
Ongoing Assessment of the Molecular Evolution of Peste Des Petits Ruminants Virus Continues to Question Viral Origins

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Whole Genome NGS
2.2. Data Set Curation
2.3. Model Selection and Phylogenetic Analysis
3. Results and Discussion
3.1. PPRV Genome Sequences of New Indian Isolates
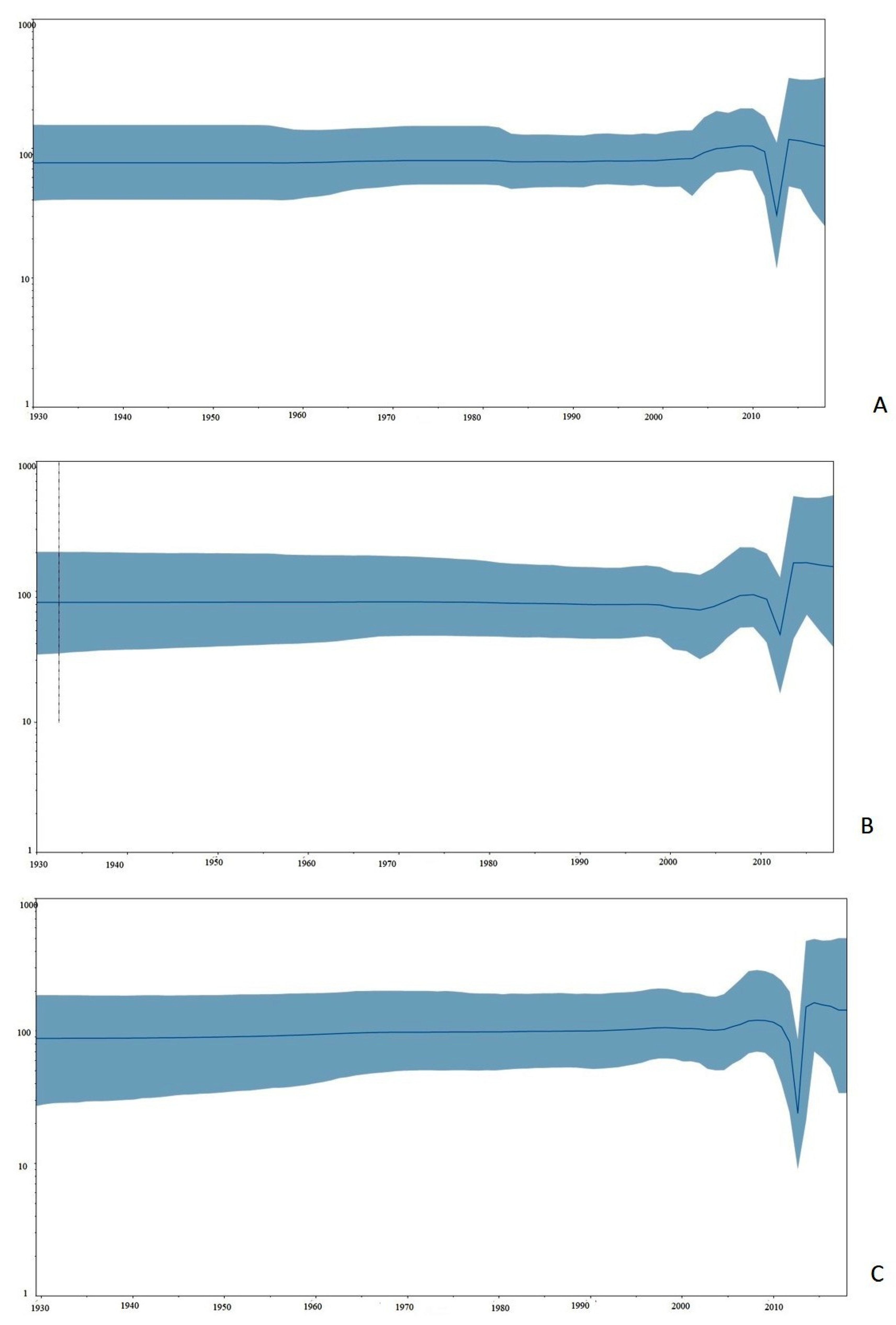
3.2. Evolutionary Rates and Lineage Divergence
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Banyard, A.C.; Parida, S.; Batten, C.; Oura, C.; Kwiatek, O.; Libeau, G. Global distribution of peste des petits ruminants virus and prospects for improved diagnosis and control. J. Gen. Virol. 2010, 91, 2885–2897. [Google Scholar] [CrossRef] [Green Version]
- Parida, S.; Muniraju, M.; Mahapatra, M.; Muthuchelvan, D.; Buczkowski, H.; Banyard, A.C. Peste des petits ruminants. Vet. Microbiol. 2015, 181, 90–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, K.S.; Babu, A.; Sundarapandian, G.; Roy, P.; Thangavelu, A.; Kumar, K.S.; Arumugam, R.; Chandran, N.D.; Muniraju, M.; Mahapatra, M.; et al. Molecular characterisation of lineage IV peste des petits ruminants virus using multi gene sequence data. Vet. Microbiol. 2014, 174, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Tounkara, K.; Kwiatek, O.; Sidibe, C.A.K.; Sery, A.; Dakouo, M.; Salami, H.; Lo, M.M.; Ba, A.; Diop, M.; Niang, M.; et al. Persistence of the historical lineage I of West Africa against the ongoing spread of the Asian lineage of peste des petits ruminants virus. Transbound. Emerg. Dis. 2021. [Google Scholar] [CrossRef]
- Mariner, J.C.; House, J.A.; Mebus, C.A.; Sollod, A.E.; Chibeu, D.; Jones, B.A.; Roeder, P.L.; Admassu, B.; van’t Klooster, G.G. Rinderpest eradication: Appropriate technology and social innovations. Science 2012, 337, 1309–1312. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.T.; Le, R.P. Further investigations into immunization of cattle against rinderpest. Onderstepoort. J. Vet. Sci. Anim. Ind. 1946, 21, 7–16. [Google Scholar]
- Anonymous. VETERINARY CORPS develops a new rinderpest vaccine. Vet. Med. 1946, 41, 153–157. [Google Scholar]
- Zhao, H.; Njeumi, F.; Parida, S.; Benfield, C.T.O. Progress towards Eradication of Peste des Petits Ruminants through Vaccination. Viruses 2021, 13, 59. [Google Scholar] [CrossRef] [PubMed]
- Furley, C.W.; Taylor, W.P.; Obi, T.U. An outbreak of peste des petits ruminants in a zoological collection. Vet. Rec. 1987, 121, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Cao, X.; Wu, J.; Dou, Y.; Meng, X.; Liu, D.; Liu, Y.; Shang, Y.; Liu, X. Epidemic and evolutionary characteristics of peste des petits ruminants virus infecting Procapra przewalskii in Western China. Infect. Genet. Evol. 2019, 75, 104004. [Google Scholar] [CrossRef] [PubMed]
- Pruvot, M.; Fine, A.E.; Hollinger, C.; Strindberg, S.; Damdinjav, B.; Buuveibaatar, B.; Chimeddorj, B.; Bayandonoi, G.; Khishgee, B.; Sandag, B.; et al. Outbreak of Peste des Petits Ruminants among Critically Endangered Mongolian Saiga and Other Wild Ungulates, Mongolia, 2016–2017. Emerg. Infect. Dis. 2020, 26, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, X.F.; Fine, A.E.; Pruvot, M.; Njeumi, F.; Walzer, C.; Kock, R.; Shiilegdamba, E. PPR virus threatens wildlife conservation. Science 2018, 362, 165–166. [Google Scholar] [CrossRef] [Green Version]
- Bailey, D.; Banyard, A.; Dash, P.; Ozkul, A.; Barrett, T. Full genome sequence of peste des petits ruminants virus, a member of the Morbillivirus genus. Virus Res. 2005, 110, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Muniraju, M.; Munir, M.; Parthiban, A.R.; Banyard, A.C.; Bao, J.; Wang, Z.; Ayebazibwe, C.; Ayelet, G.; El Harrak, M.; Mahapatra, M.; et al. Molecular Evolution of Peste des Petits Ruminants Virus. Emerg. Infect. Dis. 2014, 20, 2023–2033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diallo, A.; Taylor, W.P.; Lefevre, P.C.; Provost, A. Attenuation of a strain of rinderpest virus: Potential homologous live vaccine. Rev. Elev. Med. Vet. Pays. Trop. 1989, 42, 311–319. [Google Scholar] [PubMed]
- Adombi, C.M.; Waqas, A.; Dundon, W.G.; Li, S.; Daojin, Y.; Kakpo, L.; Aplogan, G.L.; Diop, M.; Lo, M.M.; Silber, R.; et al. Peste Des Petits Ruminants in Benin: Persistence of a Single Virus Genotype in the Country for Over 42 Years. Transbound. Emerg. Dis. 2017, 64, 1037–1044. [Google Scholar] [CrossRef]
- Benfield, C.T.O.; Hill, S.; Shatar, M.; Shiilegdamba, E.; Damdinjav, B.; Fine, A.; Willett, B.; Kock, R.; Bataille, A. Molecular epidemiology of peste des petits ruminants virus emergence in critically endangered Mongolian saiga antelope and other wild ungulates. Virus Evol. 2021, 7. [Google Scholar] [CrossRef]
- Masdooq, A.A.; Pawar, R.M.; Parthiban, A.B.; Ragavendhar, K.; Sundarapandian, G.; Thangavelu, A.; Dhinakar Raj, G. Complete Genome Sequences of Lineage IV Peste des Petits Ruminants Viruses from the Indian Subcontinent. Genome Announc. 2015, 3, e01009-15. [Google Scholar] [CrossRef] [Green Version]
- Clarke, B.; Mahapatra, M.; Friedgut, O.; Bumbarov, V.; Parida, S. Persistence of Lineage IV Peste-des-petits ruminants virus within Israel since 1993: An evolutionary perspective. PLoS ONE 2017, 12, e0177028. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A.; Lam, T.T.; Max Carvalho, L.; Pybus, O.G. Exploring the temporal structure of heterochronous sequences using TempEst (formerly Path-O-Gen). Virus Evol. 2016, 2, vew007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018, 4. [Google Scholar] [CrossRef] [Green Version]
- Baele, G.; Lemey, P.; Bedford, T.; Rambaut, A.; Suchard, M.A.; Alekseyenko, A.V. Improving the accuracy of demographic and molecular clock model comparison while accommodating phylogenetic uncertainty. Mol. Biol. Evol. 2012, 29, 2157–2167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, J.; Rambaut, A.; Pybus, O.G. Correlating viral phenotypes with phylogeny: Accounting for phylogenetic uncertainty. Infect. Genet. Evol. 2008, 8, 239–246. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Baazizi, R.; Mahapatra, M.; Clarke, B.D.; Ait-Oudhia, K.; Khelef, D.; Parida, S. Peste des petits ruminants (PPR): A neglected tropical disease in Maghreb region of North Africa and its threat to Europe. PLoS ONE 2017, 12, e0175461. [Google Scholar] [CrossRef] [PubMed]
- Sahu, A.; Wani, S.; Saminathan, M.; Rajak, K.; Sahoo, A.; Pandey, A.; Saxena, S.; Kanchan, S.; Mishra, B.; Muthuchelvan, D.; et al. Genome sequencing of an Indian peste des petits ruminants virus isolate, Izatnagar/94, and its implications for virus diversity, divergence and phylogeography. Arch. Virol. 2017, 162, 1677–1693. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Wang, Q.; Li, L.; Liu, C.; Zhang, Z.; Li, J.; Wang, S.; Wu, X.; Wang, Z. Evolutionary dynamics of recent peste des petits ruminants virus epidemic in China during 2013–2014. Virology 2017, 510, 156–164. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Data Set | Time Period | Mean Evolutionary Rate | 95% HPD Interval | Reference | |
|---|---|---|---|---|---|
| Lower | Upper | ||||
| 12 | 1976–2012 | 9.09 × 10−4 | 2.13 × 10−4 | 1.64 × 10−3 | Muniraju et al., 2014 |
| 27 | 1969–2011 | 7.8 × 10−4 | 7.3 × 10−4 | 8.4 × 10−4 | Adombi et al., 2015 |
| 37 | 1969–2015 | 7.684 ×10−4 | 7.233 ×10−4 | 8.1327 ×10−4 | Sahu et al., 2017 |
| 37 | 1969–2014 | 9.22 × 10−4 | 6.206 × 0−4 | 1.26 × 10−3 | Clarke et al., 2017 |
| 81 | 1969–2018 | 9.22 × 10−4 | 6.78 × 0−4 | 1.17 × 10−3 | Benfield et al., 2021 |
| 103 | 1969–2018 | 8.098 × 10−4 | 5.867 × 10−4 | 9.056 × 10−4 | This study |
| Set Description | Root Age | 95% HPD Interval | Number of Sequences | Time Period | |
|---|---|---|---|---|---|
| Lower | Upper | ||||
| PPRV-Common Ancestor | 1904 | 1715 | 1967 | 103 | 1969–2018 |
| Common ancestor of lineage I and III | 1924 | - | - | 9 | 1969–2018 |
| Common ancestor of lineage II and IV | 1909 | 1936 | 1954 | 94 | 1969–2018 |
| Lineage I | 1960 | 1940 | 1968 | 2 | 1969–1989 |
| Lineage II | 1957 | 1943 | 1965 | 9 | 1969–2015 |
| Lineage III | 1965 | 1944 | 1977 | 7 | 1983–2018 |
| Lineage IV | 1967 | 1948 | 1981 | 85 | 1994–2018 |
| Statistics | Observed Mean (95% CI) | Null Mean (95% CI) | p-Value |
|---|---|---|---|
| AI | 2.78 (2.77, 2.79) | 9.09 (8.26, 9.82) | 0.00 |
| PS | 29.0 (29.0, 29.0) | 61.66 (59.5, 64.0) | 0.00 |
| MC (India) | 6.0 (6.0, 6.0) | 1.23 (1.0, 2.0) | 0.009 |
| MC (Israel) | 18.0 (18.0, 18.0) | 1.81 (1.0, 3.0) | 0.009 |
| MC (China) | 14.5 (11.0, 18.0) | 2.75 (2.0, 4.0) | 0.009 |
| MC (Pakistan) | 3.0 (3.0, 3.0) | 1.0 (1.0,1.0) | 0.009 |
| MC (Ethiopia) | 2.0 (2.0, 2.0) | 1.0 (1.0,1.0) | 0.029 |
| MC (Mongolia) | 4.0 (4.0, 4.0) | 1.0 (1.0,1.0) | 0.029 |
| Gene | Evolutionary Rate (95% HPD) | Evolutionary Correlation Coefficients (95% HPD) | Relative Substitution Rate of Different Codon Position | ||
|---|---|---|---|---|---|
| 1 | 2 | 3 | |||
| N | 6.547 × 10−4 (4.691–8.444 × 10−4) | 0.917 (0.8139–1.0231) | 0.474 (0.367–0.586) | 0.283 (0.214–0.356) | 2.243 (2.119–2.361) |
| P | 6.33 × 10−4 (4.541–8.233 × 10−4) | 0.952 (0.8477–1.0538) | 0.83 (0.712–0.952) | 0.675 (0.574–0.779) | 1.494 (1.365–1.619) |
| M | 6.753 × 10−4 (4.681–8.89 × 10−4) | 0.975 (0.866–1.0822) | 0.571 (0.387–0.764) | 0.145 (0.089–0.203) | 2.284 (2.103–2.478) |
| F | 3.419 × 10−3 (2.307–4.657 × 10−3) | 1.069 (0.964–1.1745) | 0.449 (0.382–0.519) | 0.312 (0.257–0.369) | 2.230 (2.152–2.325) |
| H | 6.37 × 10−4 (4.561–8.227 × 10−4) | 0.914 (0.815–1.0206) | 0.542 (0.461–0.625) | 0.433 (0.359–0.51) | 2.026 (1.924–2.122) |
| L | 5.071 × 10−4 (3.681–6.446 × 10−4) | 0.921 (0.8178–1.0217) | 0.451 (0.405–0.497) | 0.229 (0.196–0.264) | 2.32 (2.266–2.373) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahapatra, M.; Pachauri, R.; Subramaniam, S.; Banyard, A.C.; ChandraSekar, S.; Ramakrishnan, M.A.; Njeumi, F.; Muthuchelvan, D.; Parida, S. Ongoing Assessment of the Molecular Evolution of Peste Des Petits Ruminants Virus Continues to Question Viral Origins. Viruses 2021, 13, 2144. https://doi.org/10.3390/v13112144
Mahapatra M, Pachauri R, Subramaniam S, Banyard AC, ChandraSekar S, Ramakrishnan MA, Njeumi F, Muthuchelvan D, Parida S. Ongoing Assessment of the Molecular Evolution of Peste Des Petits Ruminants Virus Continues to Question Viral Origins. Viruses. 2021; 13(11):2144. https://doi.org/10.3390/v13112144
Chicago/Turabian StyleMahapatra, Mana, Richa Pachauri, Saravanan Subramaniam, Ashley C. Banyard, Shanmugam ChandraSekar, Muthannan Andavar Ramakrishnan, Felix Njeumi, Dhanavelu Muthuchelvan, and Satya Parida. 2021. "Ongoing Assessment of the Molecular Evolution of Peste Des Petits Ruminants Virus Continues to Question Viral Origins" Viruses 13, no. 11: 2144. https://doi.org/10.3390/v13112144
APA StyleMahapatra, M., Pachauri, R., Subramaniam, S., Banyard, A. C., ChandraSekar, S., Ramakrishnan, M. A., Njeumi, F., Muthuchelvan, D., & Parida, S. (2021). Ongoing Assessment of the Molecular Evolution of Peste Des Petits Ruminants Virus Continues to Question Viral Origins. Viruses, 13(11), 2144. https://doi.org/10.3390/v13112144