
A Single Point Mutation, Asn16→Lys, Dictates the Temperature-Sensitivity of the Reovirus tsG453 Mutant

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
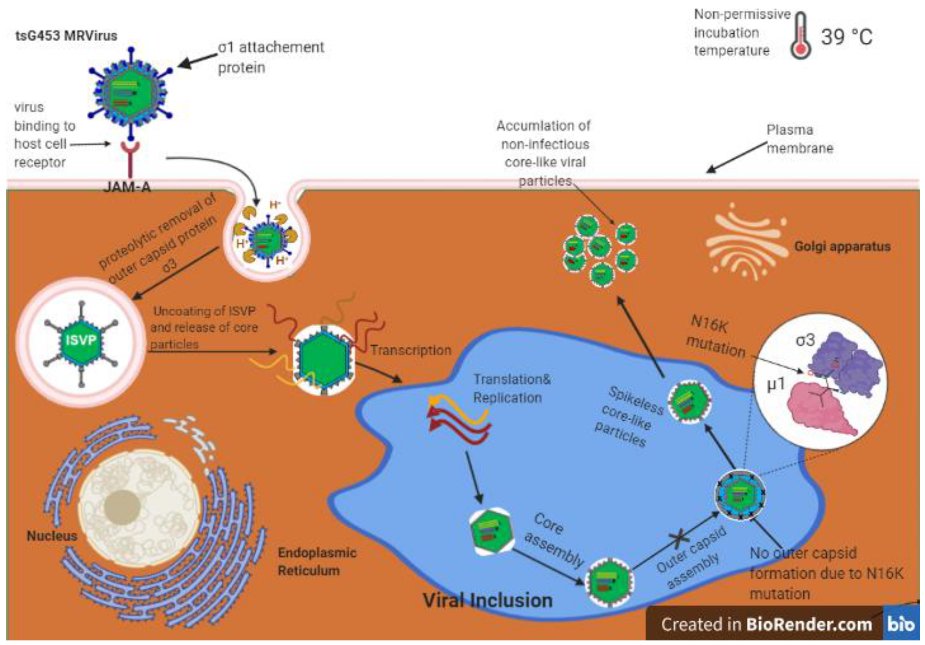{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Plasmids
2.3. Primer Design for ts Mutants
- T3D S2 A188V F-CAATGTGTATGCAATCTCTACAAACGTGTGCCCGAAATA
- T3D S2 A188V R-TATTTCGGGCACACGTTTGTAGAGATTGCATACACATTG
- T3D S2 A323V F-CATGCAATTGGTTACCAACTCTACCAGTCCAGCCA
- T3D S2 A323V R-TGGCTGGACTGGTAGAGTTGGTAACCAATTGCATG
- T3D S2 N383D F-GGATGAGCCTGACTATATTGATCGTCTTCTCTCGCC
- T3D S2 N383D R-GGCGAGAGAAGACGATCAATATAGTCAGGCTCATCC
- T3D S2 N383S F-GGATGAGCCTGACTCTATTGATCGTCTTCTCTCGCC
- T3D S2 N383S R-GGCGAGAGAAGACGATCAATATAGTCAGGCTCATCC
- T3D S4 N16K F-CCTTCAAAAGCGTTCTTAATCAAGTCCACGACCTGAT
- T3D S4 N16K R-ATCAGGTCGTGGACTTGATTAAGAACGCTTTTGAAGG
- T3D S4 M141I F-CAACTTGAGTGTATTGATCTAAATATTGAATTTGGGTCAACCTGAAG
- T3D S4 M141I R-CTTCAGGTTGACCCAAATTCAATATTTAGATCAATACACTCAAGTTG
- T3D S4 E229D F-CCCTTCGATGGATCATGATCCAGCTCAGAGTAATC
- T3D S4 E229D R-GATTACTCTGAGCTGGATCATGATCCATCGAAGGG
2.4. Reovirus Plasmid Mutagenesis
2.5. Reovirus Reverse Genetics
2.6. Sanger Sequencing for Rescued Virus
- T3D S2 F-GCTATTCGCTGGTCAGTTAT
- T3D S2 R-ATGAATGTGTGGTCAGTCGT
- T3D S4 F-CGTTGTCGCAATGGAGGTGTGCTTGC
- T3D S4 R-AGCCTGTCCCACGTCACACC
2.7. Reovirus Efficiency of Plating (EOP) Assays
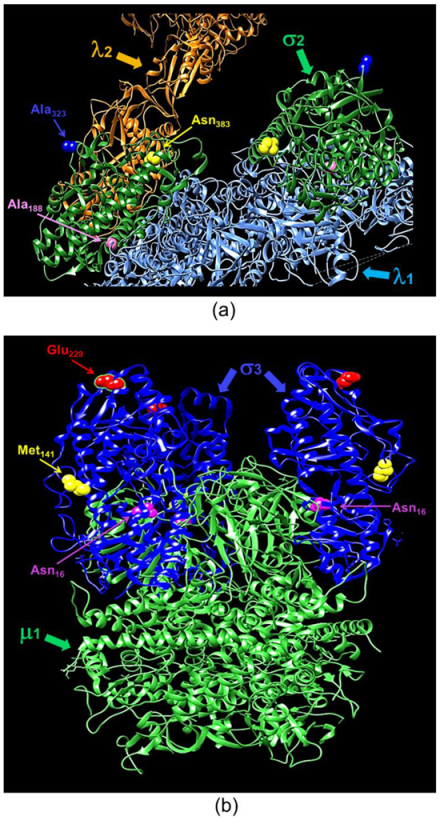
2.8. 3-Dimensional Protein Analyses
3. Results
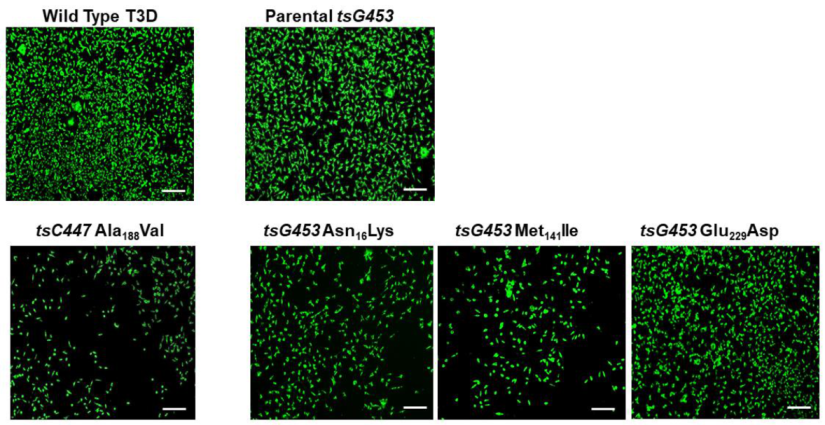
3.1. Reverse Genetics Rescue of Infectious Reovirus Clones Containing Individual and Sets of tsC447 and tsG453 Temperature-Sensitive Alterations
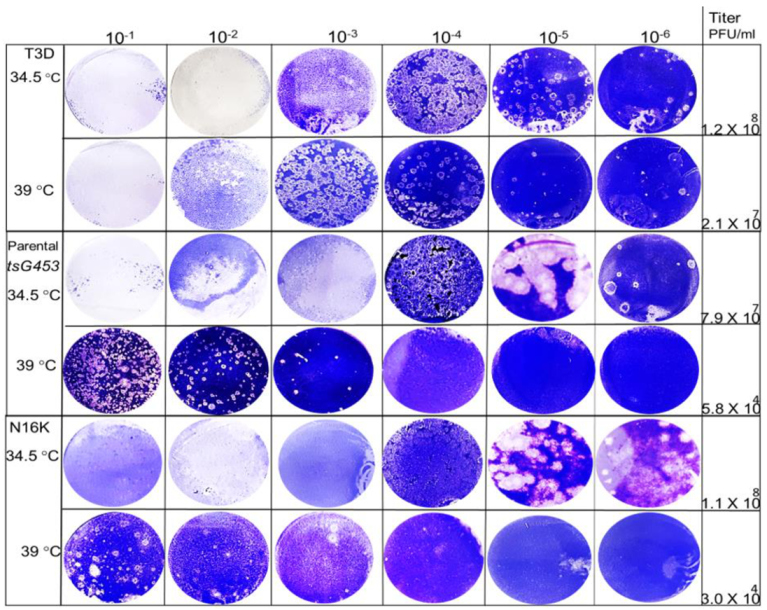
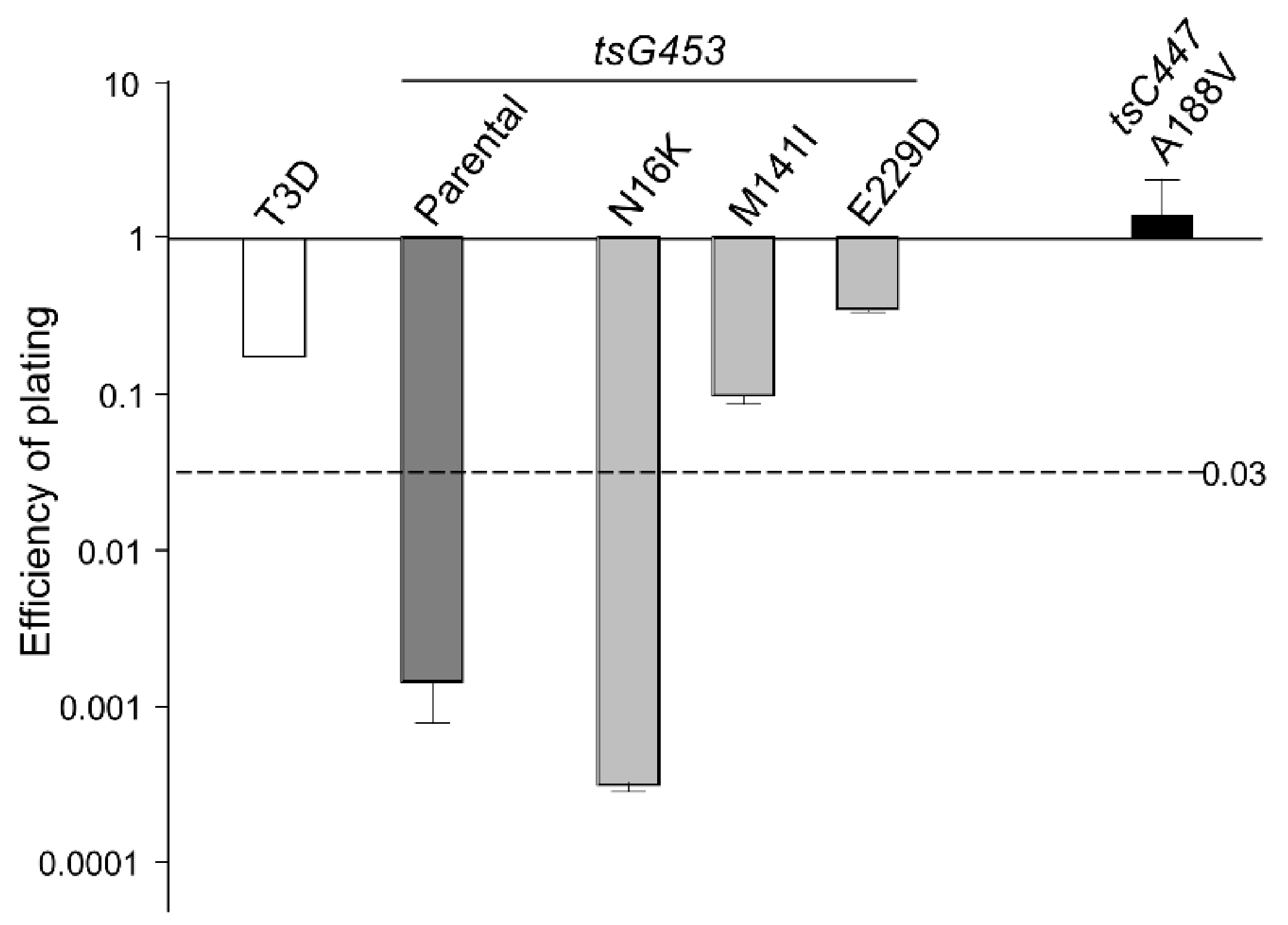
3.2. Efficiency of Plating Values of Various Rescued Clones
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dermody, T.S.; Parker, J.S.L.; Sherry, B. Orthoreoviruses. In Fields Virology, 6th ed.; Knipe, D., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; Volume 2. [Google Scholar]
- Abad, A.T.; Danthi, P. Recognition of reovirus rnas by the innate immune system. Viruses (Basel) 2020, 12, 667. [Google Scholar] [CrossRef] [PubMed]
- Boehme, K.W.; Lai, C.M.; Dermody, T.S. Mechanisms of reovirus bloodstream dissemination. Adv. Virus Res. 2013, 87, 1–35. [Google Scholar] [PubMed]
- Estes, M.K.; Kapikian, A.Z. Rotaviruses. In Fields Virology; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; Volume 5, pp. 1917–1974. [Google Scholar]
- Tate, J.E.; Burton, A.H.; Boschi-Pinto, C.; Steele, A.D.; Duque, J.; Parashar, U.D.; WHO-coordinated Global Rotavirus Surveillance Network. 2008 estimate of worldwide rotavirus-associated mortality in children younger than 5 years before the introduction of universal rotavirus vaccination programmes: A systematic review and meta-analysis. Lancet Infect. Dis. 2012, 12, 136–141. [Google Scholar] [CrossRef]
- Karampatsas, K.; Osborne, L.; Seat, M.L.; Tong, C.Y.W.; Prendergast, A.J. Clinical characteristics and complications of rotavirus gastroenteritis in children in east london: A retrospective case-control study. PLoS ONE 2018, 13, e0194009. [Google Scholar] [CrossRef]
- Roy, P. Orbiviruses. In Fields Virology; Knipe, D.M., Howley, P.M., Eds.; Lippencott Williams & Wilkins: Philadelphia, PA, USA, 2007; Volume 5, pp. 1975–1997. [Google Scholar]
- Coetzee, P.; Stokstad, M.; Venter, E.H.; Myrmel, M.; Van Vuuren, M. Bluetongue: A historical and epidemiological perspective with the emphasis on south africa. Virol. J. 2012, 9. [Google Scholar] [CrossRef]
- Gong, J.; Sachdev, E.; Mita, A.C.; Mita, M.M. Clinical development of reovirus for cancer therapy: An oncolytic virus with immune-mediated antitumor activity. World J. Methodol. 2016, 6, 25–42. [Google Scholar] [CrossRef]
- Strong, J.E.; Coffey, M.C.; Tang, D.; Sabinin, P.; Lee, P.W. The molecular basis of viral oncolysis: Usurpation of the ras signaling pathway by reovirus. EMBO J. 1998, 17, 3351–3362. [Google Scholar] [CrossRef]
- Shmulevitz, M.; Marcato, P.; Lee, P.W. Unshackling the links between reovirus oncolysis, ras signaling, translational control and cancer. Oncogene 2005, 24, 7720–7728. [Google Scholar] [CrossRef]
- Galanis, E.; Markovic, S.N.; Suman, V.J.; Nuovo, G.J.; Vile, R.G.; Kottke, T.J.; Nevala, W.K.; Thompson, M.A.; Lewis, J.E.; Rumilla, K.M.; et al. Phase ii trial of intravenous administration of reolysin (r) (reovirus serotype-3-dearing strain) in patients with metastatic melanoma. Mol. Ther. 2012, 20, 1998–2003. [Google Scholar] [CrossRef]
- Chen, H.F.; Ramachandra, M.; Padmanabhan, R. Biochemical characterization of a temperature-sensitive adenovirus DNA polymerase. Virology 1994, 205, 364–370. [Google Scholar] [CrossRef]
- Millns, A.K.; Carpenter, M.S.; DeLange, A.M. The vaccinia virus-encoded uracil DNA glycosylase has an essential role in viral DNA replication. Virology 1994, 198, 504–513. [Google Scholar] [CrossRef]
- Schwartzberg, P.L.; Roth, M.J.; Tanese, N.; Goff, S.P. Analysis of a temperature-sensitive mutation affecting the integration protein of moloney murine leukemia virus. Virology 1993, 192, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.D.; Dzianott, A.; Ahlquist, P.; Bujarski, J.J. Mutations in the helicase-like domain of protein 1a alter the sites of rna-rna recombination in brome mosaic virus. J. Virol. 1995, 69, 2547–2556. [Google Scholar] [CrossRef]
- Shikova, E.; Lin, Y.C.; Saha, K.; Brooks, B.R.; Wong, P.K.Y. Correlation of specific virus-astrocyte interactions and cytopathic effects induced by ts1, a neurovirulent mutant of moloney murine leukemia virus. J. Virol. 1993, 67, 1137–1147. [Google Scholar] [CrossRef] [PubMed]
- Wiskerchen, M.; Muesing, M.A. Human-immunodeficiency-virus type-1 integrase—effects of mutations on viral ability to integrate, direct viral gene-expression from unintegrated viral-DNA templates, and sustain viral propagation in primary-cells. J. Virol. 1995, 69, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Coombs, K.M. Temperature-sensitive mutants of reovirus. Curr. Top. Microbiol. Immunol. 1998, 233, 69–107. [Google Scholar] [PubMed]
- Black, L.W.; Showe, M.K.; Steven, A.C. Morphogenesis of the t4 head. In Molecular Biology of Bacteriophage T4; Karam, J.D., Drake, J.W., Kreuzer, K.N., Mosig, G., Hall, D., Eiserling, F.A., Black, L.W., Kutter, E., Spicer, E., Carlson, K., et al., Eds.; American Society for Microbiology: Washington, DC, USA, 1994; pp. 218–258. [Google Scholar]
- Tolley, K.P.; Marriott, A.C.; Simpson, A.; Plows, D.J.; Matthews, D.A.; Longhurst, S.J.; Evans, J.E.; Johnson, J.L.; Cane, P.A.; Randolph, V.B.; et al. Identification of mutations contributing to the reduced virulence of a modified strain of respiratory syncytial virus. Vaccine 1996, 14, 1637–1646. [Google Scholar] [CrossRef]
- Murphy, B.R.; Richman, D.D.; Spring, S.B.; Chanock, R.M. Use of temperature-sensitive mutants of influenza-a virus as live virus-vaccine strains—evaluation in laboratory-animals, adults and children. Postgrad. Med. J. 1976, 52, 381–388. [Google Scholar] [CrossRef][Green Version]
- Ikegami, N.; Gomatos, P.J. Temperature-sensitive conditional-lethal mutants of reovirus 3: I. Isolation and characterization. Virology 1968, 36, 447–458. [Google Scholar] [CrossRef]
- Fields, B.N.; Joklik, W.K. Isolation and preliminary genetic and biochemical characterization of temperature-sensitive mutants of reovirus. Virology 1969, 37, 335–342. [Google Scholar] [CrossRef]
- Fields, B.N. Temperature-sensitive mutants of reovirus type 3 features of genetic recombination. Virology 1971, 46, 142–148. [Google Scholar] [CrossRef]
- Fields, B.N.; Laskov, R.; Scharff, M.D. Temperature-sensitive mutants of reovirus type 3: Studies on the synthesis of viral peptides. Virology 1972, 50, 209–215. [Google Scholar] [CrossRef]
- Ramig, R.F.; Fields, B.N. Method for rapidly screening revertants of reovirus temperature-sensitive mutants for extragenic suppression. Virology 1977, 81, 170–173. [Google Scholar]
- Ramig, R.F.; Mustoe, T.A.; Sharpe, A.H.; Fields, B.N. A genetic map of reovirus. Ii. Assignment of the double-stranded rna-negative mutant groups c, d, and e to genome segments. Virology 1978, 85, 531–534. [Google Scholar] [CrossRef]
- Mustoe, T.A.; Ramig, R.F.; Sharpe, A.H.; Fields, B.N. Genetic map of reovirus.3. Assignment of double-stranded rna-positive mutant group-a, group-b, and group-g to genome segments. Virology 1978, 85, 545–556. [Google Scholar] [CrossRef]
- Chakraborty, P.R.; Ahmed, R.; Fields, B.N. Genetics of reovirus: The relationship of interference to complementation and reassortment of temperature-sensitive mutants at nonpermissive temperature. Virology 1979, 94, 119–127. [Google Scholar] [CrossRef]
- Hazelton, P.R.; Coombs, K.M. The reovirus mutant tsa279 has temperature-sensitive lesions in the M2 and L2 genes: The M2 gene is associated with decreased viral protein production and blockade in transmembrane transport. Virology 1995, 207, 46–58. [Google Scholar] [CrossRef]
- Coombs, K.M. Identification and characterization of a double-stranded rna- reovirus temperature-sensitive mutant defective in minor core protein mu2. J. Virol. 1996, 70, 4237–4245. [Google Scholar] [CrossRef] [PubMed]
- Hazelton, P.R.; Coombs, K.M. The reovirus mutant tsa279 L2 gene is associated with generation of a spikeless core particle: Implications for capsid assembly. J. Virol. 1999, 73, 2298–2308. [Google Scholar] [CrossRef]
- Wiener, J.R.; McLaughlin, T.; Joklik, W.K. The sequences of the S2 genome segments of reovirus serotype 3 and of the dsrna-negative mutant ts447. Virology 1989, 170, 340–341. [Google Scholar] [CrossRef]
- Dermody, T.S.; Schiff, L.A.; Nibert, M.L.; Coombs, K.M.; Fields, B.N. The S2 gene nucleotide sequences of prototype strains of the three reovirus serotypes: Characterization of reovirus core protein sigma 2. J. Virol. 1991, 65, 5721–5731. [Google Scholar] [CrossRef] [PubMed]
- Danis, C.; Garzon, S.; Lemay, G. Further characterization of the ts453 mutant of mammalian orthoreovirus serotype 3 and nucleotide sequence of the mutated s4 gene. Virology 1992, 190, 494–498. [Google Scholar] [CrossRef]
- Shing, M.; Coombs, K.M. Assembly of the reovirus outer capsid requires mu 1/sigma 3 interactions which are prevented by misfolded sigma 3 protein in temperature-sensitive mutant tsg453. Virus Res. 1996, 46, 19–29. [Google Scholar] [CrossRef]
- Coombs, K.M.; Mak, S.C.; Petrycky-Cox, L.D. Studies of the major reovirus core protein sigma 2: Reversion of the assembly-defective mutant tsc447 is an intragenic process and involves back mutation of asp-383 to asn. J. Virol. 1994, 68, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Antar, A.A.; Boehme, K.W.; Danthi, P.; Eby, E.A.; Guglielmi, K.M.; Holm, G.H.; Johnson, E.M.; Maginnis, M.S.; Naik, S.; et al. A plasmid-based reverse genetics system for animal double-stranded rna viruses. Cell Host Microbe 2007, 1, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Ooms, L.S.; Ikizler, M.; Chappell, J.D.; Dermody, T.S. An improved reverse genetics system for mammalian orthoreoviruses. Virology 2010, 398, 194–200. [Google Scholar] [CrossRef]
- Boehme, K.W.; Ikizler, M.; Kobayashi, T.; Dermody, T.S. Reverse genetics for mammalian reovirus. Methods Companion Methods Enzymol. 2011, 55, 109–113. [Google Scholar] [CrossRef]
- Dryden, K.A.; Wang, G.; Yeager, M.; Nibert, M.L.; Coombs, K.M.; Furlong, D.B.; Fields, B.N.; Baker, T.S. Early steps in reovirus infection are associated with dramatic changes in supramolecular structure and protein conformation: Analysis of virions and subviral particles by cryoelectron microscopy and image reconstruction. J. Cell Biol. 1993, 122, 1023–1041. [Google Scholar] [CrossRef]
- Reinisch, K.M.; Nibert, M.L.; Harrison, S.C. Structure of the reovirus core at 3.6 a resolution. Nature 2000, 404, 960–967. [Google Scholar] [CrossRef]
- Matsuhisa, T.; Joklik, W.K. Temperature-sensitive mutants of reovirus. V. Studies on the nature of the temperature-sensitive lesion of the group c mutant ts447. Virology 1974, 60, 380–389. [Google Scholar] [CrossRef]
- Gaspar, P.; Moura, G.; Santos, M.A.S.; Oliveira, J.L. Mrna secondary structure optimization using a correlated stem-loop prediction. Nucleic Acids Res. 2013, 41, e73. [Google Scholar] [CrossRef]
- Mao, Y.H.; Liu, H.L.; Liu, Y.L.; Tao, S.H. Deciphering the rules by which dynamics of mrna secondary structure affect translation efficiency in saccharomyces cerevisiae. Nucleic Acids Res. 2014, 42, 4813–4822. [Google Scholar] [CrossRef]
- Gruber, A.R.; Lorenz, R.; Bernhart, S.H.; Neuboock, R.; Hofacker, I.L. The vienna rna websuite. Nucleic Acids Res. 2008, 36, W70–W74. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, R.; Bernhart, S.H.; Siederdissen, C.H.Z.; Tafer, H.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. Viennarna package 2.0. Algorithms Mol. Biol. 2011, 6, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Coombs, K.M. Stoichiometry of reovirus structural proteins in virus, isvp, and core particles. Virology 1998, 243, 218–228. [Google Scholar] [CrossRef]
- Ebert, D.H.; Deussing, J.; Peters, C.; Dermody, T.S. Cathepsin l and cathepsin b mediate reovirus disassembly in murine fibroblast cells. J. Biol. Chem. 2002, 277, 24609–24617. [Google Scholar] [CrossRef] [PubMed]
- Jane-Valbuena, J.; Breun, L.A.; Schiff, L.A.; Nibert, M.L. Sites and determinants of early cleavages in the proteolytic processing pathway of reovirus surface protein sigma3. J. Virol. 2002, 76, 5184–5197. [Google Scholar] [CrossRef]
- Mendez, I.I.; She, Y.M.; Ens, W.; Coombs, K.M. Digestion pattern of reovirus outer capsid protein sigma3 determined by mass spectrometry. Virology 2003, 311, 289–304. [Google Scholar] [CrossRef]
- Golden, J.W.; Bahe, J.A.; Lucas, W.T.; Nibert, M.L.; Schiff, L.A. Cathepsin s supports acid-independent infection by some reoviruses. J. Biol. Chem. 2004, 279, 8547–8557. [Google Scholar] [CrossRef]
- Bodkin, D.K.; Nibert, M.L.; Fields, B.N. Proteolytic digestion of reovirus in the intestinal lumens of neonatal mice. J. Virol. 1989, 63, 4676–4681. [Google Scholar] [CrossRef]
- Bass, D.M.; Bodkin, D.; Dambrauskas, R.; Trier, J.S.; Fields, B.N.; Wolf, J.L. Intraluminal proteolytic activation plays an important role in replication of type 1 reovirus in the intestines of neonatal mice. J. Virol. 1990, 64, 1830–1833. [Google Scholar] [CrossRef] [PubMed]
- Imani, F.; Jacobs, B.L. Inhibitory activity for the interferon-induced protein kinase is associated with the reovirus serotype 1 sigma 3 protein. Proc. Natl. Acad. Sci. USA 1988, 85, 7887–7891. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.A.; Schmechel, S.C.; Williams, B.R.; Silverman, R.H.; Schiff, L.A. Involvement of the interferon-regulated antiviral proteins pkr and rnase l in reovirus-induced shutoff of cellular translation. J. Virol. 2005, 79, 2240–2250. [Google Scholar] [CrossRef] [PubMed]
- Cross, R.K.; Fields, B.N. Temperature-sensitive mutants of reovirus type 3: Studies on the synthesis of viral rna. Virology 1972, 50, 799–809. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glover, K.K.M.; Sutherland, D.M.; Dermody, T.S.; Coombs, K.M. A Single Point Mutation, Asn16→Lys, Dictates the Temperature-Sensitivity of the Reovirus tsG453 Mutant. Viruses 2021, 13, 289. https://doi.org/10.3390/v13020289
Glover KKM, Sutherland DM, Dermody TS, Coombs KM. A Single Point Mutation, Asn16→Lys, Dictates the Temperature-Sensitivity of the Reovirus tsG453 Mutant. Viruses. 2021; 13(2):289. https://doi.org/10.3390/v13020289
Chicago/Turabian StyleGlover, Kathleen K. M., Danica M. Sutherland, Terence S. Dermody, and Kevin M. Coombs. 2021. "A Single Point Mutation, Asn16→Lys, Dictates the Temperature-Sensitivity of the Reovirus tsG453 Mutant" Viruses 13, no. 2: 289. https://doi.org/10.3390/v13020289
APA StyleGlover, K. K. M., Sutherland, D. M., Dermody, T. S., & Coombs, K. M. (2021). A Single Point Mutation, Asn16→Lys, Dictates the Temperature-Sensitivity of the Reovirus tsG453 Mutant. Viruses, 13(2), 289. https://doi.org/10.3390/v13020289