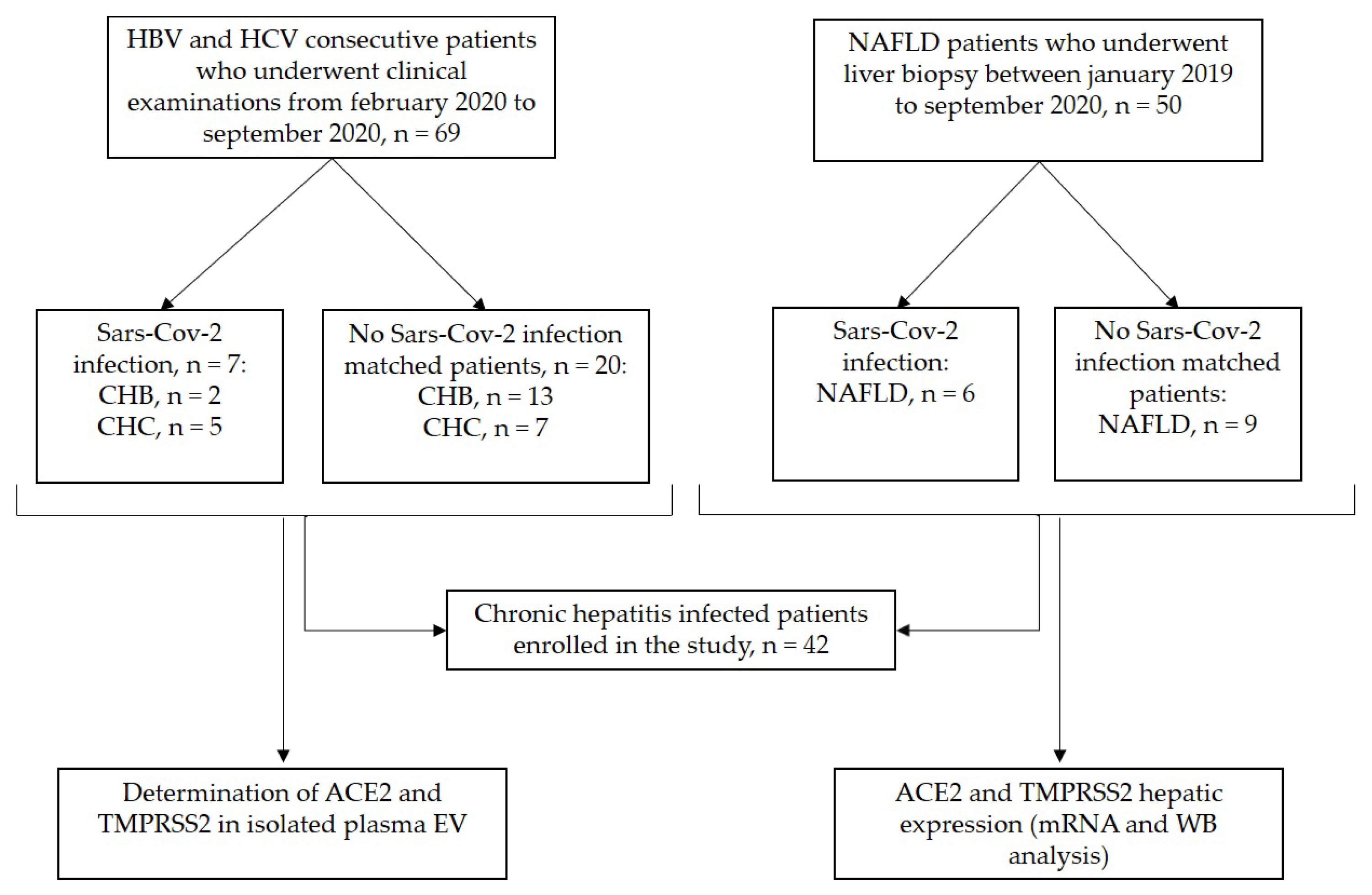
2.1. Study Population
2.2. Liver Biopsies
2.3. Western Blot Analysis
2.4. Circulating ACE2 and TMPRSS2 Determinations
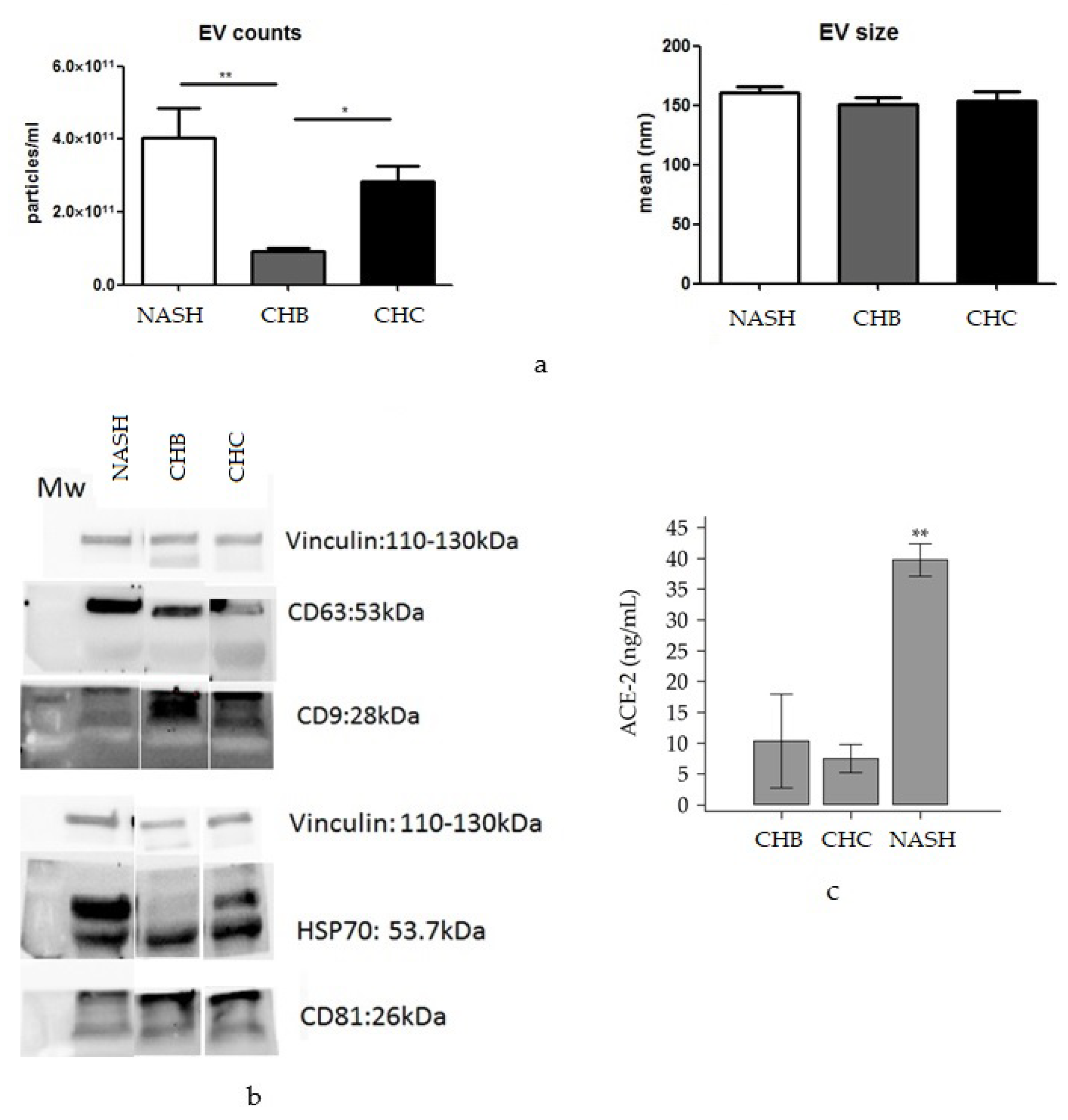
2.5. EV Isolation and Characterization, Western Blotting and ELISA
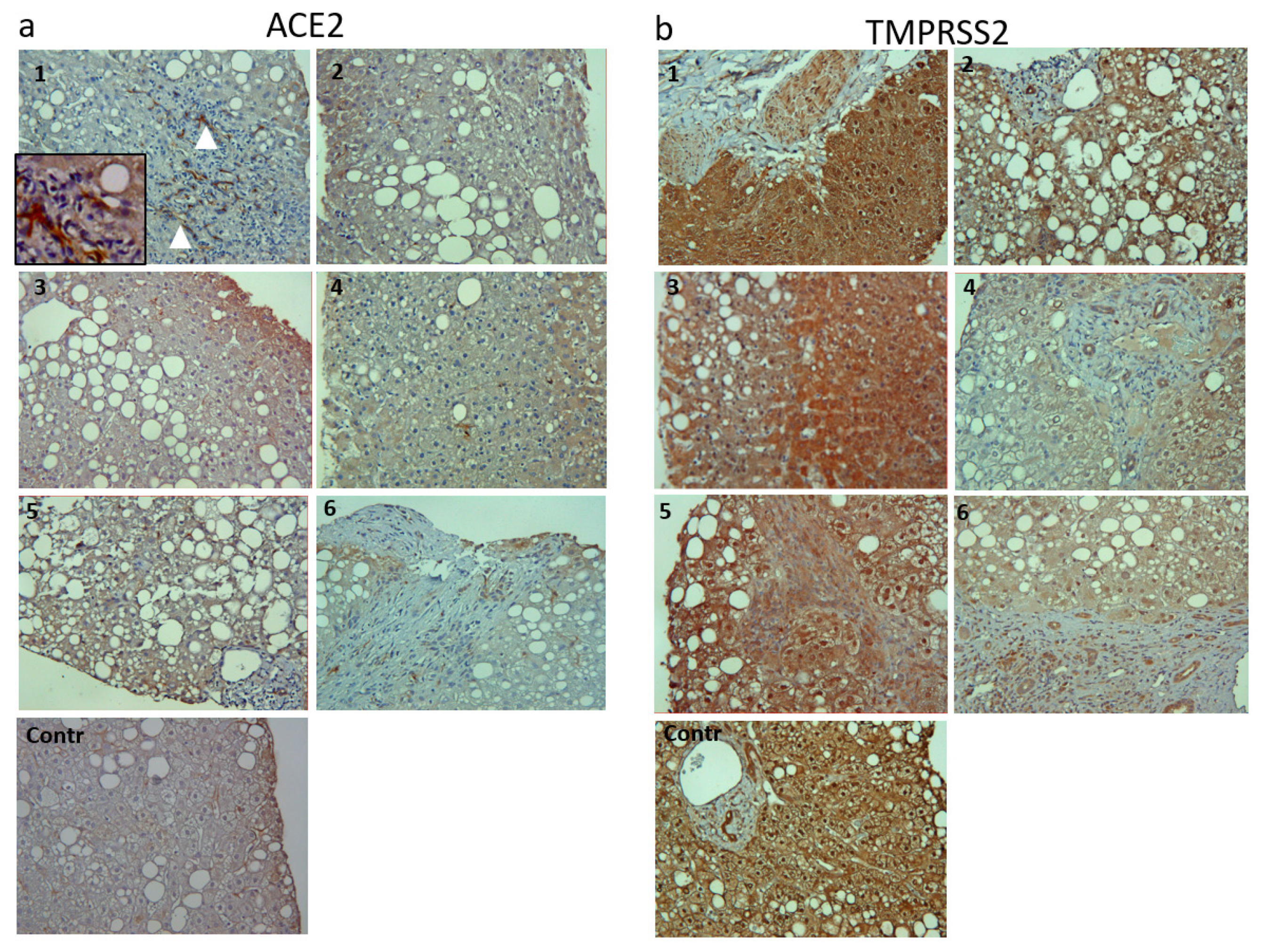
2.6. Immunohistochemistry
2.7. Statistical Analysis
3. Results
3.1. Clinical Characteristics of the Study Cohort
3.2. CirculatingACE2 and TMPRSS2 Levels in Chronic Liver Disease Patients
3.3. Hepatic Expression of ACE2 and TMPRSS2
3.3.1. Western Blot Analysis
3.3.2. Immunohistochemistry
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sanyaolu, A.; Okorie, C.; Marinkovic, A.; Patidar, R.; Younis, K.; Desai, P.; Hosein, Z.; Padda, I.; Mangat, J.; Altaf, M. Comorbidity and its Impact on Patients with COVID-19. SN Compr. Clin. Med. 2020, 2, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Su, T.-H.; Kao, J.-H. The clinical manifestations and management of COVID-19-related liver injury. J. Formos. Med Assoc. 2020, 119, 1016–1018. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Shi, L.; Wang, F.-S. Liver injury in COVID-19: Management and challenges. Lancet Gastroenterol. Hepatol. 2020, 5, 428–430. [Google Scholar] [CrossRef]
- Chen, X.; Jiang, Q.; Ma, Z.; Ling, J.; Hu, W.; Cao, Q.; Mo, P.; Yao, L.; Yang, R.; Gao, S.; et al. Clinical Characteristics of Hospitalized Patients with SARS-CoV-2 and Hepatitis B Virus Co-infection. Virol. Sin. 2020, 35, 842–845. [Google Scholar] [CrossRef] [PubMed]
- Ji, D.; Qin, E.; Xu, J.; Zhang, D.; Cheng, G.; Wang, Y.; Lau, G. Non-alcoholic fatty liver diseases in patients with COVID-19: A retrospective study. J. Hepatol. 2020, 73, 451–453. [Google Scholar] [CrossRef] [PubMed]
- Mao, R.; Qiu, Y.; He, J.-S.; Tan, J.-Y.; Li, X.-H.; Liang, J.; Shen, J.; Zhu, L.-R.; Chen, Y.; Iacucci, M.; et al. Manifestations and prognosis of gastrointestinal and liver involvement in patients with COVID-19: A systematic review and meta-analysis. Lancet Gastroenterol. Hepatol. 2020, 5, 667–678. [Google Scholar] [CrossRef]
- Gupta, A.; Madhavan, M.V.; Sehgal, K.; Nair, N.; Mahajan, S.; Sehrawat, T.S.; Bikdeli, B.; Ahluwalia, N.; Ausiello, J.C.; Wan, E.Y.; et al. Extrapulmonary manifestations of COVID-19. Nat. Med. 2020, 26, 1017–1032. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e278. [Google Scholar] [CrossRef] [PubMed]
- Paizis, G.; Cooper, M.E.; Schembri, J.M.; Tikellis, C.; Burrell, L.M.; Angus, P.W. Up-regulation of components of the renin-angiotensin system in the bile duct–ligated rat liver. Gastroenterology 2002, 123, 1667–1676. [Google Scholar] [CrossRef] [PubMed]
- Rajapaksha, I.G.; Gunarathne, L.S.; Asadi, K.; Cunningham, S.C.; Sharland, A.; Alexander, I.E.; Angus, P.W.; Herath, C.B. Liver-Targeted Angiotensin Converting Enzyme 2 Therapy Inhibits Chronic Biliary Fibrosis in Multiple Drug-Resistant Gene 2-Knockout Mice. Hepatol. Commun. 2019, 3, 1656–1673. [Google Scholar] [CrossRef]
- Rajapaksha, I.G.; Gunarathne, L.S.; Asadi, K.; Laybutt, R.; Andrikopoulous, S.; Alexander, I.E.; Watt, M.J.; Angus, P.W.; Herath, C.B. Angiotensin Converting Enzyme-2 Therapy Improves Liver Fibrosis and Glycemic Control in Diabetic Mice With Fatty Liver. Hepatol. Commun. 2021, 6, 1056–1072. [Google Scholar] [CrossRef] [PubMed]
- Yeung, M.L.; Teng, J.L.L.; Jia, L.; Zhang, C.; Huang, C.; Cai, J.-P.; Zhou, R.; Chan, K.-H.; Zhao, H.; Zhu, L.; et al. Soluble ACE2-mediated cell entry of SARS-CoV-2 via interaction with proteins related to the renin-angiotensin system. Cell 2021, 184, 2212–2228.e12. [Google Scholar] [CrossRef]
- Wang, J.; Chen, S.; Bihl, J. Exosome-Mediated Transfer of ACE2 (Angiotensin-Converting Enzyme 2) from Endothelial Progenitor Cells Promotes Survival and Function of Endothelial Cell. Oxidative Med. Cell. Longev. 2020, 85, 4213541. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, P.A.; Pisitkun, T.; Hoffert, J.D.; Tchapyjnikov, D.; Star, R.A.; Kleta, R.; Wang, N.S.; Knepper, M.A. Large-Scale Proteomics and Phosphoproteomics of Urinary Exosomes. J. Am. Soc. Nephrol. 2008, 20, 363–379. [Google Scholar] [CrossRef] [PubMed]
- Raj, D.A.; Fiume, I.; Capasso, G.; Pocsfalvi, G. A multiplex quantitative proteomics strategy for protein biomarker studies in urinary exosomes. Kidney Int. 2012, 81, 1263–1272. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Ni, C.; Gao, R.; Wang, Y.; Yang, L.; Wei, J.; Lv, T.; Liang, J.; Zhang, Q.; Xu, W.; et al. Recapitulation of SARS-CoV-2 infection and cholangiocyte damage with human liver ductal organoids. Protein Cell 2020, 11, 771–775. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Han, Y.; Nilsson-Payant, B.E.; Gupta, V.; Wang, P.; Duan, X.; Tang, X.; Zhu, J.; Zhao, Z.; Jaffré, F.; et al. A Human Pluripotent Stem Cell-based Platform to Study SARS-CoV-2 Tropism and Model Virus Infection in Human Cells and Organoids. Cell Stem Cell 2020, 27, 125–136.e7. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Ballester, M.P.; Soffientini, U.; Jalan, R.; Mehta, G. SARS-CoV-2 infection and liver involvement. Hepatol. Int. 2022, 16, 755–774. [Google Scholar] [CrossRef] [PubMed]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.-C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Manco, M.; Ala, U.; Cantarella, D.; Tolosano, E.; Medico, E.; Altruda, F.; Fagoonee, S. The RNA-Binding Protein ESRP1 Modulates the Expression of RAC1b in Colorectal Cancer Cells. Cancers 2021, 13, 4092. [Google Scholar] [CrossRef] [PubMed]
- Fagoonee, S.; Arigoni, M.; Manco, M.; Olivero, M.; Bizzaro, F.; Magagnotti, C.; Andolfo, A.; Miniscalco, B.; Forni, M.; Todeschi, S.; et al. Circulating Extracellular Vesicles Contain Liver-Derived RNA Species as Indicators of Severe Cholestasis-Induced Early Liver Fibrosis in Mice. Antioxid. Redox Signal. 2022, 36, 480–504. [Google Scholar] [CrossRef] [PubMed]
- Famulari, E.S.; Navarro-Tableros, V.; Sanchez, M.B.H.; Bortolussi, G.; Gai, M.; Conti, L.; Silengo, L.; Tolosano, E.; Tetta, C.; Muro, A.F.; et al. Human liver stem cells express UGT1A1 and improve phenotype of immunocompromised Crigler Najjar syndrome type I mice. Sci. Rep. 2020, 10, 887. [Google Scholar] [CrossRef] [PubMed]
- Santana, M.F.; Guerra, M.T.; Hundt, M.A.; Ciarleglio, M.M.; Pinto, R.A.d.A.; Dutra, B.G.; Xavier, M.S.; Lacerda, M.V.G.; Ferreira, A.J.; Wanderley, D.C.; et al. Correlation Between Clinical and Pathological Findings of Liver Injury in 27 Patients with Lethal COVID-19 Infections in Brazil. Hepatol. Commun. 2021, 6, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Rosso, C.; Kazankov, K.; Younes, R.; Esmaili, S.; Marietti, M.; Sacco, M.; Carli, F.; Gaggini, M.; Salomone, F.; Møller, H.J.; et al. Crosstalk between adipose tissue insulin resistance and liver macrophages in non-alcoholic fatty liver disease. J. Hepatol. 2019, 71, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Popov, S.S.; Kryl’Skii, E.D.; Shulgin, K.K.; Raskina, E.A.; Popova, T.N.; Pashkov, A.N.; Kuptsova, G.N. Inflammation is associated with impairment of oxidative status, carbohydrate and lipid metabolism in type 2 diabetes complicated by non-alcoholic fatty liver disease. Minerva Endocrinol. 2022, 47, 304–313. [Google Scholar] [CrossRef]
- Schuppan, D.; Surabattula, R.; Wang, X.Y. Determinants of fibrosis progression and regression in NASH. J. Hepatol. 2018, 68, 238–250. [Google Scholar] [CrossRef]
- El-Shennawy, L.; Hoffmann, A.D.; Dashzeveg, N.K.; McAndrews, K.M.; Mehl, P.J.; Cornish, D.; Yu, Z.; Tokars, V.L.; Nicolaescu, V.; Tomatsidou, A.; et al. Circulating ACE2-expressing extracellular vesicles block broad strains of SARS-CoV-2. Nat. Commun. 2022, 13, 405. [Google Scholar] [CrossRef]
- Wu, D.; Zhu, H.; Wang, H. Extracellular Vesicles in Non-alcoholic Fatty Liver Disease and Alcoholic Liver Disease. Front. Physiol. 2021, 12, 707429. [Google Scholar] [CrossRef]
- Hirsova, P.; Ibrahim, S.H.; Krishnan, A.; Verma, V.K.; Bronk, S.F.; Werneburg, N.W.; Charlton, M.R.; Shah, V.H.; Malhi, H.; Gores, G.J. Lipid-Induced Signaling Causes Release of Inflammatory Extracellular Vesicles From Hepatocytes. Gastroenterology 2016, 150, 956–967. [Google Scholar] [CrossRef]
- Kakazu, E.; Mauer, A.S.; Yin, M.; Malhi, H. Hepatocytes release ceramide-enriched pro-inflammatory extracellular vesicles in an IRE1α-dependent manner. J. Lipid Res. 2016, 57, 233–245. [Google Scholar] [CrossRef]
- Ibrahim, S.H.; Hirsova, P.; Tomita, K.; Bronk, S.F.; Werneburg, N.W.; Harrison, S.A.; Goodfellow, V.S.; Malhi, H.; Gores, G.J. Mixed lineage kinase 3 mediates release of C-X-C motif ligand 10-bearing chemotactic extracellular vesicles from lipotoxic hepatocytes. Hepatology 2016, 63, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Paizis, G.; Tikellis, C.; Cooper, M.E.; Schembri, J.M.; Lew, R.A.; Smith, I.A.; Shaw, T.; Warner, F.J.; Zuilli, A.; Burrell, L.M.; et al. Chronic liver injury in rats and humans upregulates the novel enzyme angiotensin converting enzyme 2. Gut 2005, 54, 1790–1796. [Google Scholar] [CrossRef] [PubMed]
- Österreicher, C.H.; Taura, K.; De Minicis, S.; Seki, E.; Penz-Österreicher, M.; Kodama, Y.; Kluwe, J.; Schuster, M.; Oudit, G.Y.; Penninger, J.M.; et al. Angiotensin-converting-enzyme 2 inhibits liver fibrosis in mice. Hepatology 2009, 50, 929–938. [Google Scholar] [CrossRef] [PubMed]
- Fondevila, M.F.; Mercado-Gómez, M.; Rodríguez, A.; Gonzalez-Rellan, M.J.; Iruzubieta, P.; Valentí, V.; Escalada, J.; Schwaninger, M.; Prevot, V.; Dieguez, C.; et al. Obese patients with NASH have increased hepatic expression of SARS-CoV-2 critical entry points. J. Hepatol. 2020, 74, 469–471. [Google Scholar] [CrossRef] [PubMed]
- Biquard, L.; Valla, D.; Rautou, P.-E. No evidence for an increased liver uptake of SARS-CoV-2 in metabolic-associated fatty liver disease. J. Hepatol. 2020, 73, 718–719. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | All n = 42 | CHB n = 15 | CHC n = 12 | NAFLD n = 15 | p-Value |
|---|---|---|---|---|---|
| Age, years | 56 (53–62) | 56 (52–63) | 60 (54–67) | 53 (44–60) | 0.2847 |
| Male gender, n (%) | 25 (59.5%) | 9 (60%) | 5 (41.7%) | 11 (73.3%) | 0.2495 |
| BMI, kg/m2 | 26.8 (25.5–29.3) | 25.7 (24.3–26.8) | 24.9 (23.3–26.6) | 30.9 (29.6–32.0) | <0.0001 |
| Obesity, n (%) | 15 (35.7%) | 3 (20%) | 1 (8.3%) | 11 (73.3%) | 0.0006 |
| Type 2 diabetes, n (%) | 11 (26.2%) | 0 (-) | 2 (16.7%) | 9 (60%) | 0.0006 |
| AST, U/L | 25 (22–29) | 22 (20–27) | 24 (17–27) | 37 (26–47) | 0.0183 |
| ALT, U/L | 26 (20–37) | 23 (20–26) | 14 (12–25) | 55 (40–86) | <0.0001 |
| Platelets, ×109/L | 213 (200–251) | 200 (186–242) | 205 (166–244) | 273 (225–282) | 0.0279 |
| Variables | No Infection n = 29 | SARS-Cov-2 Infection, n = 13 | p-Value |
|---|---|---|---|
| Age, years | 53 (47–58) | 51 (39–65) | 0.9060 |
| Male gender, n (%) | 14 (48%) | 11 (85%) | 0.0284 |
| Etiology, n (%) CHB | 9 (31.1%) | 2 (15.4%) | 0.6672 |
| CHC | 7 (24.1%) | 5 (38.5%) | 0.6077 |
| NAFLD | 13 (44.8%) | 6 (46.1%) | 0.9589 |
| BMI, kg/m2 | 25.7 (24.5–27.5) | 29.4 (26.8–31.5) | 0.0702 |
| Obesity, n (%) | 8 (27.6%) | 7 (53.8%) | 0.1047 |
| Type 2 diabetes, n (%) | 60 (20.7%) | 5 (38.5%) | 0.0516 |
| AST, U/L | 26 (22–29) | 25 (18–41) | 0.7032 |
| ALT, U/L | 24 (20–39) | 35 (18–64) | 0.4540 |
| Platelets, ×109/L | 218 (199–258) | 209 (181–281) | 0.9566 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosso, C.; Demelas, C.; Agostini, G.; Abate, M.L.; Vernero, M.; Caviglia, G.P.; D’Amato, D.; Armandi, A.; Tapparo, M.; Guariglia, M.; et al. Expression of SARS-Cov-2 Entry Factors in Patients with Chronic Hepatitis. Viruses 2022, 14, 2397. https://doi.org/10.3390/v14112397
Rosso C, Demelas C, Agostini G, Abate ML, Vernero M, Caviglia GP, D’Amato D, Armandi A, Tapparo M, Guariglia M, et al. Expression of SARS-Cov-2 Entry Factors in Patients with Chronic Hepatitis. Viruses. 2022; 14(11):2397. https://doi.org/10.3390/v14112397
Chicago/Turabian StyleRosso, Chiara, Cristina Demelas, Greta Agostini, Maria Lorena Abate, Marta Vernero, Gian Paolo Caviglia, Daphne D’Amato, Angelo Armandi, Marta Tapparo, Marta Guariglia, and et al. 2022. "Expression of SARS-Cov-2 Entry Factors in Patients with Chronic Hepatitis" Viruses 14, no. 11: 2397. https://doi.org/10.3390/v14112397
APA StyleRosso, C., Demelas, C., Agostini, G., Abate, M. L., Vernero, M., Caviglia, G. P., D’Amato, D., Armandi, A., Tapparo, M., Guariglia, M., Troshina, G., Massano, A., Olivero, A., Nicolosi, A., Zannetti, A., Pellicano, R., Ciancio, A., Saracco, G. M., Ribaldone, D. G., ... Fagoonee, S. (2022). Expression of SARS-Cov-2 Entry Factors in Patients with Chronic Hepatitis. Viruses, 14(11), 2397. https://doi.org/10.3390/v14112397