Abstract
Viral proteins interact with different sets of host cell components throughout the viral life cycle and are known to localize to the intracellular membraneless organelles (MLOs) of the host cell, where formation/dissolution is regulated by phase separation of intrinsically disordered proteins and regions (IDPs/IDRs). Viral proteins are rich in IDRs, implying that viruses utilize IDRs to regulate phase separation of the host cell organelles and augment replication by commandeering the functions of the organelles and/or sneaking into the organelles to evade the host immune response. This review aims to integrate current knowledge of the structural properties and intracellular localizations of viral IDPs to understand viral strategies in the host cell. First, the properties of viral IDRs are reviewed and similarities and differences with those of eukaryotes are described. The higher IDR content in viruses with smaller genomes suggests that IDRs are essential characteristics of viral proteins. Then, the interactions of the IDRs of flaviviruses with the MLOs of the host cell are investigated with emphasis on the viral proteins localized in the nucleoli and stress granules. Finally, the possible roles of viral IDRs in regulation of the phase separation of organelles and future possibilities for antiviral drug development are discussed.
1. Introduction
Viral proteins interact with different sets of host cellular components throughout the viral life cycle, beginning with entry, proceeding to replication, virion assembly, and finally exiting from the host cell surface. Since this series of events occurs in different intracellular compartments, the intracellular localization of viral proteins and genomes also plays pivotal roles in the viral life cycle. There have been a number of reports on the unique intracellular localization of viral proteins in membraneless organelles (MLOs), such as the nucleoli, stress granules (SGs), P-bodies, and other RNA-containing granules, which sometimes differ from the sites of replication and virion assembly. The formation of MLOs is mostly driven by liquid–liquid phase separation (LLPS) or condensation of disordered polypeptides (in some cases together with RNA), in which highly charged polypeptides interact with each other via promiscuous interactions to self-assemble into separated phases. In contrast to conventional “stereospecific” interactions between the solid surfaces of structured proteins, such promiscuous interactions allow for dynamic assembly/disassembly of macromolecules in the intracellular milieu and flexible responses to environmental changes. In the case of LLPS, confinement of molecules (especially enzymes and substrates) into a limited volume in the vast cytoplasmic space is considered to increase the rates and efficiency of multi-step reactions.
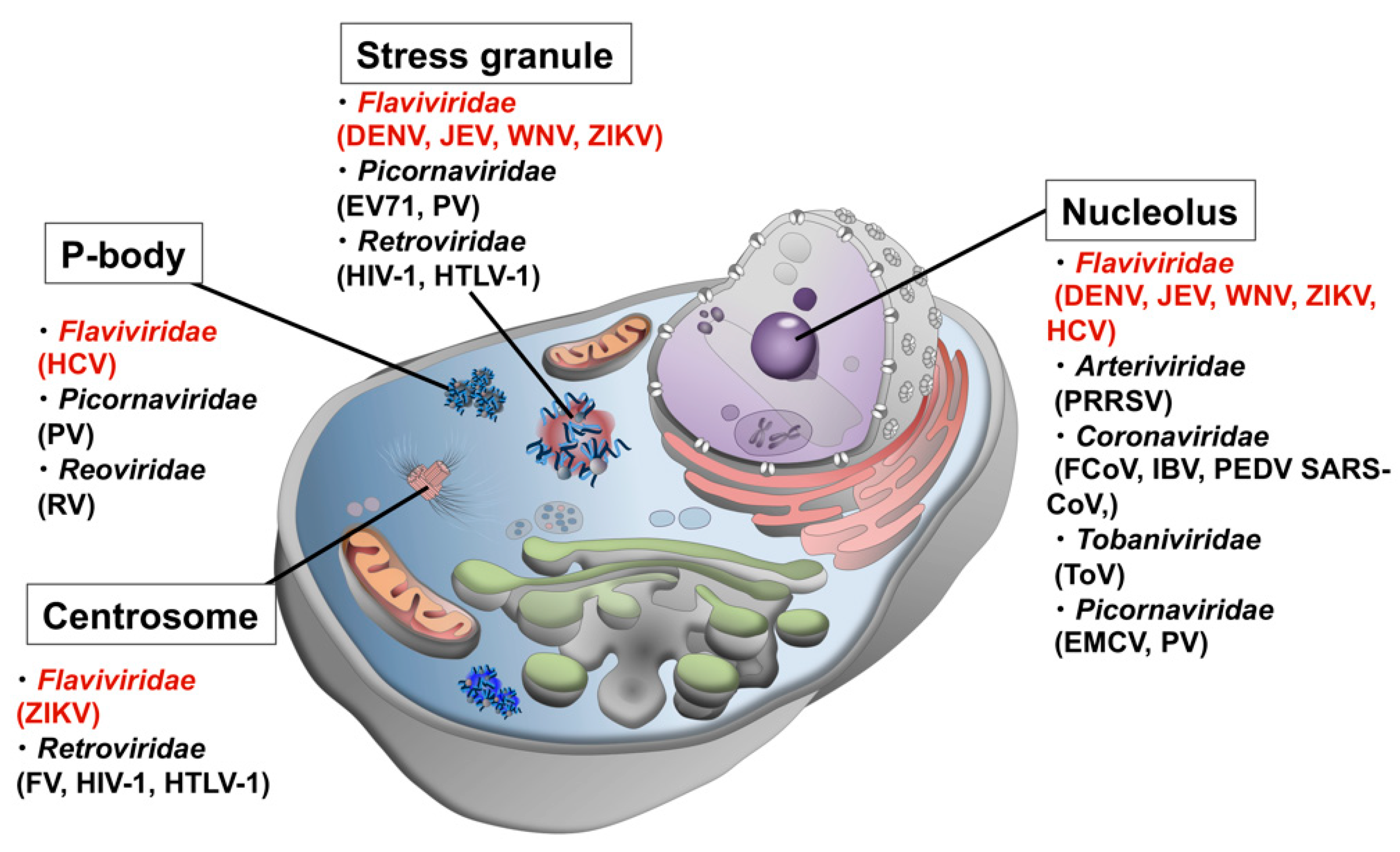
Another line of evidence from genomic and proteomic analyses using bioinformatics approaches revealed that viral proteins have high contents of disordered regions [1,2,3], which was initially interpreted as a viral strategy to efficiently use the limited genomic information by conferring multi-functionality and adaptability to a single protein. However, in the recent context of the role of disordered proteins/regions in LLPS and the formation of MLOs, it is highly probable that disordered viral proteins interact with host MLOs to either commandeer the function of the organelle to promote efficient viral replication and proliferation, or to sneak into the organelle to avoid the host cell immune response. To date, dozens of viral proteins have been found localized in MLOs of host cells (Figure 1 and Table 1).
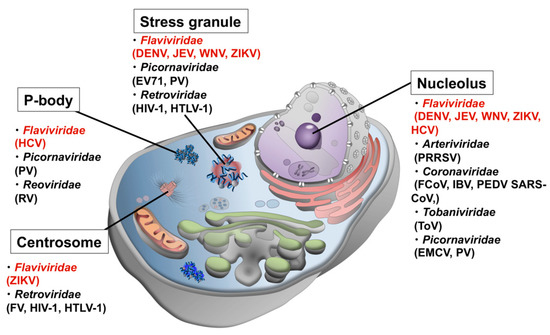
Figure 1.
Possible interactions between host MLOs and viral proteins. Viruses in Flaviviridae are shown in red.

Table 1.
Interactions of viral proteins with MLOs.
The 70 viruses in the genus Flavivirus (family Flaviviridae), including several arthropod-borne human pathogens such as dengue virus (DENV), West Nile virus (WNV), yellow fever virus (YFV), Zika virus (ZIKV), and Japanese encephalitis virus (JEV), pose major threats to human health, mainly in tropical and subtropical countries as many of them are transmitted by mosquitoes. The clinical manifestation of flavivirus infection ranges from asymptomatic cases, mild symptoms, to life-threatening hemorrhagic fever. For example, infection with neurptropic viruses JEV and WNV can lead to death [24,25], and ZIKV can cause microcephaly in infants, as seen in the ZIKV epidemic in South America from 2015 to 2016 that resulted in a high number of confirmed cases [26]. Current situation of vaccine development targeting flaviviruses is summarized in Box 1.
Box 1. Current situation of vaccine development targeting flaviviruses.
There are currently no approved modalities for the treatment of flaviviral infections. Additionally, in case of secondary DENV infection with a different serotype of DENV, pre-existing antibodies (Abs) against DENV augments infection via antibody-dependent enhancement (ADE) [27,28]. Pre-existing Abs form complexes with viral particles, which bind to monocytes/macrophages via Fc receptors on the cell surface, resulting in enhanced infections in these cell types. In fact, most severe DENV cases are associated with secondary infection with a heterologous serotype of DENV [27,28]. Also, pre-existing Abs targeting DENV can augment the infection of ZIKV [29,30,31]. While vaccines targeting YFV and JEV are available, this complication of ADE makes it difficult to develop effective vaccines against infections with DENV and ZIKV. Therefore, the development of antiviral drugs targeting the “Achilles’ heel” of flaviviruses is strongly desired.
Flavivirus assembly follows the formation of replication complexes on the endoplasmic reticulum (ER) that consist of seven non-structural proteins and genomic RNA. The flavivirus genome is an approximately 11 kb, single-stranded, positive-sense RNA molecule encoding three structural proteins (core, prM, and envelope) and seven non-structural (NS) proteins. These viral proteins are translated from a single genomic RNA as a polyprotein, which is then cleaved by cellular proteases, viral proteases, and a signal peptidase. Thus, flavivirus replication depends on cellular enzymes and host intracellular organelles, such as secretary vesicles, the Golgi apparatus, and ER. Several lines of evidence suggest that flavivirus infection induces rearrangements of the ER membrane to generate vesicle packets that optimize RNA replication and virion assembly, shielding the viral components from the host immune response, such as interferon-mediated inhibition [32,33,34,35,36,37].
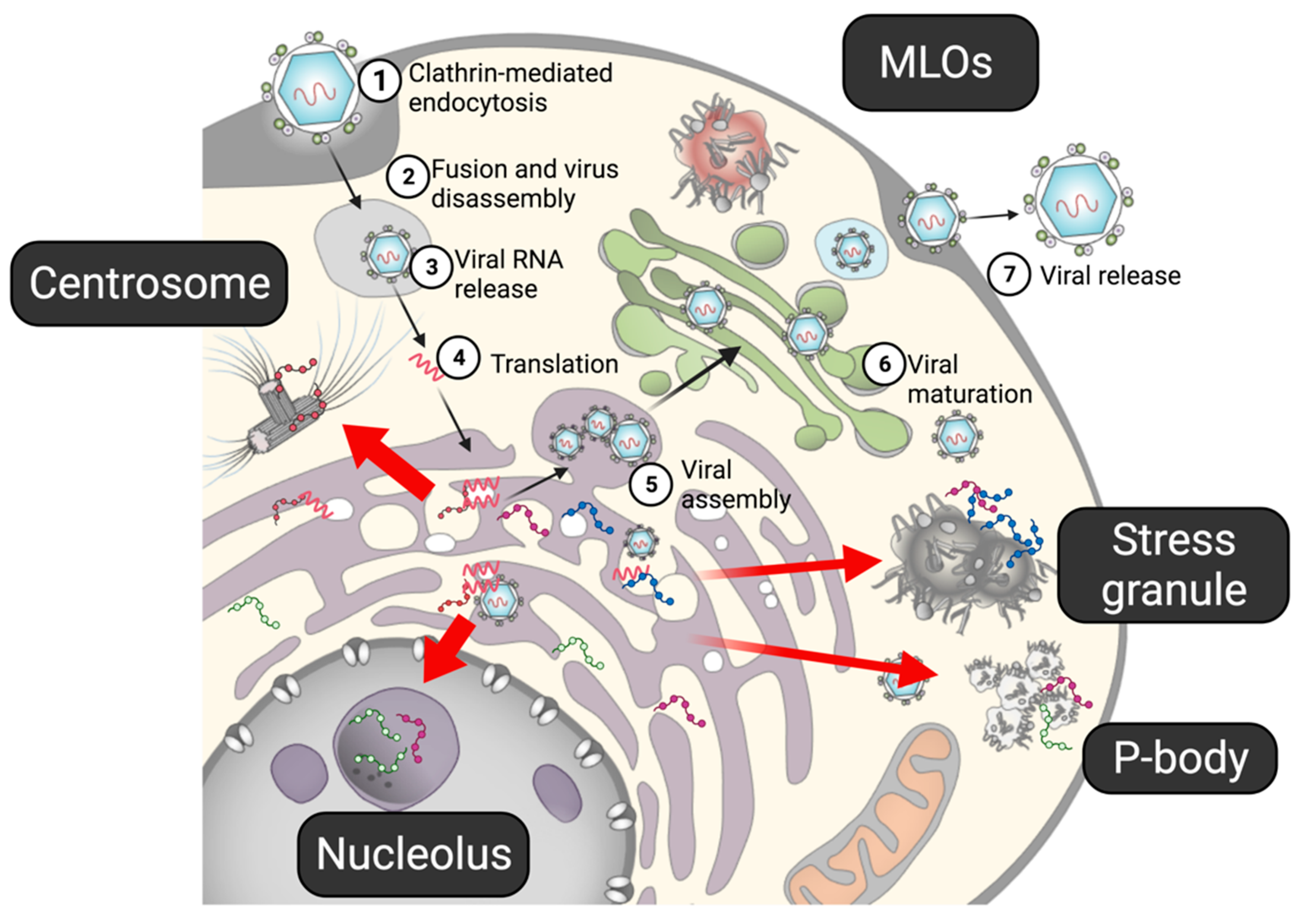
In addition to the vesicle pocket, flaviviral proteins are known to localize in MLOs, such as the core protein and NS5 in nucleoli, and the core and some NSs proteins in stress granules (SGs) (Figure 2). Mutations in the core protein abolish the protein’s nucleolar localization and reduce the infectivity of the virus [38], suggesting that the nucleolar localization of the core protein plays a critical role in viral replication. Nucleolar localization of the core protein is also observed in other flaviviruses, including WNV, DENV, and ZIKV [4], indicating a general function of the core protein in the nucleoli. Because many flaviviral proteins, including the core protein, have IDRs, they may be involved in LLPS. In this article, the properties of the disordered viral proteins obtained from genomic and proteomic analyses are reviewed. In addition, the potential interplay between the flaviviral proteins and MLOs, e.g., nucleoli and SGs, in the viral life cycle are discussed.
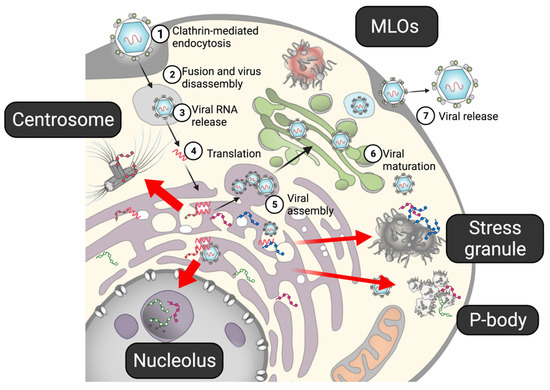
Figure 2.
The life cycle of flavivirus. (1) After a flavivirus attaches to a specific receptor on a cell, it enters the cell via clathrin-mediated endocytosis. (2,3) Under the low-pH condition in the endosome, the viral capsid is disassembled, and viral RNA is released into the cell. (4) Viral RNA, acting as mRNA, is directly translated into viral proteins in the ER. The non-structural proteins form a replication complex to propagate viral RNA, while structural proteins form viral particles. (5) Viral assembly also occurs in the ER. (6) The prM on the immature viral particles is cleaved by furin in the Golgi apparatus. (7) Mature virions are released from the cell. Several viral proteins have been shown to interact with the MLOs. Flaviviral proteins have been shown to interact with MLOs (red arrows).
2. Disordered Regions in Viral Proteins
2.1. Viral Proteins Are Rich in Disordered Regions
An intrinsically disordered region (IDR), also referred to as intrinsically disordered protein (IDP), is a protein region with no particular secondary or tertiary structure. The lack of a rigid three-dimensional structure allows an IDR to interact with several partners simultaneously. IDRs possess a high content of charged residues and a low content of hydrophobic residues. The propensity for disorder of a given protein or peptide can be estimated from the amino acid sequence with the use of several algorithms, including IUPred (http://iupred.enzim.hu/index.html [39]), DisProt (https://disprot.org/ [40]), and PONDR (http://www.pondr.com/ [41]) (all of these sites were accessed on 21 July 2021). Bioinformatic analyses of genomic sequences from different kingdoms (i.e., bacteria, archaea, and eukaryotes) using these algorithms revealed relationships between the length and frequency of IDRs and complexity of the organism [42,43,44], as 7–30% of prokaryotic proteins and 45–50% of eukaryotic proteins contain disordered regions with >30 consecutive residues [42,43,45].
Quantification of IDRs in various viral proteomes revealed that viruses have the largest range of variation in the content of disordered residues ranging from 7.3% in human coronavirus NL63 to 77.3% in avian carcinoma virus [3]. Interestingly, there is a reverse relationship between the proteome size and the IDR content. The proteomes of small viruses with five or less proteins have 50% or more disordered residues: the greatest number of all species [3]. As the proteome size increases, the IDR content decreases to 20–40%. Another informatics study of the genome of 2278 viruses in 41 families revealed that the number of IDRs was correlated with the genome size [46]. Viral families with smaller genomes have higher contents of IDRs within five of the six main viral types (ssDNA, dsDNA, ssRNA(+), dsRNA, and retroviruses), with the exception of ssRNA(−), in which the proportion of IDRs increases with the genome size. This relationship between the proteome/genome size and the IDR content shows a clear contrast to that in eukaryotes, which have an IDR content as high as 40–50%, regardless of the proteome/genome size. The high IDR content of small viral proteomes seems to support the idea that viruses utilize IDRs/IDPs to increase flexibility and/or multi-functionality of individual proteins and gain broader adaptability to environmental changes from small amounts of genomic information, indicating that a high IDR content enables efficient usage of genomic information. Alternatively, IDR may play a critical role in the function of a minimum protein component that is necessary for virion assembly, but less in other accessory proteins. More structural information on viral proteins and bioinformatics analyses are required to test these possibilities.
2.2. Advantages of High IDR Content in Viral Strategies
The high IDR content of viral proteins implies that IDR is a general and essential strategy for efficient viral replication. There are several possible explanations for this. First, the lack of any particular three-dimensional structure allows for interactions with more than one partner molecule. In contrast to the stereospecific interactions between/among the solid surfaces of structured proteins, an IDR can interact with multiple solid surfaces with different structures (induced fit). In addition to this order–disorder interface, disorder–disorder interactions are also possible. Moreover, promiscuous and multivalent interactions can occur within a single type of IDR (if the polypeptide is polyampholyte), as well as among several different types of IDRs (especially between IDRs with opposite net charges). In some cases, such interactions induce LLPS (see later section). An IDR can also expand the variety of interactions by connecting two structured (functional) domains as a flexible linker, so as to increase the accessible surface area of the structured domain. These multi-interaction mechanisms mediated by IDRs confer functional advantages to viral proteins, which must interact with various host molecules, including proteins, membrane components, and nucleic acids, at different steps of the entire life cycle.
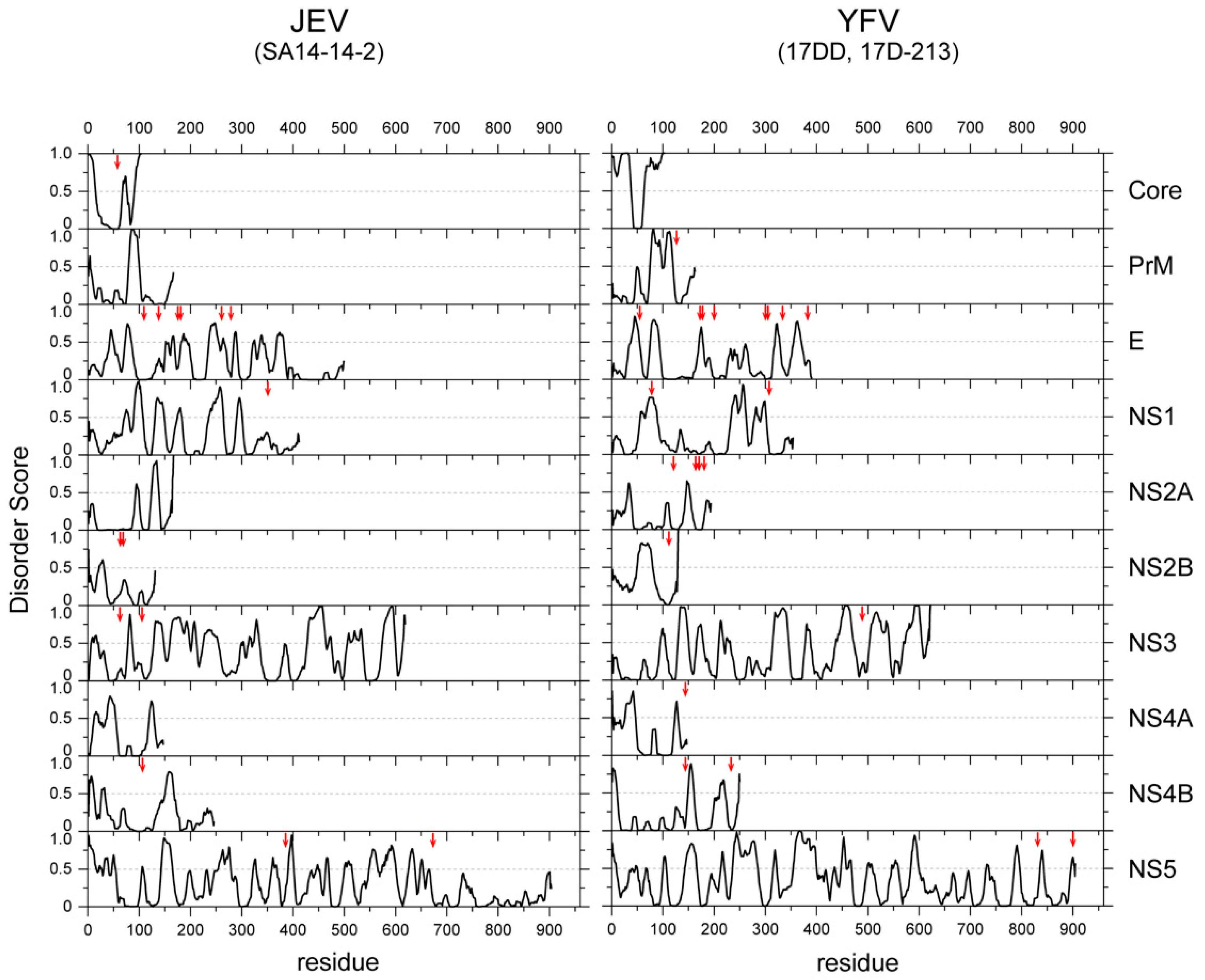
Another advantage of the viral IDR is the high mutation rate. The lack of structural constraints may help to tolerate high mutation rates and promote faster adaptation to environmental changes. Viruses must survive both within the host cells and the host environments (extracellular spaces) by adapting to a variety of environmental factors, including antiviral drugs and the host immune response. Generally, the viral genome exhibits an extremely higher mutation rate (10−5–10−3 for RNA viruses and 10−8–10−5 for DNA viruses per position per generation) than the bacterial and eukaryotic genomes (10−9 on average) [47]. Although the accuracy of polymerases and the replication machinery is the major driving force of such high mutation rates, the structural constraints of translated polypeptides also contribute to this. Furthermore, overlapping reading frames, which can be found in many viral genomes, also accelerate the evolution of viral proteins. It is intriguing that mutations to flaviviral proteins, which abolish viral infectivity, are mostly found in structured domains, rather than the IDRs (Figure 3), implying that IDRs are more tolerant to mutations than structured domains.
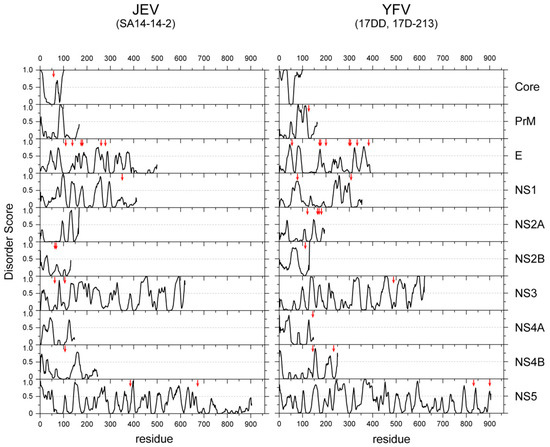
Figure 3.
Amino acid substitutions occurred predominantly in the structured regions of the viral proteins of a vaccine strain. The IDR scores of all the proteins of JEV (left) and YFV (right) are plotted. The positions of the amino acid mutations found in the vaccine strains (SA14-14-2 for JEV [48] and 17D and 17DD-21-3 of YFV [49]) are indicated with vertical red arrows.
2.3. RNA Chaperone Activity of Viral IDRs
RNA chaperoning activity is one of the distinct and unique functions of IDRs. RNA chaperones exist in various cell types, including prokaryotes and eukaryotes, and play pivotal roles in many biological processes, such as transcription, mRNA export, splicing, and ribosome biogenesis. An RNA chaperone binds to, partially unfolds, and finally refolds misfolded RNA into a more stable conformation. Because RNA is prone to post-transcriptional misfolding [50], chaperone activity is necessary for proper folding and function of cellular RNA. Many RNA viruses and retroviruses also have proteins with RNA chaperone activity. For example, the nucleocapsid protein NCp7 of human immunodeficiency virus type I (HIV-1) has RNA chaperone activity, which is necessary for maturation, reverse transcription, and integration [51,52,53] (see later section).
The results of a proteomic study demonstrated that known RNA chaperones, as well as protein chaperones, contain high percentages of IDRs (highest of all proteins groups) [54]. In the case of RNA chaperones, promiscuous interactions between basic residues within the IDR and RNA promotes annealing of two complementary sequences or refolding of misfolded RNA molecules [55]. Although the molecular mechanism of the chaperone activity of IDRs remains unclear, Tompa et al. proposed an “entropy exchange model,” where the partial order–disorder transition of a chaperone IDR is coupled to the unfolding–refolding conversion of the substrate to a more stable conformation [54]. This shows a clear contrast to the typical catalytic activities of structured enzymes of stereospecific interactions between the catalytic pocket of the enzyme and the substrate, which are critical for the reaction. Notably, the first protein enzyme appearing in the RNA world could have been an RNA chaperone [56].
The RNA chaperones of RNA viruses and retroviruses also contain significant numbers of IDRs. For example, the NCp7 protein of HIV-1, a short polypeptide of 55 amino acids, is derived from the Gag polyprotein and contains two CCHC-type zinc fingers [57]. Disorder prediction, as well as NMR solution structure, revealed that zinc fingers have relatively defined structures, although other flanking sequences are disordered in the absence of RNA [58,59,60]. The nucleocapsid protein recognizes packaging sequences within the viral RNA genome and promotes efficient packaging into a virion. Due to its basic amino acid composition, nucleocapsid protein binds to RNA in a sequence-independent manner and functions as an RNA chaperone to promote several RNA-dependent steps in the HIV life cycle, such as annealing of the tRNA primer, stimulating integration [61], and promoting DNA strand exchange reactions during reverse transcription [62,63,64]. The functional roles of IDRs of other HIV-1 accessory proteins are reviewed in a previous article [60].
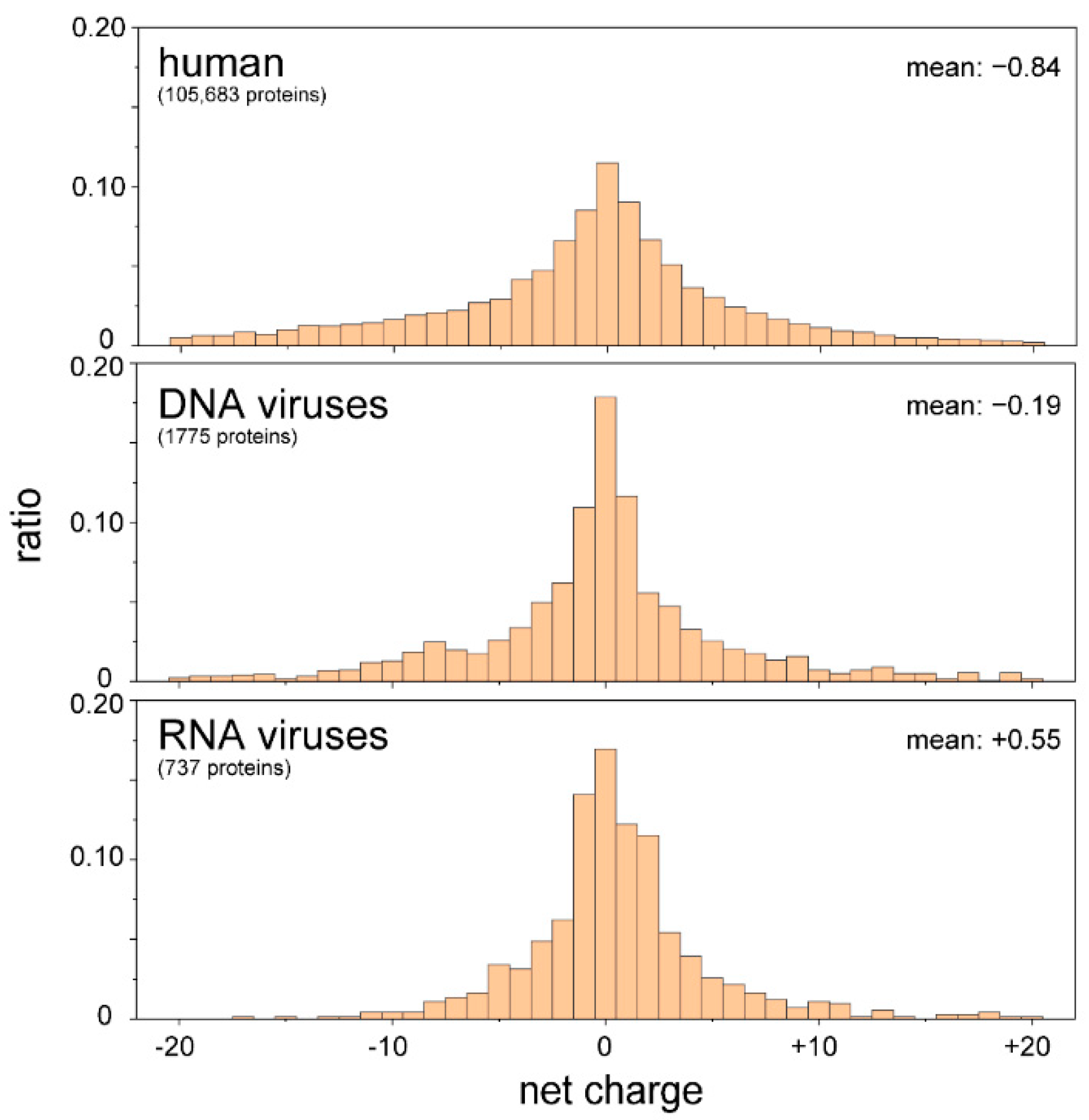
The core proteins of viruses in the family Flaviviridae (i.e., hepatitis C virus (HCV), GB virus B, West Nile virus (WNV), and bovine viral diarrhea virus (BVDV)-1) are also required for packaging the RNA genome and protecting it from the host immune response. The N-terminal IDR has RNA chaperone activity [65,66,67,68]. It can be speculated that RNA viruses use the IDRs to control the conformation of the RNA genome to facilitate replication, reverse transcription (in the case of retroviruses), and packing. Our proteomic analyses demonstrated that the IDRs of RNA viruses are rich in basic amino acids, whereas human IDRs and those of DNA viruses are slightly negative (Figure 4). These results imply that RNA viruses use basic IDRs to interact with and promote proper folding of genomic RNA in the host cell. In the following section, possible roles of IDRs/IDPs in the life cycles of flaviviruses are discussed.
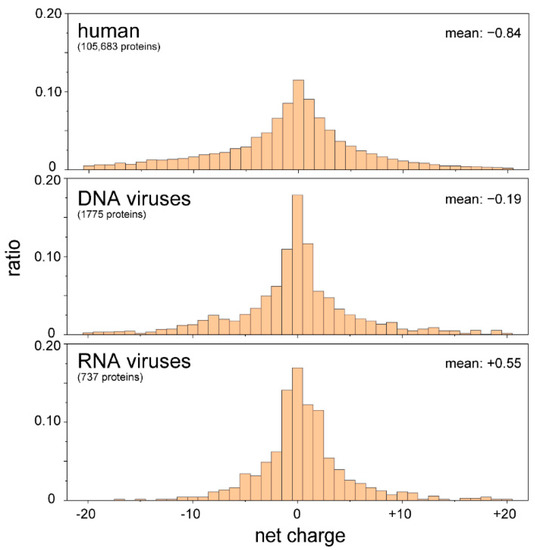
Figure 4.
IDRs of RNA viruses are positively charged. The amino acid sequences of IDRs were extracted from proteomic databases of the human, DNA viruses, and RNA viruses. The net charges of the IDRs of individual proteins were calculated and are summarized as a histogram. IDRs carrying more than 21 net charges (either negative or positive) were omitted. There were significant differences between human and RNA viruses, as well as DNA and RNA viruses, with p-values of less than 0.01 (t-test).
4. Closing Remarks and Future Directions
In this review, the basic characteristics of IDR as well as the complex interplay between viral proteins and MLOs are summarized. There must be selective pressure for RNA viruses to maintain and/or evolve the IDRs of viral proteins to optimize replication, adapt to new host species, and avoid stimulation of host immune sensors. Recent advances in research tools and understanding of LLPS has allowed for the bridging of the traditional fields of biophysics and virology, thereby creating a new field to understand the complex and sophisticated events during viral replication.
Recent studies have reported that the nucleoprotein of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) induces LLPS in the presence of viral RNA [125,126]. The nucleoprotein of SARS-CoV-2 co-localizes with G3BP, a marker of SGs. Once phosphorylated, the serine/arginine-rich region of the nucleoprotein interacts with RNA. LLPS of the nucleoprotein and RNA can recruit non-structural protein (NSP) 7, NSP8, and NSP12, suggesting that LLPS of the nucleoprotein is required for the condensation of the viral replication machinery [127]. Importantly, the (−)-gallocatechin gallate (GCG), a polyphenol from green tea, was found to disrupt the interaction between the nucleoprotein and RNA and inhibit the formation of LLPS of the nucleoprotein [128]. Treatment with GCG suppressed viral RNA replication of SARS-CoV-2, suggesting that the LLPS of SARS-CoV-2 can be a therapeutic target for COVID-19 [128]. In addition, the nucleoprotein and phosphoprotein of measles virus (MeV) are also reported to form MLOs through LLPS, which are then co-localized with RNA and contribute to efficient viral replication [129]. These data imply that MLOs formed by the viral replication machinery might be constructed by LLPS, similar to MeV [130]. The targeting of LLPS, as shown by GCG of SARS-CoV-2, has potential as an antiviral strategy, although further investigations are needed to determine the importance of LLPS on the life cycles of other RNA viruses and to develop specific inhibitors against viral LLPS.
Author Contributions
A.S., M.S., M.Y., J.H., T.O., and S.H.Y. wrote the review. H.O. and S.H.Y. analyzed the IDR regions of viral and human proteins. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported in part by the AMED Japan Program for Infectious Diseases Research and Infrastructure (grant nos. 20wm0325009 and 21wm0325009 to A.S., T.O., and S.H.Y.); AMED Research Program on Emerging and Re-emerging Infectious Diseases (grant no. 20fk0108163 to A.S. and T.O.; grant no. 20fk0108518 to A.S. and T.O.); JSPS KAKENHI Grant-in-Aid for Scientific Research (C) (grant no. 19K06382 to A.S.) and Grant-in-Aid for Scientific Research on Innovative Areas (grant no.19H04830 to S.H.Y.); and the Joint Research Project of the Research Institute for Microbial Diseases, Osaka University (to A.S.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Intrinsic disordered residues were predicted with the IUPred2A program, by using the short disorder option (https://academic.oup.com/nar/article/46/W1/W329/5026265). The residues with IUPred score >0.5 were regarded as disordered residues. The protein sequences examined and their metadata were obtained from the NCBI virus database (https://www.ncbi.nlm.nih.gov/labs/virus/vssi/#/) and the NCBI protein database (https://www.ncbi.nlm.nih.gov/protein/) (all of these sites were accessed on 3 May 2021). We here focused on the REFSEQ sequences of proteins for viruses whose hosts are human and human proteins.
Acknowledgments
The authors thank Tomoko Nishiuchi and Tamaki Okabayashi for their help.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tokuriki, N.; Oldfield, C.J.; Uversky, V.N.; Berezovsky, I.N.; Tawfik, D.S. Do Viral Proteins Possess Unique Biophysical Features? Trends Biochem. Sci. 2009, 34, 53–59. [Google Scholar] [CrossRef]
- Xue, B.; Williams, R.W.; Oldfield, C.J.; Goh, G.K.-M.; Dunker, A.K.; Uversky, V.N. Viral Disorder or Disordered Viruses: Do Viral Proteins Possess Unique Features? Protein Pept. Lett. 2010, 17, 932–951. [Google Scholar] [CrossRef]
- Xue, B.; Dunker, A.K.; Uversky, V.N. Orderly Order in Protein Intrinsic Disorder Distribution: Disorder in 3500 Proteomes from Viruses and the Three Domains of Life. J. Biomol. Struct. Dyn. 2012, 30, 137–149. [Google Scholar] [CrossRef]
- Tokunaga, M.; Miyamoto, Y.; Suzuki, T.; Otani, M.; Inuki, S.; Esaki, T.; Nagao, C.; Mizuguchi, K.; Ohno, H.; Yoneda, Y.; et al. Novel Anti-Flavivirus Drugs Targeting the Nucleolar Distribution of Core Protein. Virology 2020, 541, 41–51. [Google Scholar] [CrossRef]
- Fraser, J.E.; Rawlinson, S.M.; Heaton, S.M.; Jans, D.A. Dynamic Nucleolar Targeting of Dengue Virus Polymerase NS5 in Response to Extracellular PH. J. Virol. 2016, 90, 5797–5807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, Z.; Pan, T.; Wu, X.; Song, W.; Wang, S.; Xu, Y.; Rice, C.M.; MacDonald, M.R.; Yuan, Z. Hepatitis C Virus Co-Opts Ras-GTPase-Activating Protein-Binding Protein 1 for Its Genome Replication. J. Virol. 2011, 85, 6996–7004. [Google Scholar] [CrossRef] [Green Version]
- Hiscox, J.A.; Wurm, T.; Wilson, L.; Britton, P.; Cavanagh, D.; Brooks, G. The Coronavirus Infectious Bronchitis Virus Nucleoprotein Localizes to the Nucleolus. J. Virol. 2001, 75, 506–512. [Google Scholar] [CrossRef] [Green Version]
- You, J.-H.; Reed, M.L.; Hiscox, J.A. Trafficking Motifs in the SARS-Coronavirus Nucleocapsid Protein. Biochem. Biophys. Res. Commun. 2007, 358, 1015–1020. [Google Scholar] [CrossRef]
- Acar, D.D.; Stroobants, V.J.E.; Favoreel, H.; Saelens, X.; Nauwynck, H.J.Y. 2019 Identification of Peptide Domains Involved in the Subcellular Localization of the Feline Coronavirus 3b Protein. J. Gen. Virol. 2019, 100, 1417–1430. [Google Scholar] [CrossRef]
- Yuan, X.; Yao, Z.; Shan, Y.; Chen, B.; Yang, Z.; Wu, J.; Zhao, Z.; Chen, J.; Cong, Y. Nucleolar Localization of Non-Structural Protein 3b, a Protein Specifically Encoded by the Severe Acute Respiratory Syndrome Coronavirus. Virus Res. 2005, 114, 70–79. [Google Scholar] [CrossRef]
- Aminev, A.G.; Amineva, S.P.; Palmenberg, A.C. Encephalomyocarditis Viral Protein 2A Localizes to Nucleoli and Inhibits Cap-Dependent MRNA Translation. Virus Res. 2003, 95, 45–57. [Google Scholar] [CrossRef]
- Katoh, H.; Okamoto, T.; Fukuhara, T.; Kambara, H.; Morita, E.; Mori, Y.; Kamitani, W.; Matsuura, Y. Japanese Encephalitis Virus Core Protein Inhibits Stress Granule Formation through an Interaction with Caprin-1 and Facilitates Viral Propagation. J. Virol. 2013, 87, 489–502. [Google Scholar] [CrossRef] [Green Version]
- Emara, M.M.; Brinton, M.A. Interaction of TIA-1/TIAR with West Nile and Dengue Virus Products in Infected Cells Interferes with Stress Granule Formation and Processing Body Assembly. Proc. Natl. Acad. Sci. USA 2007, 104, 9041–9046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Hu, Z.; Fan, S.; Zhang, Q.; Zhong, Y.; Guo, D.; Qin, Y.; Chen, M. Picornavirus 2A Protease Regulates Stress Granule Formation to Facilitate Viral Translation. PLoS Pathog. 2018, 14, e1006901. [Google Scholar] [CrossRef]
- Wu, S.; Wang, Y.; Lin, L.; Si, X.; Wang, T.; Zhong, X.; Tong, L.; Luan, Y.; Chen, Y.; Li, X.; et al. Protease 2A Induces Stress Granule Formation during Coxsackievirus B3 and Enterovirus 71 Infections. Virol. J. 2014, 11, 192. [Google Scholar] [CrossRef] [Green Version]
- Abrahamyan, L.G.; Chatel-Chaix, L.; Ajamian, L.; Milev, M.P.; Monette, A.; Clément, J.-F.; Song, R.; Lehmann, M.; DesGroseillers, L.; Laughrea, M.; et al. Novel Staufen1 Ribonucleoproteins Prevent Formation of Stress Granules but Favour Encapsidation of HIV-1 Genomic RNA. J. Cell Sci. 2010, 123, 369–383. [Google Scholar] [CrossRef] [Green Version]
- Legros, S.; Boxus, M.; Gatot, J.S.; Van Lint, C.; Kruys, V.; Kettmann, R.; Twizere, J.C.; Dequiedt, F. The HTLV-1 Tax Protein Inhibits Formation of Stress Granules by Interacting with Histone Deacetylase 6. Oncogene 2011, 30, 4050–4062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Ndongwe, T.P.; Puray-Chavez, M.; Casey, M.C.; Izumi, T.; Pathak, V.K.; Tedbury, P.R.; Sarafianos, S.G. Effect of P-Body Component Mov10 on HCV Virus Production and Infectivity. FASEB J. 2020, 34, 9433–9449. [Google Scholar] [CrossRef] [PubMed]
- Bhowmick, R.; Mukherjee, A.; Patra, U.; Chawla-Sarkar, M. Rotavirus Disrupts Cytoplasmic P Bodies during Infection. Virus Res. 2015, 210, 344–354. [Google Scholar] [CrossRef]
- Dougherty, J.D.; White, J.P.; Lloyd, R.E. Poliovirus-Mediated Disruption of Cytoplasmic Processing Bodies. J. Virol. 2011, 85, 64–75. [Google Scholar] [CrossRef] [Green Version]
- Kesari, A.S.; Heintz, V.J.; Poudyal, S.; Miller, A.S.; Kuhn, R.J.; LaCount, D.J. Zika Virus NS5 Localizes at Centrosomes during Cell Division. Virology 2020, 541, 52–62. [Google Scholar] [CrossRef]
- Afonso, P.V.; Zamborlini, A.; Saïb, A.; Mahieux, R. Centrosome and Retroviruses: The Dangerous Liaisons. Retrovirology 2007, 4, 27. [Google Scholar] [CrossRef] [Green Version]
- Peloponese, J.-M.; Haller, K.; Miyazato, A.; Jeang, K.-T. Abnormal Centrosome Amplification in Cells through the Targeting of Ran-Binding Protein-1 by the Human T Cell Leukemia Virus Type-1 Tax Oncoprotein. Proc. Natl. Acad. Sci. USA 2005, 102, 18974–18979. [Google Scholar] [CrossRef] [Green Version]
- Davis, L.E.; DeBiasi, R.; Goade, D.E.; Haaland, K.Y.; Harrington, J.A.; Harnar, J.B.; Pergam, S.A.; King, M.K.; DeMasters, B.K.; Tyler, K.L. West Nile Virus Neuroinvasive Disease. Ann. Neurol. 2006, 60, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Misra, U.K.; Kalita, J. Overview: Japanese Encephalitis. Prog. Neurobiol. 2010, 91, 108–120. [Google Scholar] [CrossRef]
- Weaver, S.C.; Costa, F.; Garcia-Blanco, M.A.; Ko, A.I.; Ribeiro, G.S.; Saade, G.; Shi, P.-Y.; Vasilakis, N. Zika Virus: History, Emergence, Biology, and Prospects for Control. Antivir. Res. 2016, 130, 69–80. [Google Scholar] [CrossRef]
- Katzelnick, L.C.; Gresh, L.; Halloran, M.E.; Mercado, J.C.; Kuan, G.; Gordon, A.; Balmaseda, A.; Harris, E. Antibody-Dependent Enhancement of Severe Dengue Disease in Humans. Science 2017, 358, 929–932. [Google Scholar] [CrossRef] [Green Version]
- Dejnirattisai, W.; Jumnainsong, A.; Onsirisakul, N.; Fitton, P.; Vasanawathana, S.; Limpitikul, W.; Puttikhunt, C.; Edwards, C.; Duangchinda, T.; Supasa, S.; et al. Cross-Reacting Antibodies Enhance Dengue Virus Infection in Humans. Science 2010, 328, 745–748. [Google Scholar] [CrossRef] [Green Version]
- Bardina, S.V.; Bunduc, P.; Tripathi, S.; Duehr, J.; Frere, J.J.; Brown, J.A.; Nachbagauer, R.; Foster, G.A.; Krysztof, D.; Tortorella, D.; et al. Enhancement of Zika Virus Pathogenesis by Preexisting Antiflavivirus Immunity. Science 2017, 356, 175–180. [Google Scholar] [CrossRef] [Green Version]
- Paul, L.M.; Carlin, E.R.; Jenkins, M.M.; Tan, A.L.; Barcellona, C.M.; Nicholson, C.O.; Michael, S.F.; Isern, S. Dengue Virus Antibodies Enhance Zika Virus Infection. Clin. Transl. Immunol. 2016, 5, e117. [Google Scholar] [CrossRef]
- Dejnirattisai, W.; Supasa, P.; Wongwiwat, W.; Rouvinski, A.; Barba-Spaeth, G.; Duangchinda, T.; Sakuntabhai, A.; Cao-Lormeau, V.-M.; Malasit, P.; Rey, F.A.; et al. Dengue Virus Sero-Cross-Reactivity Drives Antibody-Dependent Enhancement of Infection with Zika Virus. Nat. Immunol. 2016, 17, 1102–1108. [Google Scholar] [CrossRef]
- Gillespie, L.K.; Hoenen, A.; Morgan, G.; Mackenzie, J.M. The Endoplasmic Reticulum Provides the Membrane Platform for Biogenesis of the Flavivirus Replication Complex. J. Virol. 2010, 84, 10438–10447. [Google Scholar] [CrossRef] [Green Version]
- Welsch, S.; Miller, S.; Romero-Brey, I.; Merz, A.; Bleck, C.K.E.; Walther, P.; Fuller, S.D.; Antony, C.; Krijnse-Locker, J.; Bartenschlager, R. Composition and Three-Dimensional Architecture of the Dengue Virus Replication and Assembly Sites. Cell Host Microbe 2009, 5, 365–375. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.; Kastner, S.; Krijnse-Locker, J.; Bühler, S.; Bartenschlager, R. The Non-Structural Protein 4A of Dengue Virus Is an Integral Membrane Protein Inducing Membrane Alterations in a 2K-Regulated Manner. J. Biol. Chem. 2007, 282, 8873–8882. [Google Scholar] [CrossRef] [Green Version]
- Kaufusi, P.H.; Kelley, J.F.; Yanagihara, R.; Nerurkar, V.R. Induction of Endoplasmic Reticulum-Derived Replication-Competent Membrane Structures by West Nile Virus Non-Structural Protein 4B. PLoS ONE 2014, 9, e84040. [Google Scholar] [CrossRef] [Green Version]
- Roosendaal, J.; Westaway, E.G.; Khromykh, A.; Mackenzie, J.M. Regulated Cleavages at the West Nile Virus NS4A-2K-NS4B Junctions Play a Major Role in Rearranging Cytoplasmic Membranes and Golgi Trafficking of the NS4A Protein. J. Virol. 2006, 80, 4623–4632. [Google Scholar] [CrossRef] [Green Version]
- Akey, D.L.; Brown, W.C.; Dutta, S.; Konwerski, J.; Jose, J.; Jurkiw, T.J.; DelProposto, J.; Ogata, C.M.; Skiniotis, G.; Kuhn, R.J.; et al. Flavivirus NS1 Structures Reveal Surfaces for Associations with Membranes and the Immune System. Science 2014, 343, 881–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, Y.; Okabayashi, T.; Yamashita, T.; Zhao, Z.; Wakita, T.; Yasui, K.; Hasebe, F.; Tadano, M.; Konishi, E.; Moriishi, K.; et al. Nuclear Localization of Japanese Encephalitis Virus Core Protein Enhances Viral Replication. J. Virol. 2005, 79, 3448–3458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogatyreva, N.S.; Finkelstein, A.V.; Galzitskaya, O.V. Trend of Amino Acid Composition of Proteins of Different Taxa. J. Bioinform. Comput. Biol. 2006, 4, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.-P.; Zhang, X.; Han, P.; Arora, N.; Anders, R.F.; Norton, R.S. Abundance of Intrinsically Unstructured Proteins in P. Falciparum and Other Apicomplexan Parasite Proteomes. Mol. Biochem. Parasitol. 2006, 150, 256–267. [Google Scholar] [CrossRef] [Green Version]
- Xue, B.; Dunbrack, R.L.; Williams, R.W.; Dunker, A.K.; Uversky, V.N. PONDR-FIT: A Meta-Predictor of Intrinsically Disordered Amino Acids. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2010, 1804, 996–1010. [Google Scholar] [CrossRef] [Green Version]
- Dunker, A.K.; Lawson, J.D.; Brown, C.J.; Williams, R.M.; Romero, P.; Oh, J.S.; Oldfield, C.J.; Campen, A.M.; Ratliff, C.M.; Hipps, K.W.; et al. Intrinsically Disordered Protein. J. Mol. Graph. Model. 2001, 19, 26–59. [Google Scholar] [CrossRef] [Green Version]
- Ward, J.J.; Sodhi, J.S.; McGuffin, L.J.; Buxton, B.F.; Jones, D.T. Prediction and Functional Analysis of Native Disorder in Proteins from the Three Kingdoms of Life. J. Mol. Biol. 2004, 337, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Schad, E.; Tompa, P.; Hegyi, H. The Relationship between Proteome Size, Structural Disorder and Organism Complexity. Genome Biol. 2011, 12, R120. [Google Scholar] [CrossRef] [Green Version]
- Romero, P.; Obradovic, Z.; Li, X.; Garner, E.C.; Brown, C.J.; Dunker, A.K. Sequence Complexity of Disordered Protein. Proteins Struct. Funct. Genet. 2001, 42, 38–48. [Google Scholar] [CrossRef]
- Pushker, R.; Mooney, C.; Davey, N.E.; Jacqué, J.-M.; Shields, D.C. Marked Variability in the Extent of Protein Disorder within and between Viral Families. PLoS ONE 2013, 8, e60724. [Google Scholar] [CrossRef] [Green Version]
- Drake, J.W.; Charlesworth, B.; Charlesworth, D.; Crow, J.F. Rates of Spontaneous Mutation. Genetics 1998, 148, 1667–1686. [Google Scholar] [CrossRef] [PubMed]
- Gromowski, G.D.; Firestone, C.-Y.; Whitehead, S.S. Genetic Determinants of Japanese Encephalitis Virus Vaccine Strain SA14-14-2 That Govern Attenuation of Virulence in Mice. J. Virol. 2015, 89, 6328–6337. [Google Scholar] [CrossRef] [Green Version]
- dos Santos, C.N.; Post, P.R.; Carvalho, R.; Ferreira, I.I.; Rice, C.M.; Galler, R. Complete Nucleotide Sequence of Yellow Fever Virus Vaccine Strains 17DD and 17D-213. Virus Res. 1995, 35, 35–41. [Google Scholar] [CrossRef]
- Treiber, D.K.; Williamson, J.R. Beyond Kinetic Traps in RNA Folding. Curr. Opin. Struct. Biol. 2001, 11, 309–314. [Google Scholar] [CrossRef]
- Bertrand, E.L.; Rossi, J.J. Facilitation of Hammerhead Ribozyme Catalysis by the Nucleocapsid Protein of HIV-1 and the Heterogeneous Nuclear Ribonucleoprotein A1. EMBO J. 1994, 13, 2904–2912. [Google Scholar] [CrossRef]
- Darlix, J.-L.; Lapadat-Tapolsky, M.; de Rocquigny, H.; Roques, B.P. First Glimpses at Structure-Function Relationships of the Nucleocapsid Protein of Retroviruses. J. Mol. Biol. 1995, 254, 523–537. [Google Scholar] [CrossRef]
- Rein, A.; Henderson, L.E.; Levin, J.G. Nucleic-Acid-Chaperone Activity of Retroviral Nucleocapsid Proteins: Significance for Viral Replication. Trends Biochem. Sci. 1998, 23, 297–301. [Google Scholar] [CrossRef]
- Tompa, P.; Csermely, P. The Role of Structural Disorder in the Function of RNA and Protein Chaperones. FASEB J. 2004, 18, 1169–1175. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Bahadur, R.P. A Structural Perspective of RNA Recognition by Intrinsically Disordered Proteins. Cell. Mol. Life Sci. 2016, 73, 4075–4084. [Google Scholar] [CrossRef]
- Poole, A.M.; Jeffares, D.C.; Penny, D. The Path from the RNA World. J. Mol. Evol. 1998, 46, 1–17. [Google Scholar] [CrossRef]
- Frankel, A.D.; Young, J.A.T. HIV-1: Fifteen Proteins and an RNA. Annu. Rev. Biochem. 1998, 67, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Summers, M.F.; Henderson, L.E.; Chance, M.R.; South, T.L.; Blake, P.R.; Perez-Alvarado, G.; Bess, J.W.; Sowder, R.C.; Arthur, L.O.; Sagi, I.; et al. Nucleocapsid Zinc Fingers Detected in Retroviruses: EXAFS Studies of Intact Viruses and the Solution-State Structure of the Nucleocapsid Protein from HIV-1. Protein Sci. 1992, 1, 563–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morellet, N.; Jullian, N.; De Rocquigny, H.; Maigret, B.; Darlix, J.L.; Roques, B.P. Determination of the Structure of the Nucleocapsid Protein NCp7 from the Human Immunodeficiency Virus Type 1 by 1H NMR. EMBO J. 1992, 11, 3059–3065. [Google Scholar] [CrossRef] [PubMed]
- Xue, B.; Mizianty, M.J.; Kurgan, L.; Uversky, V.N. Protein Intrinsic Disorder as a Flexible Armor and a Weapon of HIV-1. Cell. Mol. Life Sci. 2012, 69, 1211–1259. [Google Scholar] [CrossRef]
- Carteau, S.; Batson, S.C.; Poljak, L.; Mouscadet, J.-F.O.; Darlix, J.-L. Human Immunodeficiency Virus Type 1 Nucleocapsid Protein Specifically Stimulates Mg2+-Dependent DNA Integration In Vitro. J. Virol. 1997, 71, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Khorchid, A.; Wang, J.; Parniak, M.A.; Darlix, J.L.; Wainberg, M.A.; Kleiman, L. Effect of Mutations in the Nucleocapsid Protein (NCp7) upon Pr160(Gag-Pol) and TRNA(Lys) Incorporation into Human Immunodeficiency Virus Type 1. J. Virol. 1997, 71, 4378–4384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Henderson, L.E.; Bess, J.; Kane, B.; Levin, J.G. Human Immunodeficiency Virus Type 1 Nucleocapsid Protein Promotes Efficient Strand Transfer and Specific Viral DNA Synthesis by Inhibiting TAR-Dependent Self-Priming from Minus-Strand Strong-Stop DNA. J. Virol. 1997, 71, 5178–5188. [Google Scholar] [CrossRef] [Green Version]
- Cameron, C.E.; Ghosh, M.; Le Grice, S.F.J.; Benkovic, S.J. Mutations in HIV Reverse Transcriptase Which Alter RNase H Activity and Decrease Strand Transfer Efficiency Are Suppressed by HIV Nucleocapsid Protein. Proc. Natl. Acad. Sci. USA 1997, 94, 6700–6705. [Google Scholar] [CrossRef] [Green Version]
- Cristofari, G. The Hepatitis C Virus Core Protein Is a Potent Nucleic Acid Chaperone That Directs Dimerization of the Viral (+) Strand RNA in Vitro. Nucleic Acids Res. 2004, 32, 2623–2631. [Google Scholar] [CrossRef] [Green Version]
- Ivanyi-Nagy, R.; Kanevsky, I.; Gabus, C.; Lavergne, J.-P.; Ficheux, D.; Penin, F.; Fosse, P.; Darlix, J.-L. Analysis of Hepatitis C Virus RNA Dimerization and Core-RNA Interactions. Nucleic Acids Res. 2006, 34, 2618–2633. [Google Scholar] [CrossRef]
- Ivanyi-Nagy, R.; Lavergne, J.-P.; Gabus, C.; Ficheux, D.; Darlix, J.-L. RNA Chaperoning and Intrinsic Disorder in the Core Proteins of Flaviviridae. Nucleic Acids Res. 2008, 36, 712–725. [Google Scholar] [CrossRef]
- Pong, W.L.; Huang, Z.S.; Teoh, P.G.; Wang, C.C.; Wu, H.N. RNA Binding Property and RNA Chaperone Activity of Dengue Virus Core Protein and Other Viral RNA-Interacting Proteins. FEBS Lett. 2011, 585, 2575–2581. [Google Scholar] [CrossRef] [Green Version]
- Davey, J.; Colman, A.; Dimmock, N.J. Location of Influenza Virus M, NP and NS1 Proteins in Microinjected Cells. J. Gen. Virol. 1985, 66, 2319–2334. [Google Scholar] [CrossRef]
- Michienzi, A.; Cagnon, L.; Bahner, I.; Rossi, J.J. Ribozyme-Mediated Inhibition of HIV 1 Suggests Nucleolar Trafficking of HIV-1 RNA. Proc. Natl. Acad. Sci. USA 2000, 97, 8955–8960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonntag, F.; Schmidt, K.; Kleinschmidt, J.A. A Viral Assembly Factor Promotes AAV2 Capsid Formation in the Nucleolus. Proc. Natl. Acad. Sci. USA 2010, 107, 10220–10225. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, Y.; Mori, Y.; Abe, T.; Yamashita, T.; Okamoto, T.; Ichimura, T.; Moriishi, K.; Matsuura, Y. Nucleolar Protein B23 Interacts with Japanese Encephalitis Virus Core Protein and Participates in Viral Replication. Microbiol. Immunol. 2006, 50, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.T.; Ma, L.; Burgner, J.W.; Groesch, T.D.; Post, C.B.; Kuhn, R.J. Flavivirus Capsid Is a Dimeric Alpha-Helical Protein. J. Virol. 2003, 77, 7143–7149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, T.Y.; Fibriansah, G.; Lok, S.-M. Capsid Protein Is Central to the Birth of Flavivirus Particles. PLoS Pathog. 2020, 16, e1008542. [Google Scholar] [CrossRef]
- Kiermayr, S.; Kofler, R.M.; Mandl, C.W.; Messner, P.; Heinz, F.X. Isolation of Capsid Protein Dimers from the Tick-Borne Encephalitis Flavivirus and in Vitro Assembly of Capsid-like Particles. J. Virol. 2004, 78, 8078–8084. [Google Scholar] [CrossRef] [Green Version]
- Poonsiri, T.; Wright, G.S.A.; Solomon, T.; Antonyuk, S.V. Crystal Structure of the Japanese Encephalitis Virus Capsid Protein. Viruses 2019, 11, 623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, T.Y.; Fibriansah, G.; Kostyuchenko, V.A.; Ng, T.-S.; Lim, X.-X.; Zhang, S.; Lim, X.-N.; Wang, J.; Shi, J.; Morais, M.C.; et al. Capsid Protein Structure in Zika Virus Reveals the Flavivirus Assembly Process. Nat. Commun. 2020, 11, 895. [Google Scholar] [CrossRef]
- Roby, J.A.; Setoh, Y.X.; Hall, R.A.; Khromykh, A.A. Post-Translational Regulation and Modifications of Flavivirus Structural Proteins. J. Gen. Virol. 2015, 96, 1551–1569. [Google Scholar] [CrossRef]
- Oliveira, E.R.A.; Mohana-Borges, R.; de Alencastro, R.B.; Horta, B.A.C. The Flavivirus Capsid Protein: Structure, Function and Perspectives towards Drug Design. Virus Res. 2017, 227, 115–123. [Google Scholar] [CrossRef]
- Khromykh, A.A.; Westaway, E.G. RNA Binding Properties of Core Protein of the Flavivirus Kunjin. Arch. Virol. 1996, 141, 685–699. [Google Scholar] [CrossRef]
- Samsa, M.M.; Mondotte, J.A.; Caramelo, J.J.; Gamarnik, A.V. Uncoupling Cis-Acting RNA Elements from Coding Sequences Revealed a Requirement of the N-Terminal Region of Dengue Virus Capsid Protein in Virus Particle Formation. J. Virol. 2012, 86, 1046–1058. [Google Scholar] [CrossRef] [Green Version]
- Kofler, R.M.; Heinz, F.X.; Mandl, C.W. Capsid Protein C of Tick-Borne Encephalitis Virus Tolerates Large Internal Deletions and Is a Favorable Target for Attenuation of Virulence. J. Virol. 2002, 76, 3534–3543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlick, P.; Taucher, C.; Schittl, B.; Tran, J.L.; Kofler, R.M.; Schueler, W.; von Gabain, A.; Meinke, A.; Mandl, C.W. Helices A2 and A3 of West Nile Virus Capsid Protein Are Dispensable for Assembly of Infectious Virions. J. Virol. 2009, 83, 5581–5591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slomnicki, L.P.; Chung, D.H.; Parker, A.; Hermann, T.; Boyd, N.L.; Hetman, M. Ribosomal Stress and Tp53-Mediated Neuronal Apoptosis in Response to Capsid Protein of the Zika Virus. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Anderson, R.; Hobman, T.C. The Capsid-Binding Nucleolar Helicase DDX56 Is Important for Infectivity of West Nile Virus. J. Virol. 2011, 85, 5571–5580. [Google Scholar] [CrossRef] [Green Version]
- Ishida, K.; Goto, S.; Ishimura, M.; Amanuma, M.; Hara, Y.; Suzuki, R.; Katoh, K.; Morita, E. Functional Correlation between Subcellular Localizations of Japanese Encephalitis Virus Capsid Protein and Virus Production. J. Virol. 2019, 93, e00612-19. [Google Scholar] [CrossRef]
- Tiwary, A.K.; Cecilia, D. Kinetics of the Association of Dengue Virus Capsid Protein with the Granular Component of Nucleolus. Virology 2017, 502, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Kunkel, M.; Lorinczi, M.; Rijnbrand, R.; Lemon, S.M.; Watowich, S.J. Self-Assembly of Nucleocapsid-Like Particles from Recombinant Hepatitis C Virus Core Protein. J. Virol. 2001, 75, 2119–2129. [Google Scholar] [CrossRef] [Green Version]
- Molliex, A.; Temirov, J.; Lee, J.; Coughlin, M.; Kanagaraj, A.P.; Kim, H.J.; Mittag, T.; Taylor, J.P. Phase Separation by Low Complexity Domains Promotes Stress Granule Assembly and Drives Pathological Fibrillization. Cell 2015, 163, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.; Lee, H.O.; Jawerth, L.; Maharana, S.; Jahnel, M.; Hein, M.Y.; Stoynov, S.; Mahamid, J.; Saha, S.; Franzmann, T.M.; et al. A Liquid-to-Solid Phase Transition of the ALS Protein FUS Accelerated by Disease Mutation. Cell 2015, 162, 1066–1077. [Google Scholar] [CrossRef] [Green Version]
- Kedersha, N.; Panas, M.D.; Achorn, C.A.; Lyons, S.; Tisdale, S.; Hickman, T.; Thomas, M.; Lieberman, J.; McInerney, G.M.; Ivanov, P.; et al. G3BP–Caprin1–USP10 Complexes Mediate Stress Granule Condensation and Associate with 40S Subunits. J. Cell Biol. 2016, 212, e201508028. [Google Scholar] [CrossRef] [Green Version]
- Courtney, S.C.; Scherbik, S.V.; Stockman, B.M.; Brinton, M.A. West Nile Virus Infections Suppress Early Viral RNA Synthesis and Avoid Inducing the Cell Stress Granule Response. J. Virol. 2012, 86, 3647–3657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonenfant, G.; Williams, N.; Netzband, R.; Schwarz, M.C.; Evans, M.J.; Pager, C.T. Zika Virus Subverts Stress Granules To Promote and Restrict Viral Gene Expression. J. Virol. 2019, 93, 22. [Google Scholar] [CrossRef] [Green Version]
- Amorim, R.; Temzi, A.; Griffin, B.D.; Mouland, A.J. Zika Virus Inhibits EIF2α-Dependent Stress Granule Assembly. PLoS Negl. Trop. Dis. 2017, 11, e0005775. [Google Scholar] [CrossRef] [Green Version]
- Hou, S.; Kumar, A.; Xu, Z.; Airo, A.M.; Stryapunina, I.; Wong, C.P.; Branton, W.; Tchesnokov, E.; Götte, M.; Power, C.; et al. Zika Virus Hijacks Stress Granule Proteins and Modulates the Host Stress Response. J. Virol. 2017, 91, e00474-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blázquez, A.-B.; Martín-Acebes, M.A.; Poderoso, T.; Saiz, J.-C. Relevance of Oxidative Stress in Inhibition of EIF2 Alpha Phosphorylation and Stress Granules Formation during Usutu Virus Infection. PLOS Negl. Trop. Dis. 2021, 15, e0009072. [Google Scholar] [CrossRef] [PubMed]
- Basu, M.; Courtney, S.C.; Brinton, M.A. Arsenite-Induced Stress Granule Formation Is Inhibited by Elevated Levels of Reduced Glutathione in West Nile Virus-Infected Cells. PLoS Pathog. 2017, 13, e1006240. [Google Scholar] [CrossRef]
- Bidet, K.; Dadlani, D.; Garcia-Blanco, M.A. G3BP1, G3BP2 and CAPRIN1 Are Required for Translation of Interferon Stimulated MRNAs and Are Targeted by a Dengue Virus Non-Coding RNA. PLOS Pathog. 2014, 10, e1004242. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.S.-Y.; Sze, L.; Lam, K.-P. The Stress Granule Protein G3BP1 Binds Viral DsRNA and RIG-I to Enhance Interferon-β Response. J. Biol. Chem. 2019, 294, 6430–6438. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Ru, Y.; Ren, J.; Bai, J.; Wei, J.; Fu, S.; Liu, X.; Li, D.; Zheng, H. G3BP1 Inhibits RNA Virus Replication by Positively Regulating RIG-I-Mediated Cellular Antiviral Response. Cell Death Dis. 2019, 10, 946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Alam, U.; Willis, C.; Kennedy, D. Role of Chikungunya NsP3 in Regulating G3BP1 Activity, Stress Granule Formation and Drug Efficacy. Arch. Med. Res. 2021, 52, 48–57. [Google Scholar] [CrossRef]
- Fros, J.J.; Domeradzka, N.E.; Baggen, J.; Geertsema, C.; Flipse, J.; Vlak, J.M.; Pijlman, G.P. Chikungunya Virus NsP3 Blocks Stress Granule Assembly by Recruitment of G3BP into Cytoplasmic Foci. J. Virol. 2012, 86, 10873–10879. [Google Scholar] [CrossRef] [Green Version]
- Götte, B.; Utt, A.; Fragkoudis, R.; Merits, A.; McInerney, G.M. Sensitivity of Alphaviruses to G3BP Deletion Correlates with Efficiency of Replicase Polyprotein Processing. J. Virol. 2020, 94, e01681-19. [Google Scholar] [CrossRef] [PubMed]
- Alam, U.; Kennedy, D. G3BP1 and G3BP2 Regulate Translation of Interferon-Stimulated Genes: IFITM1, IFITM2 and IFITM3 in the Cancer Cell Line MCF7. Mol. Cell. Biochem. 2019, 459, 189–204. [Google Scholar] [CrossRef]
- Reineke, L.C.; Tsai, W.-C.; Jain, A.; Kaelber, J.T.; Jung, S.Y.; Lloyd, R.E. Casein Kinase 2 Is Linked to Stress Granule Dynamics through Phosphorylation of the Stress Granule Nucleating Protein G3BP1. Mol. Cell. Biol. 2017, 37. [Google Scholar] [CrossRef] [Green Version]
- Masaki, T.; Matsunaga, S.; Takahashi, H.; Nakashima, K.; Kimura, Y.; Ito, M.; Matsuda, M.; Murayama, A.; Kato, T.; Hirano, H.; et al. Involvement of Hepatitis C Virus NS5A Hyperphosphorylation Mediated by Casein Kinase I- in Infectious Virus Production. J. Virol. 2014, 88, 7541–7555. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Hwang, J.; Sharma, S.D.; Hargittai, M.R.S.; Chen, Y.; Arnold, J.J.; Raney, K.D.; Cameron, C.E. Hepatitis C Virus Nonstructural Protein 5A (NS5A) Is an RNA-Binding Protein. J. Biol. Chem. 2005, 280, 36417–36428. [Google Scholar] [CrossRef] [Green Version]
- Morozova, O.V.; Tsekhanovskaya, N.A.; Maksimova, T.G.; Bachvalova, V.N.; Matveeva, V.A.; Kit, Y.Y. Phosphorylation of Tick-Borne Encephalitis Virus NS5 Protein. Virus Res. 1997, 49, 9–15. [Google Scholar] [CrossRef]
- Mackenzie, J.M.; Kenney, M.T.; Westaway, E.G. West Nile Virus Strain Kunjin NS5 Polymerase Is a Phosphoprotein Localized at the Cytoplasmic Site of Viral RNA Synthesis. J. Gen. Virol. 2007, 88, 1163–1168. [Google Scholar] [CrossRef] [PubMed]
- Reed, K.E.; Gorbalenya, A.E.; Rice, C.M. The NS5A/NS5 Proteins of Viruses from Three Genera of the Family Flaviviridae Are Phosphorylated by Associated Serine/Threonine Kinases. J. Virol. 1998, 72, 6199–6206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapoor, M.; Zhang, L.; Ramachandra, M.; Kusukawa, J.; Ebner, K.E.; Padmanabhan, R. Association between NS3 and NS5 Proteins of Dengue Virus Type 2 in the Putative RNA Replicase Is Linked to Differential Phosphorylation of NS5. J. Biol. Chem. 1995, 270, 19100–19106. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, D.; Ansari, I.H.; Striker, R. The Flaviviral Methyltransferase Is a Substrate of Casein Kinase 1. Virus Res. 2009, 141, 101–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keating, J.A.; Bhattacharya, D.; Lim, P.-Y.; Falk, S.; Weisblum, B.; Bernard, K.A.; Sharma, M.; Kuhn, R.J.; Striker, R. West Nile Virus Methyltransferase Domain Interacts with Protein Kinase G. Virol. J. 2013, 10, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iakoucheva, L.M. The Importance of Intrinsic Disorder for Protein Phosphorylation. Nucleic Acids Res. 2004, 32, 1037–1049. [Google Scholar] [CrossRef] [Green Version]
- Darling, A.L.; Uversky, V.N. Intrinsic Disorder and Posttranslational Modifications: The Darker Side of the Biological Dark Matter. Front. Genet. 2018, 9, 158. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, H.; Kosako, H.; Yoshimura, S.H. Quantitative Proteomics Indicate a Strong Correlation of Mitotic Phospho-/Dephosphorylation with Non-Structured Regions of Substrates. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2020, 1868, 140295. [Google Scholar] [CrossRef]
- Wippich, F.; Bodenmiller, B.; Trajkovska, M.G.; Wanka, S.; Aebersold, R.; Pelkmans, L. Dual Specificity Kinase DYRK3 Couples Stress Granule Condensation/Dissolution to MTORC1 Signaling. Cell 2013, 152, 791–805. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.T.; Smith, J.; Chen, B.-C.; Schmidt, H.; Rasoloson, D.; Paix, A.; Lambrus, B.G.; Calidas, D.; Betzig, E.; Seydoux, G. Regulation of RNA Granule Dynamics by Phosphorylation of Serine-Rich, Intrinsically Disordered Proteins in C. Elegans. eLife 2014, 3, e04591. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Conicella, A.E.; Schmidt, H.B.; Martin, E.W.; Rhoads, S.N.; Reeb, A.N.; Nourse, A.; Ramirez Montero, D.; Ryan, V.H.; Rohatgi, R.; et al. A Single N-terminal Phosphomimic Disrupts TDP-43 Polymerization, Phase Separation, and RNA Splicing. EMBO J. 2018, 37, e97452. [Google Scholar] [CrossRef]
- Greig, J.A.; Nguyen, T.A.; Lee, M.; Holehouse, A.S.; Posey, A.E.; Pappu, R.V.; Jedd, G. Arginine-Enriched Mixed-Charge Domains Provide Cohesion for Nuclear Speckle Condensation. Mol. Cell 2020, 77, 1237–1250. [Google Scholar] [CrossRef]
- Das, S.; Eisen, A.; Lin, Y.-H.; Chan, H.S. A Lattice Model of Charge-Pattern-Dependent Polyampholyte Phase Separation. J. Phys. Chem. B 2018, 122, 5418–5431. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-H.; Chan, H.S. Phase Separation and Single-Chain Compactness of Charged Disordered Proteins Are Strongly Correlated. Biophys. J. 2017, 112, 2043–2046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.-H.; Song, J.; Forman-Kay, J.D.; Chan, H.S. Random-Phase-Approximation Theory for Sequence-Dependent, Biologically Functional Liquid-Liquid Phase Separation of Intrinsically Disordered Proteins. J. Mol. Liq. 2017, 228, 176–193. [Google Scholar] [CrossRef] [Green Version]
- Castelnovo, M.; Joanny, J.F. Phase Diagram of Diblock Polyampholyte Solutions. Macromolecules 2002, 35, 4531–4538. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Cui, Y.; Han, X.; Hu, W.; Sun, M.; Zhang, Y.; Wang, P.-H.; Song, G.; Chen, W.; Lou, J. Liquid–Liquid Phase Separation by SARS-CoV-2 Nucleocapsid Protein and RNA. Cell Res. 2020, 30, 1143–1145. [Google Scholar] [CrossRef]
- Carlson, C.R.; Asfaha, J.B.; Ghent, C.M.; Howard, C.J.; Hartooni, N.; Safari, M.; Frankel, A.D.; Morgan, D.O. Phosphoregulation of Phase Separation by the SARS-CoV-2 N Protein Suggests a Biophysical Basis for Its Dual Functions. Mol. Cell 2020, 80, 1092–1103. [Google Scholar] [CrossRef] [PubMed]
- Savastano, A.; Ibáñez de Opakua, A.; Rankovic, M.; Zweckstetter, M. Nucleocapsid Protein of SARS-CoV-2 Phase Separates into RNA-Rich Polymerase-Containing Condensates. Nat. Commun. 2020, 11, 6041. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Yu, Y.; Sun, L.-M.; Xing, J.-Q.; Li, T.; Zhu, Y.; Wang, M.; Yu, Y.; Xue, W.; Xia, T.; et al. GCG Inhibits SARS-CoV-2 Replication by Disrupting the Liquid Phase Condensation of Its Nucleocapsid Protein. Nat. Commun. 2021, 12, 2114. [Google Scholar] [CrossRef]
- Guseva, S.; Milles, S.; Jensen, M.R.; Salvi, N.; Kleman, J.-P.; Maurin, D.; Ruigrok, R.W.H.; Blackledge, M. Measles Virus Nucleo- and Phosphoproteins Form Liquid-like Phase-Separated Compartments that Promote Nucleocapsid Assembly. Sci. Adv. 2020, 6, eaaz7095. [Google Scholar] [CrossRef] [Green Version]
- Nevers, Q.; Albertini, A.A.; Lagaudrière-Gesbert, C.; Gaudin, Y. Negri Bodies and Other Virus Membrane-Less Replication Compartments. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118831. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).