Expression Profiles of Exosomal MicroRNAs from HEV- and HCV-Infected Blood Donors and Patients: A Pilot Study

Abstract
1. Introduction
2. Materials and Methods
2.1. Blood Donor and Patient Samples
2.2. Exosome Precipitation
2.3. Density Gradient Ultracentrifugation of Enriched EXs
2.4. Nanoparticle Tracking Analysis (NTA)
2.5. Protein Extraction
2.6. Dot Blots
2.7. Automated Protein Gel Electrophoresis
2.8. Nucleic Acid Extraction
2.9. microRNA Analysis
2.10. Statistics
3. Results and Discussion
3.1. Exosome Preparation and Purification
3.2. Determination of EX-Containing Fractions
3.3. Exosomal microRNA Analysis
3.3.1. Comparison of Blood Donor and Patient Sets of Samples
3.3.2. Pre- and Post-Treatment Samples of the HEV/HCV Co-Infected Patient
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Anthoniou, A.; Arab, A.A.T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the international society for extracellular vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles. 2018, 7, 1535750. [Google Scholar] [PubMed]
- van Balkom, B.W.M.; Eisele, A.S.; Pegtel, D.M.; Bervoets, S.; Verhaar, M.C. Quantitative and qualitative analysis of small RNAs in human endothelial cells and exosomes provides insights into localized RNA processing, degradation and sorting. J. Extracell. Vesicles 2015, 4, 26760. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Samuel, M.; Kumar, S.; Mathivanan, S. Ticket to a bubble ride: Cargo sorting into exosomes and extracellular vesicles. Biochim. Biophys. Acta Proteins Proteom. 2019, 1867, 140203. [Google Scholar] [CrossRef]
- Villarroya-Beltri, C.; Gutierrez-Vazquez, C.; Sanchez-Cabo, F.; Perez- Hernandez, D.; Vazquez, J.; Martin-Cofreces, N.; Martinez-Herrera, D.J.; Pascual-Montano, A.; Mittelbrunn, M.; Sánchez-Madrid, F. Sumoylated hnRNPA2B1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat. Commun. 2013, 4, 2980. [Google Scholar] [CrossRef]
- Schorey, J.S.; Cheng, Y.; Singh, P.P.; Smith, V.L. Exosomes and other extracellular vesicles in host-pathogen interactions. EMBO Rep. 2015, 16, 24–43. [Google Scholar] [CrossRef]
- Petrik, J. Immunomodulatory effects of exosomes produced by virus-infected cells. Transfus. Apher. Sci. 2016, 55, 84–91. [Google Scholar] [CrossRef]
- Shen, J.; Huang, C.-K.; Yu, H.; Shen, B.; Zhang, Y.; Liang, Y.; Li, Z.; Feng, X.; Zhao, J.; Duan, L.; et al. The role of exosomes in hepatitis, liver cirrhosis and hepatocellular carcinoma. J. Cell. Mol. Med. 2017, 21, 986–992. [Google Scholar] [CrossRef]
- Ramakrishnaiah, V.; Thumann, C.; Fofana, I.; Habersetzer, F.; Pan, Q.; de Ruiter, P.E.; Willemsen, R.; Demmers, J.A.; Stalin Raj, V.; Jenster, G.; et al. Exosome-mediated transmission of hepatitis C virus between human hepatoma Huh7.5 cells. Proc. Natl. Acad. Sci. USA 2013, 110, 13109–13113. [Google Scholar] [CrossRef]
- Malik, M.A.; Mirza, J.I.A.; Umar, M.; Manzoor, S. CD81 + Exosomes Play a Pivotal Role in the Establishment of Hepatitis C Persistent Infection and Contribute Toward the Progression of Hepatocellular Carcinoma. Viral Immunol. 2019, 32, 453–462. [Google Scholar] [CrossRef]
- Feng, Z.; Hirai-Yuki, A.; McKnight, K.L.; Lemon, S.M. Naked viruses that aren’t always naked: Quasi-enveloped agents of acute hepatitis. Annu. Rev. Virol. 2014, 1, 539–560. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Ambardekar, C.; Lu, Y.; Feng, Z. Distinct Entry Mechanisms for Nonenveloped and Quasi-Enveloped Hepatitis E Viruses. J. Virol. 2016, 90, 4232–4242. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, M.C.; Imlach, S.N.; Abravanel, F.; Ramalingam, S.; Johannessen, I.; Petrik, J.; Fraser, A.R.; Campbell, J.D.; Bramley, P.; Dalton, H.R.; et al. Sofosbuvir and Daclatasvir Anti–Viral Therapy Fails to Clear HEV Viremia and Restore Reactive T Cells in a HEV/HCV Co-Infected Liver Transplant Recipient. Gastroenterology 2017, 152, 300–301. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, X. miRDB: An online database for prediction of functional microRNA targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef]
- Liang, J.J.; Wang, J.Y.; Zhang, T.J.; An, G.S.; Ni, J.H.; Jia, H.T.; Li, S.Y. MiR-509-3-5p-NONHSAT112228.2 Axis Regulates p21 and Suppresses Proliferation and Migration of Lung Cancer Cells. Curr. Top. Med. Chem. 2020, 20, 835–846. [Google Scholar] [CrossRef]
- Zhou, Y.; Xia, Z.; Cheng, Z.; Xu, G.; Yang, X.; Liu, S.; Zhu, Y. Inducible microRNA-590-5p inhibits host antiviral response by targeting the soluble interleukin-6 (IL6) receptor. J. Biol. Chem. 2018, 293, 18168–18179. [Google Scholar] [CrossRef]
- Mekky, R.Y.; El-Ekiaby, N.M.; Hamza, M.T.; Elemam, N.M.; El-Sayed, M.; Esmat, G.; Abdelaziz, A.I. Mir-194 is a hepatocyte gate keeper hindering HCV entry through targeting CD81 receptor. J. Infect. 2015, 70, 78–87. [Google Scholar] [CrossRef]
- Wang, R.Y.; Weng, K.F.; Huang, Y.C.; Chen, C.J. Elevated expression of circulating miR876-5p is a specific response to severe EV71 infections. Sci. Rep. 2016, 6, 24149. [Google Scholar] [CrossRef]
- Eilam-Frenkel, B.; Naaman, H.; Brkic, G.; Veksler-Lublinsky, I.; Rall, G.; Shemer-Avni, Y.; Gopas, J. MicroRNA 146-5p, miR-let-7c-5p, miR-221 and miR-345-5p are differentially expressed in Respiratory Syncytial Virus (RSV) persistently infected HEp-2 cells. Virus Res. 2018, 251, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, M.; Zeremski, M.; Talal, A.H.; Ginwala, R.; Elrod, E.; Grakoui, A.; Li, Q.G.; Philip, R.; Khan, Z.K.; Jain, P. IFN-α-Induced Downregulation of miR-221 in Dendritic Cells: Implications for HCV Pathogenesis and Treatment. J. Interf. Cytokine Res. 2015, 35, 698–709. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Xu, C.; Fang, S.; Zhao, P.; Wang, Y.; Liu, H.; Yuan, W.; Qi, Z. Exosomal MicroRNAs Derived From Umbilical Mesenchymal Stem Cells Inhibit Hepatitis C Virus Infection. Stem Cells Transl. Med. 2016, 5, 1190–1203. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Ouyang, Y.; Che, J.; Li, X.; Zhao, Y.; Yang, K.; Zhao, X.; Chen, Y.; Fan, C.; Yuan, W. Potential Value of miR-221/222 as Diagnostic, Prognostic, and Therapeutic Biomarkers for Diseases. Front. Immunol. 2017, 8, 56. [Google Scholar] [CrossRef] [PubMed]
- Khanizadeh, S.; Ravanshad, M.; Hosseini, S.Y.; Davoodian, P.; Almasian, M.; Khanlari, Z. The effect of the hepatitis C virus (HCV) NS3 protein on the expression of miR-150, miR-199a, miR-335, miR-194 and miR-27a. Microb. Pathog. 2017, 110, 688–693. [Google Scholar] [CrossRef] [PubMed]
- El-Diwany, R.; Wasilewski, L.N.; Witwer, K.W.; Bailey, J.R.; Page, K.; Ray, S.C.; Cox, A.L.; Thomas, D.L.; Balagopal, A. Acute hepatitis C virus infection induces consistent changes in circulating microRNAs that are associated with nonlytic hepatocyte release. J. Virol. 2015, 89, 9454–9464. [Google Scholar] [CrossRef] [PubMed]
- Zekri, A.N.; Youssef, A.S.; El-Desouky, E.D.; Ahmed, O.S.; Lotfy, M.M.; Nassar, A.A.; Bahnassey, A.A. Serum microRNA panels as potential biomarkers for early detection of hepatocellular carcinoma on top of HCV infection. Tumor Biol. 2016, 37, 12273–12286. [Google Scholar] [CrossRef]
- Nasser, M.Z.; Zayed, N.A.; Mohamed, A.M.; Attia, D.; Esmat, G.; Khairy, A. Circulating microRNAs (miR-21, miR-223, miR-885-5p) along the clinical spectrum of HCV-related chronic liver disease in Egyptian patients. Arab J. Gastroenterol. 2019, 20, 198–204. [Google Scholar] [CrossRef]
- Li, Q.; Lowey, B.; Sodroski, C.; Krishnamurthy, S.; Alao, H.; Cha, H.; Chiu, S.; El-Diwany, R.; Ghany, M.G.; Liang, T.J. Cellular microRNA networks regulate host dependency of hepatitis C virus infection. Nat. Commun. 2017, 8, 1789. [Google Scholar] [CrossRef]
- Boštjančič, E.; Bandelj, E.; Luzar, B.; Poljak, M.; Glavač, D. Hepatic expression of miR-122, miR-126, miR-136 and miR-181a and their correlation to histopathological and clinical characteristics of patients with hepatitis C. J. Viral Hepat. 2015, 22, 146–157. [Google Scholar] [CrossRef]
- Harms, D.; Choi, M.; Allers, K.; Wang, B.; Pietsch, H.; Papp, C.-P.; Hanisch, L.; Kurreck, J.; Hofmann, J.; Bock, C.T. Specific circulating microRNAs during hepatitis E infection can serve as indicator for chronic hepatitis E. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Santangelo, L.; Bordoni, V.; Montaldo, C.; Cimini, E.; Zingoni, A.; Battistelli, C.; D’Offizi, G.; Capobianchi, M.R.; Santoni, A.; Tripodi, M.; et al. Hepatitis C virus direct-acting antivirals therapy impacts on extracellular vesicles microRNAs content and on their immunomodulating properties. Liver Int. 2018, 38, 1741–1750. [Google Scholar] [CrossRef] [PubMed]
- Morishita, A.; Yoneyama, H.; Iwama, H.; Fujita, K.; Watanabe, M.; Hirose, K.; Tadokoro, T.; Oura, K.; Sakamoto, T.; Mimura, S.; et al. Circulating microRNA-636 is associated with the elimination of hepatitis C virus by ombitasvir/paritaprevir/ritonavir. Oncotarget 2018, 9, 32054–32062. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wong, Q.W.; Lung, R.W.; Law, P.T.; Lai, P.B.; Chan, K.Y.; To, K.F.; Wong, N. MicroRNA-223 is commonly repressed in hepatocellular carcinoma and potentiates expression of Stathmin1. Gastroenterology 2008, 135, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Lin, C.C.; Hsieh, W.L.; Lai, H.W.; Tsai, C.H.; Cheng, Y.W. MicroRNA expression profiling in PBMCs: A potential diagnostic biomarker of chronic hepatitis C. Dis. Markers 2014, 2014, 367157. [Google Scholar] [CrossRef] [PubMed]
- Ashmawy, A.M.; Elgeshy, K.M.; Abdel Salam, E.T.; Ghareeb, M.; Kobaisi, M.H.; Amin, H.A.A.; Sharawy, S.K.; Abdel Wahab, A.H.A. Crosstalk between liver-related microRNAs and Wnt/β-catenin pathway in hepatocellular carcinoma patients. Arab J. Gastroenterol. 2017, 18, 144–150. [Google Scholar] [CrossRef] [PubMed]
- El-Abd, N.E.; Fawzy, N.A.; El-Sheikh, S.M.; Soliman, M.E. Circulating miRNA-122, miRNA-199a, and miRNA-16 as Biomarkers for Early Detection of Hepatocellular Carcinoma in Egyptian Patients with Chronic Hepatitis C Virus Infection. Mol. Diagn. Ther. 2015, 19, 213–220. [Google Scholar] [CrossRef]
- Cheng, J.C.; Yeh, Y.J.; Tseng, C.P.; Hsu, S.D.; Chang, Y.L.; Sakamoto, N.; Huang, H.D. Let-7b is a novel regulator of hepatitis C virus replication. Cell. Mol. Life Sci. 2012, 69, 2621–2633. [Google Scholar] [CrossRef]
- Conrad, K.D.; Niepmann, M. The role of microRNAs in hepatitis C virus RNA replication. Arch. Virol. 2014, 159, 849–862. [Google Scholar] [CrossRef]
- Liao, T.L.; Hsieh, S.L.; Chen, Y.M.; Chen, H.H.; Liu, H.J.; Lee, H.C.; Chen, D.Y. Rituximab May Cause Increased Hepatitis C Virus Viremia in Rheumatoid Arthritis Patients Through Declining Exosomal MicroRNA-155. Arthritis Rheumatol. 2018, 70, 1209–1219. [Google Scholar] [CrossRef]
- Bandiera, S.; Pernot, S.; El Saghire, H.; Durand, S.C.; Thumann, C.; Crouchet, E.; Ye, T.; Fofana, I.; Oudot, M.A.; Barths, J.; et al. Hepatitis C Virus-Induced Upregulation of MicroRNA miR-146a-5p in Hepatocytes Promotes Viral Infection and Deregulates Metabolic Pathways Associated with Liver Disease Pathogenesis. J. Virol. 2016, 90, 6387–6400. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Daucher, M.; Armistead, D.; Russell, R.; Kottilil, S. MicroRNA expression profiling in HCV-infected human hepatoma cells identifies potential anti-viral targets induced by interferon-α. PLoS ONE 2013, 8, e55733. [Google Scholar] [CrossRef] [PubMed]
- Tsubota, A.; Mogushi, K.; Aizaki, H.; Miyaguchi, K.; Nagatsuma, K.; Matsudaira, H.; Kushida, T.; Furihata, T.; Tanaka, H.; Matsuura, T. Involvement of MAP3K8 and miR-17-5p in poor virologic response to interferon-based combination therapy for chronic hepatitis C. PLoS ONE 2014, 9, e97078. [Google Scholar] [CrossRef] [PubMed]
- Brochado-Kith, Ó.; Gómez Sanz, A.; Real, L.M.; Crespo García, J.; Ryan Murúa, P.; Macías, J.; Cabezas González, J.; Troya, J.; Pineda, J.A.; Arias Loste, M.T.; et al. MicroRNA Profile of HCV Spontaneous Clarified Individuals, Denotes Previous HCV Infection. J. Clin. Med. 2019, 8, 849. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, J.; Wang, H.; Shi, J.; Wu, K.; Liu, S.; Liu, Y.; Wu, J. HCV-induced miR-21 contributes to evasion of host immune system by targeting MyD88 and IRAK1. PLoS Pathog. 2013, 9, e1003248. [Google Scholar] [CrossRef]
- Clément, S.; Sobolewski, C.; Gomes, D.; Rojas, A.; Goossens, N.; Conzelmann, S.; Calo, N.; Negro, F.; Foti, M. Activation of the oncogenic miR-21-5p promotes HCV replication and steatosis induced by the viral core 3a protein. Liver Int. 2019, 39, 1226–1236. [Google Scholar] [CrossRef]
- Bose, M.; Bhattacharyya, S.N. Target-dependent biogenesis of cognate microRNAs in human cells. Nat. Commun. 2016, 7, 12200. [Google Scholar] [CrossRef]
- Wilson, J.A.; Zhang, C.; Huys, A.; Richardson, C.D. Human Ago2 is required for efficient microRNA 122 regulation of hepatitis C virus RNA accumulation and translation. J. Virol. 2011, 85, 2342–2350. [Google Scholar] [CrossRef]
- Shimakami, T.; Yamane, D.; Jangra, R.K.; Kempf, B.J.; Spaniel, C.; Barton, D.J.; Lemon, S.M. Stabilization of hepatitis C virus RNA by an Ago2-miR-122 complex. Proc. Natl. Acad. Sci. USA 2012, 109, 941–946. [Google Scholar] [CrossRef]
- Haldipur, B.; Bhukya, P.L.; Arankalle, V.; Lole, K. Positive regulation of hepatitis E virus replication by microRNA-122. J. Virol. 2018, 92, e01999-17. [Google Scholar] [CrossRef]
- Morita, K.; Taketomi, A.; Shirabe, K.; Umeda, K.; Kayashima, H.; Ninomiya, M.; Uchiyama, H.; Soejima, Y.; Maehara, Y. Clinical significance and potential of hepatic microRNA-122 expression in hepatitis C. Liver Int. 2011, 31, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Sarasin-Filipowicz, M.; Krol, J.; Markiewicz, I.; Heim, M.H.; Filipowicz, W. Decreased levels of microRNA miR-122 in individuals with hepatitis C responding poorly to interferon therapy. Nat. Med. 2009, 15, 31–33. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.; Fan, Z.; Chen, H.; He, P.; Li, Y.; Zhang, Q.; Ke, C. Serum and exosomal miR-122 and miR-199a as a biomarker to predict therapeutic efficacy of hepatitis C patients. J. Med. Virol. 2017, 89, 1597–1605. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Abbreviation | Definition of Groups |
|---|---|
| DNA | D = donor (D1–4); N = negative; A = Taqman card A |
| DNB | D = donor (D1–4); N = negative; B = Taqman card B |
| DEA | D = donor (D1–4); E = HEV-positive; A = Taqman card A |
| DEB | D = donor (D1–4); E = HEV-positive; B = Taqman card B |
| DCA | D = donor (D1–4); C = HCV-positive; A = Taqman card A |
| DCB | D = donor (D1–4); C = HCV-positive; B = Taqman card B |
| PEA | P = patient (P1.1, 1.2, 2, 4); E = HEV-positive; A = Taqman card A |
| PEB | P = patient (P1.1, 1.2, 2, 4); E = HEV-positive; B = Taqman card B |
| PCA | P = patient (P1.1, 3); C = HCV-positive; A = Taqman card A |
| PCB | P = patient (P1.1, 3); C = HCV-positive; B = Taqman card B |
| Sample | Concentration (Particles/mL) +/− SD | Mean Size nm +/− SD (nm) | Merged Data Mean (nm) |
|---|---|---|---|
| HCV BD 1 | 5.62 × 106 +/− 2.58 × 106 | 118.8 +/− 10.5 | 110.3 |
| HCV BD 2 | 9.20 × 108 +/− 1.53 × 106 | 146.2 +/− 4.7 | 144.8 |
| HCV BD 3 | 3.70 × 108 +/− 2.94 × 107 | 111.0 +/− 2.3 | 110.7 |
| HCV BD 4 | 6.16 × 108 +/− 7.12 × 107 | 106.3 +/− 2.8 | 106.4 |
| Negative BD 1 | 3.16 × 107 +/− 6.85 × 106 | 168.6 +/− 7.9 | 169 |
| Negative BD 2 | 5.84 × 108 +/− 4.43 × 107 | 100.6 +/− 2.0 | 100.3 |
| Negative BD 3 | 9.71 × 108 +/− 2.65 × 107 | 142.6 +/− 1.2 | 142.6 |
| Negative BD 4 | 9.61 × 108 +/− 3.73 × 107 | 154.9 +/− 2.7 | 154.9 |
| HEV BD 1 | 1.08 × 107 +/− 5.3 × 106 | 115.9 +/− 32.7 | 144.2 |
| HEV BD 2 | 1.94 × 107 +/− 6.63 × 106 | 137.9 +/− 22.0 | 125.7 |
| HEV BD 3 | 1.77 × 107 +/− 2.33 × 106 | 236.9 +/− 53.5 | 222.9 |
| HEV BD 4 | 1.40 × 107 +/− 7.93 × 105 | 175.4 +/− 2.1 | 175.6 |
| Patient 1.1 | 2.17 × 108 +/− 2.27 × 107 | 121.8 +/− 3.8 | 121.3 |
| Patient 1.2 | 7.7 × 106 +/− 3.36 × 106 | 113.7 +/− 75.1 | 166.6 |
| Patient 2 | 2.15 × 107 +/− 4.17 × 106 | 229.3 +/− 30.4 | 218.4 |
| Patient 3 | 2.05 × 107 +/− 6.07 × 106 | 182.9 +/− 24.9 | 168.2 |
| Patient 4 | 3.27 × 107 +/− 4.77 × 106 | 242.1 +/− 101.1 | 242.1 |
| Experiment | Hsa-miR- | ↑Up or ↓Down Regulation | p = (Global Normalisation) * | p = (ath-159a Normalisation) * |
|---|---|---|---|---|
| DEA/DNA | 526b | ↓DEA | 0.034 | 0.020 |
| DEB/DNB | 335 | ↑DEB | 0.013 | 0.022 |
| DCA/DNA | 194 | ↑DCA | 0.006 | 0.002 |
| 509-3-5p | ↑DCA | 0.010 | 0.041 | |
| 627 | ↓DCA | 0.007 | 0.026 | |
| DCB/DNB | -- | |||
| DEA/DCA | 885-5p | ↑DCA | 0.004 | 0.005 |
| 365 | ↑DCA | 0.012 | 0.013 | |
| 374-5p | ↑DCA | 0.032 | 0.036 | |
| DEB/DCB | 659 | ↑DCB | 0.003 | 0.003 |
| 32 * | ↑DCB | 0.019 | 0.011 | |
| DCA + DEA/DNA | 194 | ↑DCA + DEA | 0.011 | 0.009 |
| 509-3-5p | ↑DCA + DEA | 0.006 | 0.023 | |
| 654-3p | ↓DCA + DEA | 0.007 | 0.017 | |
| DCB + DEB/DNB | 335 * | ↑DCB + DEB | 0.021 | 0.017 |
| 628-3p | ↑ DCB + DEB | 0.017 | 0.040 | |
| 151-3p | ↑ DCB + DEB | 0.021 | 0.031 | |
| Let-7i * | ↓ DCB + DEB | 0.026 | 0.034 | |
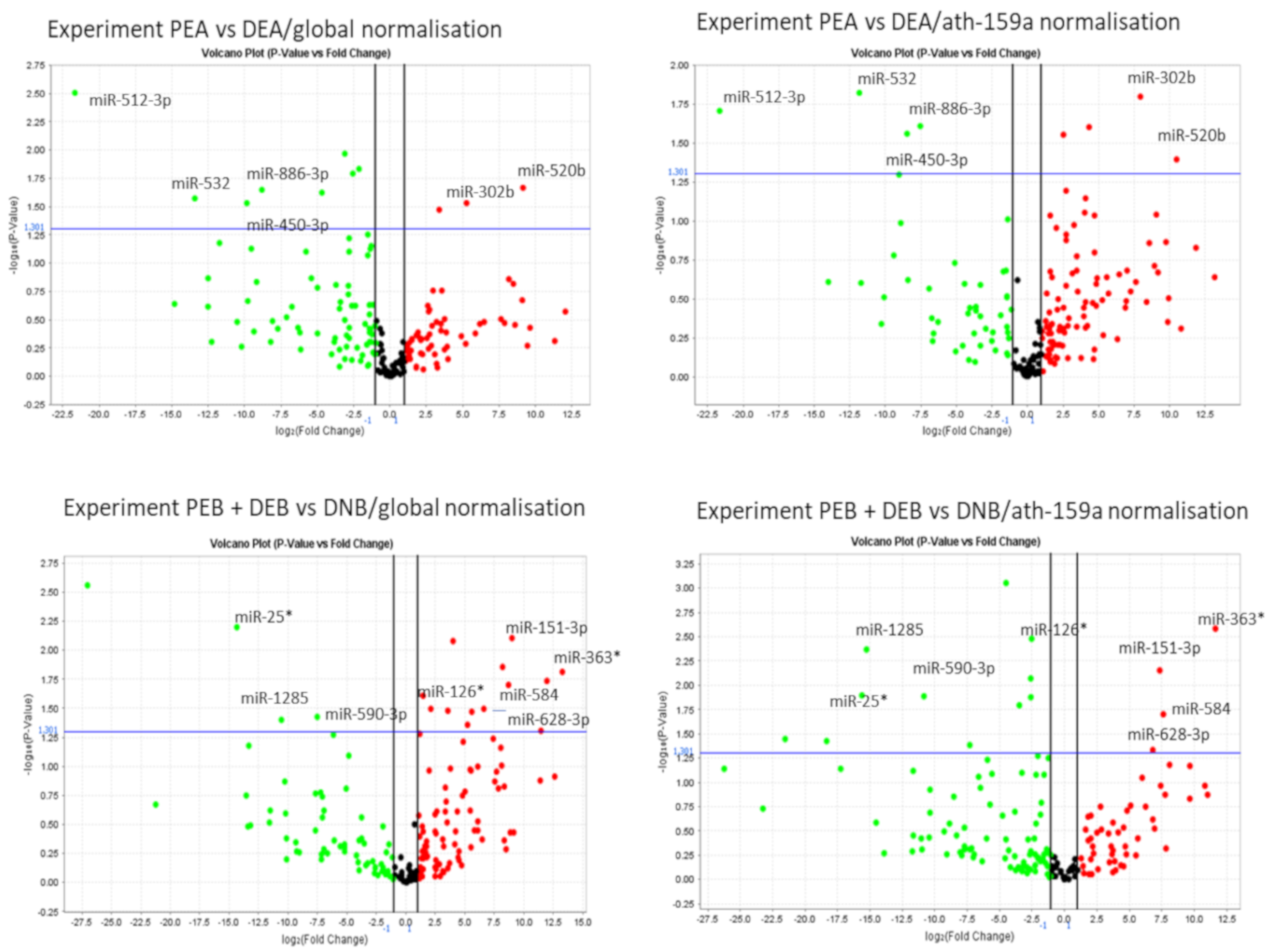
| PEA/DEA | 512-3p | ↓PEA | 0.020 | 0.003 |
| 532 | ↓PEA | 0.015 | 0.027 | |
| 450-3b | ↓PEA | 0.028 | 0.030 | |
| 886-3p | ↓PEA | 0.025 | 0.023 | |
| 302b | ↑PEA | 0.016 | 0.029 | |
| 520b | ↑PEA | 0.041 | 0.021 | |
| PEB/DEB | 335 * | ↓PEB | 0.019 | 0.014 |
| PCA/DCA | 365 | ↓PCA | 0.045 | 0.007 |
| 509-3-5p | ↓PCA | 0.030 | 0.045 | |
| PCB/DCB | 1285 | ↑PCB | 0.046 | 0.045 |
| PEA + DEA/DNA | 509-3-5p | ↑PEA + DEA | 0.002 | 0.027 |
| 526b | ↓PEA + DEA | 0.010 | 0.011 | |
| 548a/b | ↓PEA + DEA | 0.011(b) | 0.042(a) | |
| 590-5p | ↓PEA + DEA | 0.040 | 0.007 | |
| PEB + DEB/DNB | 25 * | ↓PEB + DEB | 0.006 | 0.013 |
| 1285 | ↓PEB + DEB | 0.040 | 0.004 | |
| 590-3p | ↓PEB + DEB | 0.038 | 0.013 | |
| 126 * | ↑PEB + DEB | 0.024 | 0.003 | |
| 628-3p | ↑PEB + DEB | 0.032 | 0.047 | |
| 584 | ↑PEB + DEB | 0.020 | 0.020 | |
| 151-3p | ↑PEB + DEB | 0.008 | 0.007 | |
| 363 * | ↑PEB + DEB | 0.015 | 0.003 | |
| PCA + DCA/DNA | 221 | ↓PCA + DCA | 0.013 | 0.041 |
| 590-5p | ↓PCA + DCA | 0.027 | 0.024 | |
| 509-3-5p | ↑PCA + DCA↓ | 0.013 | 0.050 | |
| PCB + DCB/DNB | --- | |||
| PEA + PCA/DEA + DCA | 512-3p | ↓PEA + PCA | 0.005 | 0.001 |
| 105 | ↓PEA + PCA | 0.050 | 0.032 | |
| 874 | ↓PEA + PCA | 0.038 | 0.042 | |
| 876-5p | ↑PEA + PCA | 0.007 | 0.019 | |
| PEB + PCB/DEB + DCB | 609 | ↓PEB + PCB | 0.019 | 0.019 |
| 590-3p | ↓PEB + PCB | 0.036 | 0.043 | |
| 614 | ↑PEB + PCB | 0.001 | 0.046 |
| microRNAs Targeting HCV 3′UTR * | microRNAs Targeting HEV 3′UTR * | ||||
|---|---|---|---|---|---|
| hsa-miR- | Frequency of Target Sequence ** | Amplification in P1.1 and P1.2 | hsa-miR- | Frequency of Target Sequence ** | Amplification in P1.1 and P1.2 |
| 122 | 33 | ↓in P1.2 | 298 | 9 | Not amplified in either |
| 505-5p | 17 | Failed to amplify in P1.2. Not included | 539-5p | 6 | 539 no significant change |
| 651-3p | 14 | Not amplified in either | 328-5p | 5 | 328 no significant change |
| 548 family | 9 | 548c ↓ in P1.2 | 548 family | 4 | 548c ↓ in P1.2 |
| 1252-5p | 8 | 1252 no significant change | |||
| 939-5p | 4 | 939 no significant change | |||
| 509-3/5p | 4 | No significant change | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McGowan, K.; Simpson, K.J.; Petrik, J. Expression Profiles of Exosomal MicroRNAs from HEV- and HCV-Infected Blood Donors and Patients: A Pilot Study. Viruses 2020, 12, 833. https://doi.org/10.3390/v12080833
McGowan K, Simpson KJ, Petrik J. Expression Profiles of Exosomal MicroRNAs from HEV- and HCV-Infected Blood Donors and Patients: A Pilot Study. Viruses. 2020; 12(8):833. https://doi.org/10.3390/v12080833
Chicago/Turabian StyleMcGowan, Karen, Kenneth J. Simpson, and Juraj Petrik. 2020. "Expression Profiles of Exosomal MicroRNAs from HEV- and HCV-Infected Blood Donors and Patients: A Pilot Study" Viruses 12, no. 8: 833. https://doi.org/10.3390/v12080833
APA StyleMcGowan, K., Simpson, K. J., & Petrik, J. (2020). Expression Profiles of Exosomal MicroRNAs from HEV- and HCV-Infected Blood Donors and Patients: A Pilot Study. Viruses, 12(8), 833. https://doi.org/10.3390/v12080833