Human Endogenous Retrovirus K in Cancer: A Potential Biomarker and Immunotherapeutic Target

 ,
,
Abstract
1. Introduction
2. HERVs: Classification and Genome
2.1. Nomenclature and Classification
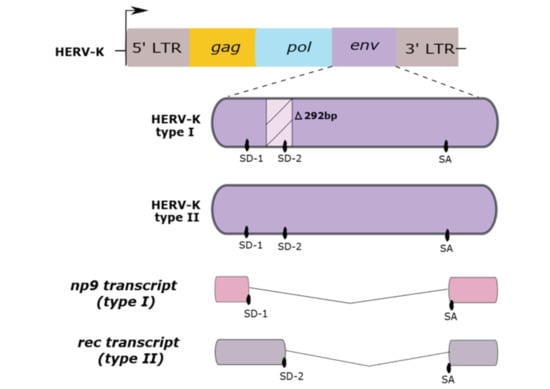
2.2. Genome Structure
2.3. HERV-K Oncoproteins
2.3.1. Rec
2.3.2. Np9
3. HERV-K in Cancer Immunotherapy
3.1. Melanoma
3.2. Breast Cancer
3.3. Prostate Cancer
4. Other Cancers
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bannert, N.; Kurth, R. Retroelements and the human genome: New perspectives on an old relation. Proc. Natl. Acad. Sci. USA 2004, 101, 14572–14579. [Google Scholar] [CrossRef] [PubMed]
- Hohn, O.; Hanke, K.; Bannert, N. HERV-K(HML-2), the Best Preserved Family of HERVs: Endogenization, Expression, and Implications in Health and Disease. Front. Oncol. 2013, 3, 246. [Google Scholar] [CrossRef] [PubMed]
- Grandi, N.; Tramontano, E. Human Endogenous Retroviruses Are Ancient Acquired Elements Still Shaping Innate Immune Responses. Front. Immunol. 2018, 9, 2039. [Google Scholar] [CrossRef] [PubMed]
- Magiorkinis, G.; Hurst, T.P. Activation of the innate immune response by endogenous retroviruses. J. Gen. Virol. 2015, 96, 1207–1218. [Google Scholar] [CrossRef]
- Blond, J.-L.; Lavillette, D.; Cheynet, V.; Bouton, O.; Oriol, G.; Chapel-Fernandes, S.; Mandrand, B.; Mallet, F.; Cosset, F.-L. An Envelope Glycoprotein of the Human Endogenous Retrovirus HERV-W Is Expressed in the Human Placenta and Fuses Cells Expressing the Type D Mammalian Retrovirus Receptor. J. Virol. 2000, 74, 3321–3329. [Google Scholar] [CrossRef] [PubMed]
- Groh, S.; Schotta, G. Silencing of endogenous retroviruses by heterochromatin. Cell. Mol. Life Sci. 2017, 74, 2055–2065. [Google Scholar] [CrossRef] [PubMed]
- Ishak, C.A.; Classon, M.; De Carvalho, D.D. Deregulation of Retroelements as an Emerging Therapeutic Opportunity in Cancer. Trends Cancer 2018, 4, 583–597. [Google Scholar] [CrossRef]
- Goering, W.; Ribarska, T.; Schulz, W.A. Selective changes of retroelement expression in human prostate cancer. Carcinogenesis 2011, 32, 1484–1492. [Google Scholar] [CrossRef]
- Schulz, W.A.; Steinhoff, C.; Florl, A.R. Methylation of Endogenous Human Retroelements in Health and Disease. In DNA Methylation: Development, Genetic Disease and Cancer; Springer: Berlin/Heidelberg, Germany, 2006; Volume 310, pp. 211–250. ISBN 3540311807. [Google Scholar]
- Misiak, B.; Ricceri, L.; Sąsiadek, M.M. Transposable Elements and Their Epigenetic Regulation in Mental Disorders: Current Evidence in the Field. Front. Genet. 2019, 10, 580. [Google Scholar] [CrossRef] [PubMed]
- Hurst, T.; Magiorkinis, G. Epigenetic Control of Human Endogenous Retrovirus Expression: Focus on Regulation of Long-Terminal Repeats (LTRs). Viruses 2017, 9, 130. [Google Scholar] [CrossRef]
- Bannert, N.; Hofmann, H.; Block, A.; Hohn, O. HERVs New Role in Cancer: From Accused Perpetrators to Cheerful Protectors. Front. Microbiol. 2018, 9, 178. [Google Scholar] [CrossRef]
- Loo Yau, H.; Ettayebi, I.; De Carvalho, D.D. The Cancer Epigenome: Exploiting Its Vulnerabilities for Immunotherapy. Trends Cell Biol. 2019, 29, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Attermann, A.S.; Bjerregaard, A.M.; Saini, S.K.; Grønbæk, K.; Hadrup, S.R. Human endogenous retroviruses and their implication for immunotherapeutics of cancer. Ann. Oncol. 2018, 29, 2183–2191. [Google Scholar] [CrossRef] [PubMed]
- Tatkiewicz, W.; Dickie, J.; Bedford, F.; Jones, A.; Atkin, M.; Kiernan, M.; Maze, E.A.; Agit, B.; Farnham, G.; Kanapin, A.; et al. Characterising a human endogenous retrovirus(HERV)-derived tumour-associated antigen: Enriched RNA-Seq analysis of HERV-K(HML-2) in mantle cell lymphoma cell lines. Mob. DNA 2020, 11, 9. [Google Scholar] [CrossRef] [PubMed]
- Wang-Johanning, F.; Radvanyi, L.; Rycaj, K.; Plummer, J.B.; Yan, P.; Sastry, K.J.; Piyathilake, C.J.; Hunt, K.K.; Johanning, G.L. Human Endogenous Retrovirus K Triggers an Antigen-Specific Immune Response in Breast Cancer Patients. Cancer Res. 2008, 68, 5869–5877. [Google Scholar] [CrossRef]
- Stoye, J.P. Studies of endogenous retroviruses reveal a continuing evolutionary saga. Nat. Rev. Microbiol. 2012, 10, 395–406. [Google Scholar] [CrossRef]
- Martin, M.A.; Bryan, T.; Rasheed, S.; Khan, A.S. Identification and cloning of endogenous retroviral sequences present in human DNA. Proc. Natl. Acad. Sci. USA 1981, 78, 4892–4896. [Google Scholar] [CrossRef]
- Garcia-Montojo, M.; Doucet-O’Hare, T.; Henderson, L.; Nath, A. Human endogenous retrovirus-K (HML-2): A comprehensive review. Crit. Rev. Microbiol. 2018, 44, 715–738. [Google Scholar] [CrossRef]
- Gifford, R.J.; Blomberg, J.; Coffin, J.M.; Fan, H.; Heidmann, T.; Mayer, J.; Stoye, J.; Tristem, M.; Johnson, W.E. Nomenclature for endogenous retrovirus (ERV) loci. Retrovirology 2018, 15, 59. [Google Scholar] [CrossRef]
- Grandi, N.; Cadeddu, M.; Pisano, M.P.; Esposito, F.; Blomberg, J.; Tramontano, E. Identification of a novel HERV-K(HML10): Comprehensive characterization and comparative analysis in non-human primates provide insights about HML10 proviruses structure and diffusion. Mob. DNA 2017, 8, 15. [Google Scholar] [CrossRef]
- Pisano, M.P.; Grandi, N.; Cadeddu, M.; Blomberg, J.; Tramontano, E. Comprehensive Characterization of the Human Endogenous Retrovirus HERV-K(HML-6) Group: Overview of Structure, Phylogeny, and Contribution to the Human Genome. J. Virol. 2019, 93. [Google Scholar] [CrossRef]
- Magin, C.; Löwer, R.; Löwer, J. cORF and RcRE, the Rev/Rex and RRE/RxRE Homologues of the Human Endogenous Retrovirus Family HTDV/HERV-K. J. Virol. 1999, 73, 9496–9507. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, R.P.; Wildschutte, J.H.; Russo, C.; Coffin, J.M. Identification, characterization, and comparative genomic distribution of the HERV-K (HML-2) group of human endogenous retroviruses. Retrovirology 2011, 8, 90. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, C.; Simmonds, P. Allelic Variation of HERV-K(HML-2) Endogenous Retroviral Elements in Human Populations. J. Mol. Evol. 2004, 59, 642–656. [Google Scholar] [CrossRef] [PubMed]
- Blomberg, J.; Benachenhou, F.; Blikstad, V.; Sperber, G.; Mayer, J. Classification and nomenclature of endogenous retroviral sequences (ERVs). Gene 2009, 448, 115–123. [Google Scholar] [CrossRef]
- Mayer, J.; Blomberg, J.; Seal, R.L. A revised nomenclature for transcribed human endogenous retroviral loci. Mob. DNA 2011, 2, 7. [Google Scholar] [CrossRef]
- Wang, X.; Huang, J.; Zhu, F. Human Endogenous Retroviral Envelope Protein Syncytin-1 and Inflammatory Abnormalities in Neuropsychological Diseases. Front. Psychiatry 2018, 9, 422. [Google Scholar] [CrossRef]
- van der Kuyl, A.C. HIV infection and HERV expression: A review. Retrovirology 2012. [Google Scholar] [CrossRef]
- Jern, P.; Sperber, G.O.; Blomberg, J. Use of endogenous retroviral sequences (ERVs) and structural markers for retroviral phylogenetic inference and taxonomy. Retrovirology 2005, 2, 50. [Google Scholar] [CrossRef]
- Parrish, N.F.; Tomonaga, K. Endogenized viral sequences in mammals. Curr. Opin. Microbiol. 2016, 31, 176–183. [Google Scholar] [CrossRef]
- Gemmell, P.; Hein, J.; Katzourakis, A. Phylogenetic Analysis Reveals That ERVs “Die Young” but HERV-H Is Unusually Conserved. PLOS Comput. Biol. 2016, 12, e1004964. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.F.; Coffin, J.M. Human endogenous retrovirus K solo-LTR formation and insertional polymorphisms: Implications for human and viral evolution. Proc. Natl. Acad. Sci. USA 2004, 101, 1668–1672. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.; Perron, H.; Feschotte, C. Variation in proviral content among human genomes mediated by LTR recombination. Mob. DNA 2018, 9, 36. [Google Scholar] [CrossRef]
- Cohen, C.J.; Lock, W.M.; Mager, D.L. Endogenous retroviral LTRs as promoters for human genes: A critical assessment. Gene 2009, 448, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Galindo, R.; Kaplan, M.H.; Leissner, P.; Verjat, T.; Ferlenghi, I.; Bagnoli, F.; Giusti, F.; Dosik, M.H.; Hayes, D.F.; Gitlin, S.D.; et al. Human Endogenous Retrovirus K (HML-2) Elements in the Plasma of People with Lymphoma and Breast Cancer. J. Virol. 2008, 82, 9329–9336. [Google Scholar] [CrossRef] [PubMed]
- Lower, R.; Lower, J.; Frank, H.; Harzmann, R.; Kurth, R. Human Teratocarcinomas Cultured in vitro Produce Unique Retrovirus-like Viruses. J. Gen. Virol. 1984, 65, 887–898. [Google Scholar] [CrossRef]
- Büscher, K.; Trefzer, U.; Hofmann, M.; Sterry, W.; Kurth, R.; Denner, J. Expression of Human Endogenous Retrovirus K in Melanomas and Melanoma Cell Lines. Cancer Res. 2005, 65, 4172–4180. [Google Scholar] [CrossRef]
- Schmitt, K.; Reichrath, J.; Roesch, A.; Meese, E.; Mayer, J. Transcriptional Profiling of Human Endogenous Retrovirus Group HERV-K(HML-2) Loci in Melanoma. Genome Biol. Evol. 2013, 5, 307–328. [Google Scholar] [CrossRef]
- Moyes, D.; Griffiths, D.J.; Venables, P.J. Insertional polymorphisms: A new lease of life for endogenous retroviruses in human disease. Trends Genet. 2007, 23, 326–333. [Google Scholar] [CrossRef]
- Löwer, R.; Löwer, J.; Tondera-Koch, C.; Kurth, R. A General Method for the Identification of Transcribed Retrovirus Sequences (R-U5 PCR) Reveals the Expression of the Human Endogenous Retrovirus Loci HERV-H and HERV-K in Teratocarcinoma Cells. Virology 1993, 192, 501–511. [Google Scholar] [CrossRef]
- Chan, S.M.; Sapir, T.; Park, S.-S.; Rual, J.-F.; Contreras-Galindo, R.; Reiner, O.; Markovitz, D.M. The HERV-K accessory protein Np9 controls viability and migration of teratocarcinoma cells. PLoS ONE 2019, 14, e0212970. [Google Scholar] [CrossRef] [PubMed]
- Kassiotis, G. Endogenous Retroviruses and the Development of Cancer. J. Immunol. 2014, 192, 1343–1349. [Google Scholar] [CrossRef] [PubMed]
- Lavie, L.; Kitova, M.; Maldener, E.; Meese, E.; Mayer, J. CpG Methylation Directly Regulates Transcriptional Activity of the Human Endogenous Retrovirus Family HERV-K(HML-2). J. Virol. 2005, 79, 876–883. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, K.; Shinkai, Y. SETDB1-Mediated Silencing of Retroelements. Viruses 2020, 12, 596. [Google Scholar] [CrossRef] [PubMed]
- Cuellar, T.L.; Herzner, A.-M.; Zhang, X.; Goyal, Y.; Watanabe, C.; Friedman, B.A.; Janakiraman, V.; Durinck, S.; Stinson, J.; Arnott, D.; et al. Silencing of retrotransposons by SETDB1 inhibits the interferon response in acute myeloid leukemia. J. Cell Biol. 2017, 216, 3535–3549. [Google Scholar] [CrossRef]
- Zhang, M.; Liang, J.Q. Expressional activation and functional roles of human endogenous retroviruses in cancers. Rev. Med. Virol. 2019, 29, e2025. [Google Scholar] [CrossRef]
- Laska, M.J.; Nissen, K.K.; Nexø, B.A. (Some) Cellular Mechanisms Influencing the Transcription of Human Endogenous Retrovirus, HERV-Fc1. PLoS ONE 2013, 8, e53895. [Google Scholar] [CrossRef]
- Goodier, J.L. Restricting retrotransposons: A review. Mob. DNA 2016, 7, 16. [Google Scholar] [CrossRef]
- Stengel, S.; Fiebig, U.; Kurth, R.; Denner, J. Regulation of human endogenous retrovirus-K expression in melanomas by CpG methylation. Genes Chromosomes Cancer 2010, 49, 401–411. [Google Scholar] [CrossRef]
- Kreimer, U.; Schulz, W.A.; Koch, A.; Niegisch, G.; Goering, W. HERV-K and LINE-1 DNA Methylation and Reexpression in Urothelial Carcinoma. Front. Oncol. 2013, 3, 1–12. [Google Scholar] [CrossRef]
- Hurst, T.; Pace, M.; Katzourakis, A.; Phillips, R.; Klenerman, P.; Frater, J.; Magiorkinis, G. Human endogenous retrovirus (HERV) expression is not induced by treatment with the histone deacetylase (HDAC) inhibitors in cellular models of HIV-1 latency. Retrovirology 2016, 13, 10. [Google Scholar] [CrossRef] [PubMed]
- Iramaneerat, K.; Rattanatunyong, P.; Khemapech, N.; Triratanachat, S.; Mutirangura, A. HERV-K Hypomethylation in Ovarian Clear Cell Carcinoma Is Associated With a Poor Prognosis and Platinum Resistance. Int. J. Gynecol. Cancer 2011, 21, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Montesion, M.; Williams, Z.H.; Subramanian, R.P.; Kuperwasser, C.; Coffin, J.M. Promoter expression of HERV-K (HML-2) provirus-derived sequences is related to LTR sequence variation and polymorphic transcription factor binding sites. Retrovirology 2018, 15, 57. [Google Scholar] [CrossRef] [PubMed]
- Zhdanov, V.M.; Soloviev, V.D.; Bektemirov, T.A.; Ilyin, K.V.; Bykovsky, A.F.; Mazurenko, N.P.; Irlin, I.S.; Yershov, F.I. Isolation of Oncornaviruses from Continuous Human Cell Cultures. Intervirology 1973, 1, 19–26. [Google Scholar] [CrossRef]
- Sarngadharan, M.G.; Sarin, P.S.; Reitz, M.S.; Gallo, R.C. Reverse Transcriptase Activity of Human Acute Leukaemic Cells: Purification of the Enzyme, Response to AMV 70S RNA, and Characterization of the DNA Product. Nat. New Biol. 1972, 240, 67–72. [Google Scholar] [CrossRef]
- Matteucci, C.; Balestrieri, E.; Argaw-Denboba, A.; Sinibaldi-Vallebona, P. Human endogenous retroviruses role in cancer cell stemness. Semin. Cancer Biol. 2018, 53, 17–30. [Google Scholar] [CrossRef]
- Barth, M.; Gröger, V.; Cynis, H.; Staege, M.S. Identification of human endogenous retrovirus transcripts in Hodgkin Lymphoma cells. Mol. Biol. Rep. 2019, 46, 1885–1893. [Google Scholar] [CrossRef]
- Grabski, D.F.; Hu, Y.; Sharma, M.; Rasmussen, S.K. Close to the Bedside: A Systematic Review of Endogenous Retroviruses and Their Impact in Oncology. J. Surg. Res. 2019, 240, 145–155. [Google Scholar] [CrossRef]
- Golan, M.; Hizi, A.; Resau, J.H.; Yaal-Hahoshen, N.; Reichman, H.; Keydar, I.; Tsarfaty, I. Human Endogenous Retrovirus (HERV-K) Reverse Transcriptase as a Breast Cancer Prognostic Marker. Neoplasia 2008, 10, 521-IN2. [Google Scholar] [CrossRef] [PubMed]
- Tavakolian, S.; Goudarzi, H.; Faghihloo, E. Evaluating the expression level of HERV-K env, np9, rec and gag in breast tissue. Infect. Agent. Cancer 2019, 14, 1–5. [Google Scholar] [CrossRef]
- Zhao, J.; Rycaj, K.; Geng, S.; Li, M.; Plummer, J.B.; Yin, B.; Liu, H.; Xu, X.; Zhang, Y.; Yan, Y.; et al. Expression of Human Endogenous Retrovirus Type K Envelope Protein is a Novel Candidate Prognostic Marker for Human Breast Cancer. Genes Cancer 2011, 2, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Johanning, G.L.; Malouf, G.G.; Zheng, X.; Esteva, F.J.; Weinstein, J.N.; Wang-Johanning, F.; Su, X. Expression of human endogenous retrovirus-K is strongly associated with the basal-like breast cancer phenotype. Sci. Rep. 2017, 7, 41960. [Google Scholar] [CrossRef] [PubMed]
- Wang-Johanning, F.; Li, M.; Esteva, F.J.; Hess, K.R.; Yin, B.; Rycaj, K.; Plummer, J.B.; Garza, J.G.; Ambs, S.; Johanning, G.L. Human endogenous retrovirus type K antibodies and mRNA as serum biomarkers of early-stage breast cancer. Int. J. Cancer 2014, 134, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Muster, T.; Waltenberger, A.; Grassauer, A.; Hirschl, S.; Caucig, P.; Romirer, I.; Födinger, D.; Seppele, H.; Schanab, O.; Magin-Lachmann, C.; et al. An Endogenous Retrovirus Derived from Human Melanoma Cells. Cancer Res. 2003, 63, 8735–8741. [Google Scholar]
- Singh, S.; Kaye, S.; Francis, N.; Peston, D.; Gore, M.; McClure, M.; Bunker, C. Human endogenous retrovirus K (HERV-K) rec mRNA is expressed in primary melanoma but not in benign naevi or normal skin. Pigment Cell Melanoma Res. 2013, 26, 426–428. [Google Scholar] [CrossRef]
- María, G.-C.; Paola, I.; Niki, K.; Mariacarmela, S.; Julià, B.; Rafael, R. Human endogenous retroviruses and cancer. Cancer Biol. Med. 2016, 13, 483. [Google Scholar] [CrossRef]
- Wallace, A.D.; Wendt, G.A.; Barcellos, L.F.; de Smith, A.J.; Walsh, K.M.; Metayer, C.; Costello, J.F.; Wiemels, J.L.; Francis, S.S. To ERV Is Human: A Phenotype-Wide Scan Linking Polymorphic Human Endogenous Retrovirus-K Insertions to Complex Phenotypes. Front. Genet. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Kahyo, T.; Tao, H.; Shinmura, K.; Yamada, H.; Mori, H.; Funai, K.; Kurabe, N.; Suzuki, M.; Tanahashi, M.; Niwa, H.; et al. Identification and association study with lung cancer for novel insertion polymorphisms of human endogenous retrovirus. Carcinogenesis 2013, 34, 2531–2538. [Google Scholar] [CrossRef]
- Burmeister, T.; Ebert, A.D.; Pritze, W.; Loddenkemper, C.; Schwartz, S.; Thiel, E. Insertional Polymorphisms of Endogenous HERV-K113 and HERV-K115 Retroviruses in Breast Cancer Patients and Age-Matched Controls. AIDS Res. Hum. Retrovir. 2004, 20, 1223–1229. [Google Scholar] [CrossRef]
- Wildschutte, J.H.; Ram, D.; Subramanian, R.; Stevens, V.L.; Coffin, J.M. The distribution of insertionally polymorphic endogenous retroviruses in breast cancer patients and cancer-free controls. Retrovirology 2014, 11, 62. [Google Scholar] [CrossRef]
- Fuentes, D.R.; Swigut, T.; Wysocka, J. Systematic perturbation of retroviral LTRs reveals widespread long-range effects on human gene regulation. Elife 2018, 7, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.S.; Shah, N.M.; Du, A.Y.; Dailey, Z.Z.; Pehrsson, E.C.; Godoy, P.M.; Zhang, D.; Li, D.; Xing, X.; Kim, S.; et al. Transposable elements drive widespread expression of oncogenes in human cancers. Nat. Genet. 2019, 51, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Babaian, A.; Mager, D.L. Endogenous retroviral promoter exaptation in human cancer. Mob. DNA 2016, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Lamprecht, B.; Walter, K.; Kreher, S.; Kumar, R.; Hummel, M.; Lenze, D.; Köchert, K.; Bouhlel, M.A.; Richter, J.; Soler, E.; et al. Derepression of an endogenous long terminal repeat activates the CSF1R proto-oncogene in human lymphoma. Nat. Med. 2010, 16, 571–579. [Google Scholar] [CrossRef]
- Grandi, N.; Tramontano, E. HERV Envelope Proteins: Physiological Role and Pathogenic Potential in Cancer and Autoimmunity. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Berndt, B.; Haverkampf, S.; Reith, G.; Keil, S.; Niggemann, B.; Zänker, K.S.; Dittmar, T. Fusion of CCL21 Non-Migratory Active Breast Epithelial and Breast Cancer Cells Give Rise to CCL21 Migratory Active Tumor Hybrid Cell Lines. PLoS ONE 2013, 8, e63711. [Google Scholar] [CrossRef]
- Bastida-Ruiz, D.; Van Hoesen, K.; Cohen, M. The Dark Side of Cell Fusion. Int. J. Mol. Sci. 2016, 17, 638. [Google Scholar] [CrossRef]
- Kassiotis, G.; Stoye, J.P. Making a virtue of necessity: The pleiotropic role of human endogenous retroviruses in cancer. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160277. [Google Scholar] [CrossRef]
- Ruprecht, K.; Mayer, J.; Sauter, M.; Roemer, K.; Mueller-Lantzsch, N. Endogenous retroviruses. Cell. Mol. Life Sci. 2008, 65, 3366–3382. [Google Scholar] [CrossRef] [PubMed]
- Alcazer, V.; Bonaventura, P.; Depil, S. Human Endogenous Retroviruses (HERVs): Shaping the Innate Immune Response in Cancers. Cancers 2020, 12, 610. [Google Scholar] [CrossRef]
- Krone, B.; Kölmel, K.F.; Henz, B.M.; Grange, J.M. Protection against melanoma by vaccination with Bacille Calmette-Guérin (BCG) and/or vaccinia: An epidemiology-based hypothesis on the nature of a melanoma risk factor and its immunological control. Eur. J. Cancer 2005, 41, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Radvanyi, L.; Yin, B.; Rycaj, K.; Li, J.; Chivukula, R.; Lin, K.; Lu, Y.; Shen, J.; Chang, D.Z.; et al. Downregulation of Human Endogenous Retrovirus Type K (HERV-K) Viral env RNA in Pancreatic Cancer Cells Decreases Cell Proliferation and Tumor Growth. Clin. Cancer Res. 2017, 23, 5892–5911. [Google Scholar] [CrossRef] [PubMed]
- Ibba, G.; Piu, C.; Uleri, E.; Serra, C.; Dolei, A. Disruption by SaCas9 Endonuclease of HERV-Kenv, a Retroviral Gene with Oncogenic and Neuropathogenic Potential, Inhibits Molecules Involved in Cancer and Amyotrophic Lateral Sclerosis. Viruses 2018, 10, 412. [Google Scholar] [CrossRef]
- Magin-Lachmann, C.; Hahn, S.; Strobel, H.; Held, U.; Löwer, J.; Löwer, R. Rec (Formerly Corf) Function Requires Interaction with a Complex, Folded RNA Structure within Its Responsive Element rather than Binding to a Discrete Specific Binding Site. J. Virol. 2001, 75, 10359–10371. [Google Scholar] [CrossRef]
- Johnson, W.E. Origins and evolutionary consequences of ancient endogenous retroviruses. Nat. Rev. Microbiol. 2019, 17, 355–370. [Google Scholar] [CrossRef] [PubMed]
- Lower, R.; Boller, K.; Hasenmaier, B.; Korbmacher, C.; Muller-Lantzsch, N.; Lower, J.; Kurth, R. Identification of human endogenous retroviruses with complex mRNA expression and particle formation. Proc. Natl. Acad. Sci. USA 1993, 90, 4480–4484. [Google Scholar] [CrossRef]
- Boese, A.; Sauter, M.; Galli, U.; Best, B.; Herbst, H.; Mayer, J.; Kremmer, E.; Roemer, K.; Mueller-Lantzsch, N. Human endogenous retrovirus protein cORF supports cell transformation and associates with the promyelocytic leukemia zinc finger protein. Oncogene 2000, 19, 4328–4336. [Google Scholar] [CrossRef]
- Galli, U.M.; Sauter, M.; Lecher, B.; Maurer, S.; Herbst, H.; Roemer, K.; Mueller-Lantzsch, N. Human endogenous retrovirus rec interferes with germ cell development in mice and may cause carcinoma in situ, the predecessor lesion of germ cell tumors. Oncogene 2005, 24, 3223–3228. [Google Scholar] [CrossRef]
- Denne, M.; Sauter, M.; Armbruester, V.; Licht, J.D.; Roemer, K.; Mueller-Lantzsch, N. Physical and Functional Interactions of Human Endogenous Retrovirus Proteins Np9 and Rec with the Promyelocytic Leukemia Zinc Finger Protein. J. Virol. 2007, 81, 5607–5616. [Google Scholar] [CrossRef] [PubMed]
- Hanke, K.; Chudak, C.; Kurth, R.; Bannert, N. The Rec protein of HERV-K(HML-2) upregulates androgen receptor activity by binding to the human small glutamine-rich tetratricopeptide repeat protein (hSGT). Int. J. Cancer 2013, 132, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, S.; Sauter, M.; Schmitt, M.; Baumert, B.; Best, B.; Boese, A.; Roemer, K.; Mueller-Lantzsch, N. Human endogenous retrovirus protein Rec interacts with the testicular zinc-finger protein and androgen receptor. J. Gen. Virol. 2010, 91, 1494–1502. [Google Scholar] [CrossRef] [PubMed]
- Hanke, K.; Hohn, O.; Bannert, N. HERV-K(HML-2), a seemingly silent subtenant—But still waters run deep. APMIS 2016, 124, 67–87. [Google Scholar] [CrossRef] [PubMed]
- Armbruester, V.; Sauter, M.; Krautkraemer, E.; Meese, E.; Kleiman, A.; Best, B.; Roemer, K.; Mueller-Lantzsch, N. A novel gene from the human endogenous retrovirus K expressed in transformed cells. Clin. Cancer Res. 2002, 8, 1800–1807. [Google Scholar]
- Armbruester, V.; Sauter, M.; Roemer, K.; Best, B.; Hahn, S.; Nty, A.; Schmid, A.; Philipp, S.; Mueller, A.; Mueller-Lantzsch, N. Np9 Protein of Human Endogenous Retrovirus K Interacts with Ligand of Numb Protein X. J. Virol. 2004, 78, 10310–10319. [Google Scholar] [CrossRef][Green Version]
- Flores, A.N.; McDermott, N.; Meunier, A.; Marignol, L. NUMB inhibition of NOTCH signalling as a therapeutic target in prostate cancer. Nat. Rev. Urol. 2014, 11, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Heyne, K.; Kölsch, K.; Bruand, M.; Kremmer, E.; Grässer, F.A.; Mayer, J.; Roemer, K. Np9, a cellular protein of retroviral ancestry restricted to human, chimpanzee and gorilla, binds and regulates ubiquitin ligase MDM2. Cell Cycle 2015, 14, 2619–2633. [Google Scholar] [CrossRef]
- Chen, T.; Meng, Z.; Gan, Y.; Wang, X.; Xu, F.; Gu, Y.; Xu, X.; Tang, J.; Zhou, H.; Zhang, X.; et al. The viral oncogene Np9 acts as a critical molecular switch for co-activating β-catenin, ERK, Akt and Notch1 and promoting the growth of human leukemia stem/progenitor cells. Leukemia 2013, 27, 1469–1478. [Google Scholar] [CrossRef]
- Smith, C.C.; Selitsky, S.R.; Chai, S.; Armistead, P.M.; Vincent, B.G.; Serody, J.S. Alternative tumour-specific antigens. Nat. Rev. Cancer 2019, 19, 465–478. [Google Scholar] [CrossRef]
- Chiappinelli, K.B.; Strissel, P.L.; Desrichard, A.; Li, H.; Henke, C.; Akman, B.; Hein, A.; Rote, N.S.; Cope, L.M.; Snyder, A.; et al. Inhibiting DNA Methylation Causes an Interferon Response in Cancer via dsRNA Including Endogenous Retroviruses. Cell 2015, 162, 974–986. [Google Scholar] [CrossRef]
- Goel, S.; DeCristo, M.J.; Watt, A.C.; BrinJones, H.; Sceneay, J.; Li, B.B.; Khan, N.; Ubellacker, J.M.; Xie, S.; Metzger-Filho, O.; et al. CDK4/6 inhibition triggers anti-tumour immunity. Nature 2017, 548, 471–475. [Google Scholar] [CrossRef]
- Smith, C.C.; Beckermann, K.E.; Bortone, D.S.; De Cubas, A.A.; Bixby, L.M.; Lee, S.J.; Panda, A.; Ganesan, S.; Bhanot, G.; Wallen, E.M.; et al. Endogenous retroviral signatures predict immunotherapy response in clear cell renal cell carcinoma. J. Clin. Investig. 2018, 128, 4804–4820. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Harashima, N.; Kajigaya, S.; Yokoyama, H.; Cherkasova, E.; McCoy, J.P.; Hanada, K.; Mena, O.; Kurlander, R.; Abdul, T.; et al. Regression of human kidney cancer following allogeneic stem cell transplantation is associated with recognition of an HERV-E antigen by T cells. J. Clin. Investig. 2008. [Google Scholar] [CrossRef]
- Cherkasova, E.; Scrivani, C.; Doh, S.; Weisman, Q.; Takahashi, Y.; Harashima, N.; Yokoyama, H.; Srinivasan, R.; Linehan, W.M.; Lerman, M.I.; et al. Detection of an Immunogenic HERV-E Envelope with Selective Expression in Clear Cell Kidney Cancer. Cancer Res. 2016, 76, 2177–2185. [Google Scholar] [CrossRef] [PubMed]
- Wang-Johanning, F.; Rycaj, K.; Plummer, J.B.; Li, M.; Yin, B.; Frerich, K.; Garza, J.G.; Shen, J.; Lin, K.; Yan, P.; et al. Immunotherapeutic potential of anti-human endogenous retrovirus-K envelope protein antibodies in targeting breast tumors. J. Natl. Cancer Inst. 2012, 104, 189–210. [Google Scholar] [CrossRef]
- Kraus, B.; Fischer, K.; Büchner, S.M.; Wels, W.S.; Löwer, R.; Sliva, K.; Schnierle, B.S. Vaccination Directed against the Human Endogenous Retrovirus-K Envelope Protein Inhibits Tumor Growth in a Murine Model System. PLoS ONE 2013, 8, e72756. [Google Scholar] [CrossRef]
- Rycaj, K.; Plummer, J.B.; Yin, B.; Li, M.; Garza, J.; Radvanyi, L.; Ramondetta, L.M.; Lin, K.; Johanning, G.L.; Tang, D.G.; et al. Cytotoxicity of Human Endogenous Retrovirus K-Specific T Cells toward Autologous Ovarian Cancer Cells. Clin. Cancer Res. 2015, 21, 471–483. [Google Scholar] [CrossRef]
- Zhou, F.; Krishnamurthy, J.; Wei, Y.; Li, M.; Hunt, K.; Johanning, G.L.; Cooper, L.J.N.; Wang-Johanning, F. Chimeric antigen receptor T cells targeting HERV-K inhibit breast cancer and its metastasis through downregulation of Ras. Oncoimmunology 2015, 4, e1047582. [Google Scholar] [CrossRef]
- Kraus, B.; Fischer, K.; Sliva, K.; Schnierle, B.S. Vaccination directed against the human endogenous retrovirus-K (HERV-K) gag protein slows HERV-K gag expressing cell growth in a murine model system. Virol. J. 2014, 11, 58. [Google Scholar] [CrossRef]
- Mastrangelo, G.; Krone, B.; Fadda, E.; Buja, A.; Grange, J.M.; Rausa, G.; de Vries, E.; Koelmel, K.F. Does yellow fever 17D vaccine protect against melanoma? Vaccine 2009, 27, 588–591. [Google Scholar] [CrossRef]
- Tran, T.; Burt, D.; Eapen, L.; Keller, O.R. Spontaneous regression of metastatic melanoma after inoculation with tetanus-diphtheria-pertussis vaccine. Curr. Oncol. 2013, 20, 270. [Google Scholar] [CrossRef]
- Schiavetti, F.; Thonnard, J.; Colau, D.; Boon, T.; Coulie, P.G. A human endogenous retroviral sequence encoding an antigen recognized on melanoma by cytolytic T lymphocytes. Cancer Res. 2002, 62, 5510–5516. [Google Scholar]
- Büscher, K.; Hahn, S.; Hofmann, M.; Trefzer, U.; Özel, M.; Sterry, W.; Löwer, J.; Löwer, R.; Kurth, R.; Denner, J. Expression of the human endogenous retrovirus-K transmembrane envelope, Rec and Np9 proteins in melanomas and melanoma cell lines. Melanoma Res. 2006, 16, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Humer, J.; Waltenberger, A.; Grassauer, A.; Kurz, M.; Valencak, J.; Rapberger, R.; Hahn, S.; Löwer, R.; Wolff, K.; Bergmann, M.; et al. Identification of a Melanoma Marker Derived from Melanoma-Associated Endogenous Retroviruses. Cancer Res. 2006, 66, 1658–1663. [Google Scholar] [CrossRef] [PubMed]
- Hahn, S.; Ugurel, S.; Hanschmann, K.-M.; Strobel, H.; Tondera, C.; Schadendorf, D.; Löwer, J.; Löwer, R. Serological Response to Human Endogenous Retrovirus K in Melanoma Patients Correlates with Survival Probability. AIDS Res. Hum. Retrovir. 2008, 24, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, J.; Rabinovich, B.A.; Mi, T.; Switzer, K.C.; Olivares, S.; Maiti, S.N.; Plummer, J.B.; Singh, H.; Kumaresan, P.R.; Huls, H.M.; et al. Genetic Engineering of T Cells to Target HERV-K, an Ancient Retrovirus on Melanoma. Clin. Cancer Res. 2015, 21, 3241–3251. [Google Scholar] [CrossRef] [PubMed]
- Reis, B.S.; Jungbluth, A.A.; Frosina, D.; Holz, M.; Ritter, E.; Nakayama, E.; Ishida, T.; Obata, Y.; Carver, B.; Scher, H.; et al. Prostate Cancer Progression Correlates with Increased Humoral Immune Response to a Human Endogenous Retrovirus GAG Protein. Clin. Cancer Res. 2013, 19, 6112–6125. [Google Scholar] [CrossRef]
- Wallace, T.A.; Downey, R.F.; Seufert, C.J.; Schetter, A.; Dorsey, T.H.; Johnson, C.A.; Goldman, R.; Loffredo, C.A.; Yan, P.; Sullivan, F.J.; et al. Elevated HERV-K mRNA expression in PBMC is associated with a prostate cancer diagnosis particularly in older men and smokers. Carcinogenesis 2014, 35, 2074–2083. [Google Scholar] [CrossRef]
- Rastogi, A.; Ali, A.; Tan, S.-H.; Banerjee, S.; Chen, Y.; Cullen, J.; Xavier, C.P.; Mohamed, A.A.; Ravindranath, L.; Srivastav, J.; et al. Autoantibodies against oncogenic ERG protein in prostate cancer: Potential use in diagnosis and prognosis in a panel with C-MYC, AMACR and HERV-K Gag. Genes Cancer 2016, 7, 394–413. [Google Scholar] [CrossRef][Green Version]
- Kleiman, A.; Senyuta, N.; Tryakin, A.; Sauter, M.; Karseladze, A.; Tjulandin, S.; Gurtsevitch, V.; Mueller-Lantzsch, N. HERV-K(HML-2) GAG/ENV antibodies as indicator for therapy effect in patients with germ cell tumors. Int. J. Cancer 2004, 110, 459–461. [Google Scholar] [CrossRef]
- Schmitz-Winnenthal, F.H.; Galindo-Escobedo, L.V.; Rimoldi, D.; Geng, W.; Romero, P.; Koch, M.; Weitz, J.; Krempien, R.; Niethammer, A.G.; Beckhove, P.; et al. Potential target antigens for immunotherapy in human pancreatic cancer. Cancer Lett. 2007, 252, 290–298. [Google Scholar] [CrossRef]
- Ma, W.; Hong, Z.; Liu, H.; Chen, X.; Ding, L.; Liu, Z.; Zhou, F.; Yuan, Y. Human Endogenous Retroviruses-K (HML-2) Expression Is Correlated with Prognosis and Progress of Hepatocellular Carcinoma. BioMed Res. Int. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pérot, P.; Cheynet, V.; Decaussin-Petrucci, M.; Oriol, G.; Mugnier, N.; Rodriguez-Lafrasse, C.; Ruffion, A.; Mallet, F. Microarray-based Identification of Individual HERV Loci Expression: Application to Biomarker Discovery in Prostate Cancer. J. Vis. Exp. 2013. [Google Scholar] [CrossRef] [PubMed]
- Schulz, W.A. Does HERV-K represent a potential therapeutic target for prostate cancer? Expert Opin. Ther. Targets 2017, 21, 921–924. [Google Scholar] [CrossRef] [PubMed]
- Ishida, T.; Obata, Y.; Ohara, N.; Matsushita, H.; Sato, S.; Uenaka, A.; Saika, T.; Miyamura, T.; Chayama, K.; Nakamura, Y.; et al. Identification of the HERV-K gag antigen in prostate cancer by SEREX using autologous patient serum and its immunogenicity. Cancer Immun. 2008, 8, 15. [Google Scholar]
- Wang-Johanning, F.; Frost, A.R.; Jian, B.; Azerou, R.; Lu, D.W.; Chen, D.-T.; Johanning, G.L. Detecting the expression of human endogenous retrovirus Eenvelope transcripts in human prostate adenocarcinoma. Cancer 2003, 98, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Rakoff-Nahoum, S.J.; Kuebler, P.J.; Heymann, J.E.; Sheehy, M.M.; Ortiz, G.S.; Ogg, G.; Barbour, J.D.; Lenz, J.; Steinfeld, A.D.; Nixon, D.F. Detection of T Lymphocytes Specific for Human Endogenous Retrovirus K (HERV-K) in Patients with Seminoma. AIDS Res. Hum. Retrovir. 2006, 22, 52–56. [Google Scholar] [CrossRef]
- Casau, A.E.; Vaughan, J.E.; Lozano, G.; Levine, A.J. Germ Cell Expression of an Isolated Human Endogenous Retroviral Long Terminal Repeat of the HERV-K/HTDV Family in Transgenic Mice. J. Virol. 1999, 73, 9976–9983. [Google Scholar] [CrossRef]
- Mueller, T.; Hantsch, C.; Volkmer, I.; Staege, M.S. Differentiation-Dependent Regulation of Human Endogenous Retrovirus K Sequences and Neighboring Genes in Germ Cell Tumor Cells. Front. Microbiol. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sauter, M.; Schommer, S.; Kremmer, E.; Remberger, K.; Dölken, G.; Lemm, I.; Buck, M.; Best, B.; Neumann-Haefelin, D.; Mueller-Lantzsch, N. Human endogenous retrovirus K10: Expression of Gag protein and detection of antibodies in patients with seminomas. J. Virol. 1995, 69, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Wang-Johanning, F.; Liu, J.; Rycaj, K.; Huang, M.; Tsai, K.; Rosen, D.G.; Chen, D.-T.; Lu, D.W.; Barnhart, K.F.; Johanning, G.L. Expression of multiple human endogenous retrovirus surface envelope proteins in ovarian cancer. Int. J. Cancer 2007, 120, 81–90. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Class | Family | Genus |
|---|---|---|
| Class I | HERV-H, HERV-F, HERV-W, HERV-R, HERV-P, HERV-E, HERV-I, HERV-T, ERV-FTD, ERV-FRD | Gammaretrovirus |
| Class II | HERV-K (HML 1–10) | Betaretrovirus |
| Class III | HERV-L | Distantly related to Lentivirus and Spumavirus |
| Cancer | Study (Year) | Approach | Main Findings | Reference |
|---|---|---|---|---|
| Breast cancer | Golan, M. (2008) | The HERV-K RT expression was examined in 110 paraffin sections from breast carcinoma patients. | HERV-K RT expression correlated with poor prognosis in disease-free patients that go on to develop disease, suggesting HERV-K could be an early prognostic biomarker for breast cancer | [60] |
| Wang-Johanning, F. (2012) | Human breast tissues and peripheral blood mononuclear cells from breast cancer patients and health women were used to analyze anti-HERV Env antibody and T-cell immune responses. | Breast cancer patients show HERV-specific antibody and T-cell immune responses, as well as proinflammatory cytokine production. The HERV-K-specific CD8 T-cell immune response was able to lyse breast cancer cells expressing HERV-K Env. | [16] | |
| Wang-Johanning, F. (2012) | The antitumor effect from anti-HERV-K Env monoclonal antibody was analyzed in vitro by quantifying cellular growth and apoptosis in breast cancer cells. In vivo, the tumor growth was analyzed using a mouse xenograft breast cancer model. | Anti HERV-K Env antibody shows antitumor effect. The antibody was able to inhibit cellular growth and induce apoptosis from breast cancer cells in vitro and in vivo. | [105] | |
| Wang-Johanning, F. (2013) | HERV-K mRNA and anti-HERV-K Env antibody were analyzed in serum samples collected from healthy women and breast cancer women patients. ELISA assay and real-time PCR were used to detect the antibody titer and the levels of HERV-K mRNA, respectively. | Anti-HERV-K Env antibody shows a diagnostic value compared to mammograms. Besides, HERV-K gag mRNA and Gag antibody showed sensitivity and specificity to be used as screening test to early-stage breast cancer diagnosis. | [64] | |
| Zhou, F. (2015) | The chimeric antigen receptor (CAR) specific for HERV-K Env was generated using anti-HERV-K Env antibody. Its antitumor effect was evaluated in vitro and in vivo, using breast cancer cell lines and xenograft breast cancer models, respectively. | HERV-K CAR T-cells showed a tumor-specific cytotoxicity in breast cancer cell lines and in a xenograft mouse breast cancer model. HERV-K CAR T-cells were also able to prevent tumor metastasis. | [108] | |
| Johanning, G.L. (2017) | A total of 512 breast cancer samples (117 basal, 53 Her2-enriched, 212 Luminal A and 130 Luminal B) deposited in the Cancer Genome Atlas were used to analyze four HERV-K loci expressions (HERV-K108 (7p22.1), HERV-K109 (6q14.1), HERV-K113 (19p12b) and HERV-K115 (8p23.1)) in breast cancer patients. | Four HERV loci were upregulated in the basal subtype (poor prognosis breast cancer subtype). HERV-K Env expression was significantly overexpressed in basal tumors in comparison with other upregulated HERV-K genes. | [63] | |
| Melanoma | Schiavetti, F. (2002) | Peripheral blood mononuclear cells from melanoma patients treated with MAGE peptides and that showed tumor regression were isolated for identification of the antigen recognized by their CD8 T-cells. | Melanoma patients vaccinated with MAGE peptides are able to develop cytotoxic CD8 T-cells against HERV-K and to lyse melanoma cells in vitro. | [112] |
| Büscher, K. (2006) | Melanoma biopsies and serum samples from melanoma patients were collected to analyze the anti HERV-K antibody and env, rec and np9 HERV-K expression. | Expression of both env and rec were detected in 39% of the melanoma samples and in 40% of the cell lines. The np9 was detected in 29% melanoma samples and in 21% of the cell lines. Anti-HERV-specific Env antibodies were also detected in melanoma patients, however anti HERV-K Np9 and Rec antibodies were not identified. Immunosuppressive Env protein activity and release of virus particles were reported in vitro. | [113] | |
| Humer, J. (2006) | Serum samples from healthy and melanoma patients from stage I to stage IV were used to analyze anti HERV-K antibodies in melanoma patients. | Serum samples from melanoma patients show statistically significant differences in seroprevalence of anti-HERV-K Env antibody when compared to healthy subjects. | [114] | |
| Hahn, S. (2008) | Serum samples from healthy and melanoma patients were used to analyze anti HERV-K Gag and Env antibodies | Melanoma patients showed anti-HERV-K Gag and Env antibodies levels in the sera. Besides, patients with Anti HERV antibody show a significantly decreased disease-specific overall survival (stage I–IV). | [115] | |
| Krishnamurthy, J. (2015) | Chimeric antigen receptor (CAR) specific to HERV-K Env (K-CAR) were analyzed to kill melanoma cells in vivo using mouse xenograft melanoma model. | HERV-K Env CAR T-cell showed significant antitumor effect in melanoma in vivo, reducing primary tumor and metastatic burden in the mouse xenograft model | [116] | |
| Prostate cancer | Reis, B.S. (2013) | HERV-K gag expression was analyzed in vitro using tissues (normal and tumor) and cell line. Anti HERV-K Gag antibody was also analyzed using serum samples from prostate cancer patients and healthy subjects. | HERV-K gag expression was upregulated in prostate cancer tissues and its expression was regulated both by demethylation and by androgen stimulation. Anti-HERV-K Gag antibody was also most frequent in serum from patients with advanced prostate cancer (stage III-IV) when compared to early prostate cancer (stages I-II), and it was correlated with worse survival. | [117] |
| Wallace, T. A. (2014) | A total of 429 blood samples from African–American and European–American healthy men (n = 135) and those with prostate cancer (n = 294) were used to evaluate HERV-K gag mRNA and Env protein expression by quantitative real-time PCR and immunohistochemistry, respectively. | HERV-K Env protein was upregulated in prostate patients; however African–American patients showed higher expression than European–American patients. High HERV-K gag expression showed 12.87 fold increased odds (95% confidence interval 6.3–26.25) of being diagnosed with prostate cancer in comparison to patients that showed lower expression. HERV-K gag expressions were also associated with older age and smoking status, factors associated with risk of more aggressive prostate cancer disease. | [118] | |
| Rastogi, A. (2016) | Serum samples from 93 prostate cancer patients and 37 healthy subjects were used to analyze the autoantibody detection panel containing ERG, AMACR, C-MYC and HERV-K Gag proteins. | ERG, AMACR, and HERV-K Gag autoantibody detection were able to differentiate prostate cancer patients from healthy subjects. | [119] | |
| Germ cell tumors | Kleiman, A. (2004) | Serum samples from germ cell tumor patients and control donors were collected. The anti-HERV-K Gag and anti-HERV-K Env were detected and clinical analyses were performed | Anti-HERV-K antibodies were detected in 67% of patients. Serological response was associated with clinical manifestation and cancer therapy success. The antibodies may have an important positive prognostic value to chemotherapy. | [120] |
| Ovarian Cancer | Rycaj, K. (2014) | HERV-K expression was analyzed in blood, cancer and normal tissue samples from patients with ovarian cancer and benign diseases. The anti-HERV-K antibodies were investigated in blood samples. PBMC was isolated and in vitro HERV-K Env antigen stimulation was performed. | HERV-K expression was higher in ovarian cancer in comparison to normal and adjacent normal tissues. Moreover, RT protein activity and anti-HERV-K antibodies were detected in blood from ovarian cancer patients. The immune HERV-K-specific T-cells, generated through autologous dendritic cell stimulation by HERV-K Env antigens, showed T-cell proliferation and cytotoxic T-lymphocyte activity against ovarian cancer cells. | [107] |
| Pancreatic cancer | Li, M. (2017) | Pancreatic cancer cell lines, biopsy tissue and patient sera were used for HERV-K expression analyses, virus-like particle detection and knockdown of HERV-K env to analyze the role of HERV-K expression in pancreatic cancer. In addition, an in vivo model was used to analyze the effect of HERV-K knockdown. | HERV-K expression and RT activity were shown in pancreatic cells, cancer tissue and patient sera. Virus-like particles were observed in cell culture supernatants. Moreover, knockdown of HERV-K env expression downregulated the RAS-ERK-RSK signaling pathway, important for cancer progression. The findings suggested that HERV-K proteins can be used as biomarkers and as a target to cancer immunotherapy. | [83] |
| Schmitz-Winnenthal, F.H. (2007) | A total of 130 pancreatic adenocarcinoma tumors and 23 control tissue samples were collected from patients with chronic pancreatitis and from cadaveric donors. Tumor-associated antigen expression of 10 genes, including HERV-K, was assessed by PCR. | HERV-K expression showed a relatively high prevalence, with positivity in 23% of cases, which may be a tumor-associated antigen candidate for specific cancer immunotherapy. | [121] | |
| Hepatocellular Carcinoma (HCC) | Ma, W. (2016) | A total of 84 HCC and normal adjacent tissue samples were collected to detect HERV-K expression by quantitative real-time PCR and clinical correlation analysis was performed. | HEVR-K levels were significantly increased in HCC and were associated with cirrhosis, tumor differentiation and TNM staging. Higher HERV-K expression was reported with poorer cancer prognosis. In addition, HERV-K expression demonstrated diagnostic accuracy (74.7% sensitivity and 67.8% specificity), which may be used as a prognostic biomarker for HCC. | [122] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curty, G.; Marston, J.L.; de Mulder Rougvie, M.; Leal, F.E.; Nixon, D.F.; Soares, M.A. Human Endogenous Retrovirus K in Cancer: A Potential Biomarker and Immunotherapeutic Target. Viruses 2020, 12, 726. https://doi.org/10.3390/v12070726
Curty G, Marston JL, de Mulder Rougvie M, Leal FE, Nixon DF, Soares MA. Human Endogenous Retrovirus K in Cancer: A Potential Biomarker and Immunotherapeutic Target. Viruses. 2020; 12(7):726. https://doi.org/10.3390/v12070726
Chicago/Turabian StyleCurty, Gislaine, Jez L. Marston, Miguel de Mulder Rougvie, Fabio E. Leal, Douglas F. Nixon, and Marcelo A. Soares. 2020. "Human Endogenous Retrovirus K in Cancer: A Potential Biomarker and Immunotherapeutic Target" Viruses 12, no. 7: 726. https://doi.org/10.3390/v12070726
APA StyleCurty, G., Marston, J. L., de Mulder Rougvie, M., Leal, F. E., Nixon, D. F., & Soares, M. A. (2020). Human Endogenous Retrovirus K in Cancer: A Potential Biomarker and Immunotherapeutic Target. Viruses, 12(7), 726. https://doi.org/10.3390/v12070726