Forced Zika Virus Infection of Culex pipiens Leads to Limited Virus Accumulation in Mosquito Saliva

 , , ,
, , , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Mosquito Rearing
2.2. Cells and Viruses
2.3. Infectious Blood Meal
2.4. Intrathoracic Injection
2.5. Salivation Assay
2.6. Infectivity Assay
2.7. RNA Extraction and Reverse Transcriptase PCR
2.8. Mosquito Wing Length Measurement
2.9. Statistical Analysis
3. Results
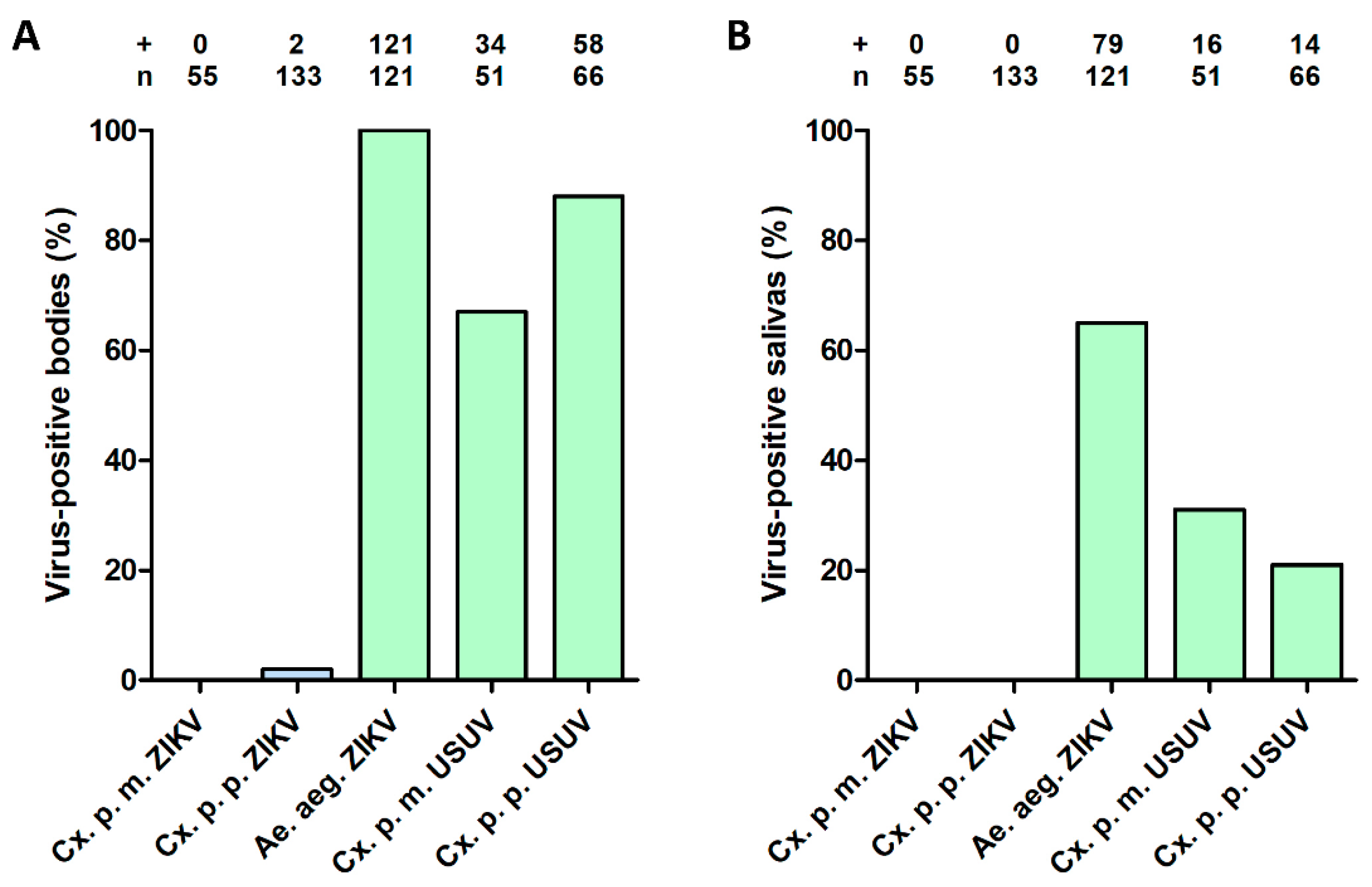
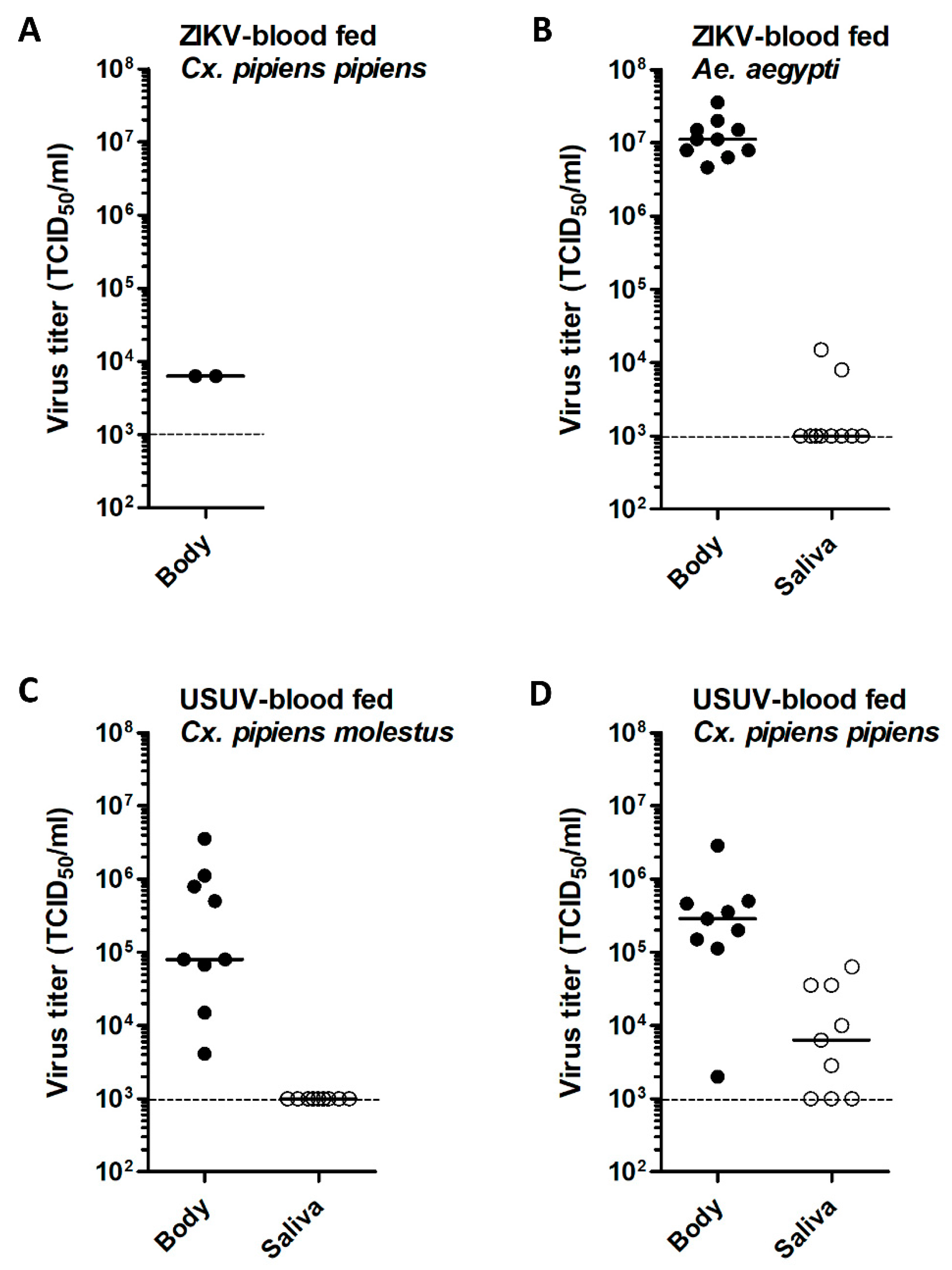
3.1. No ZIKV Transmission by Cx. pipiens after an Infectious Blood Meal
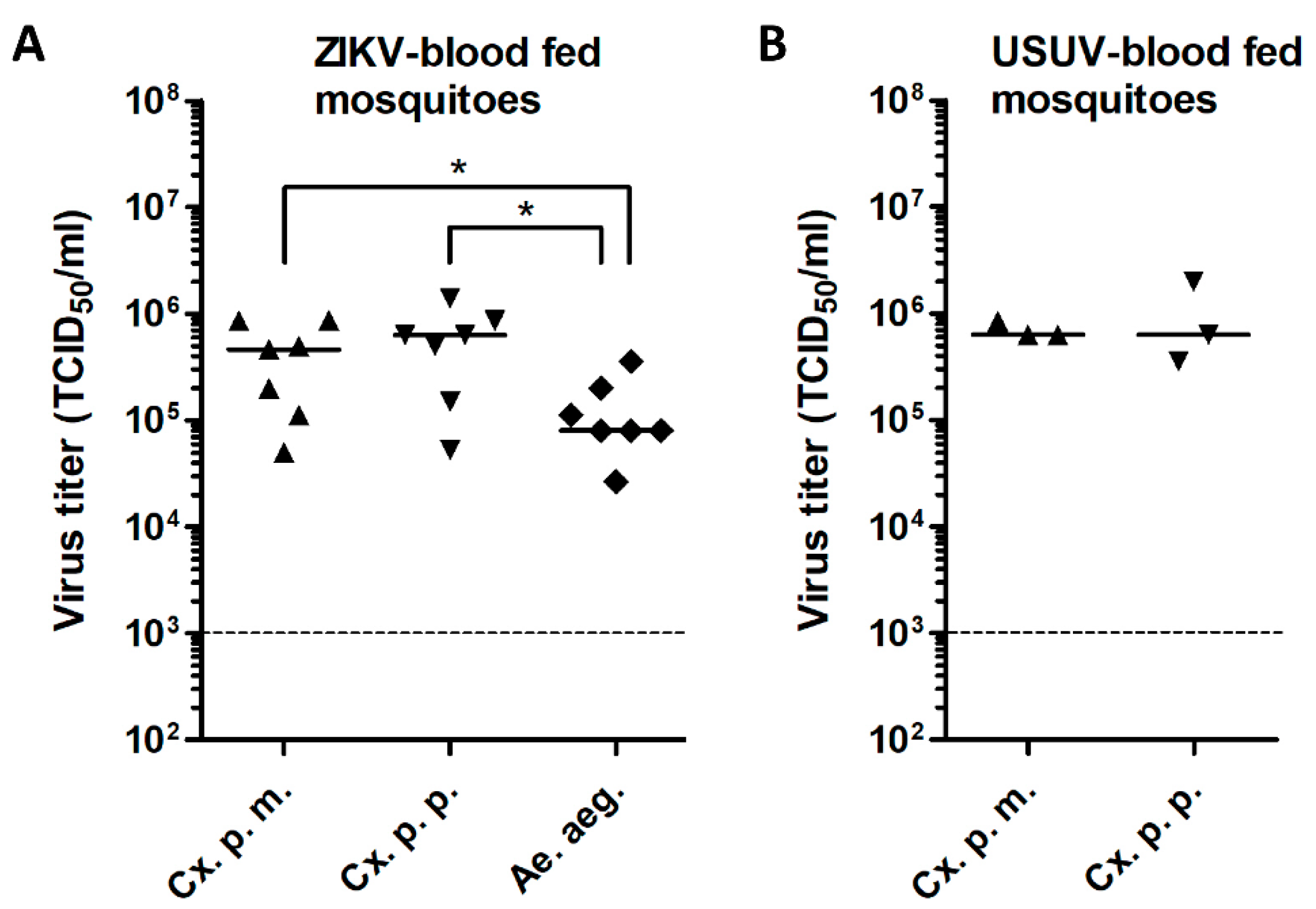
3.2. Low ZIKV Titers in Cx. pipiens after an Infectious Blood Meal
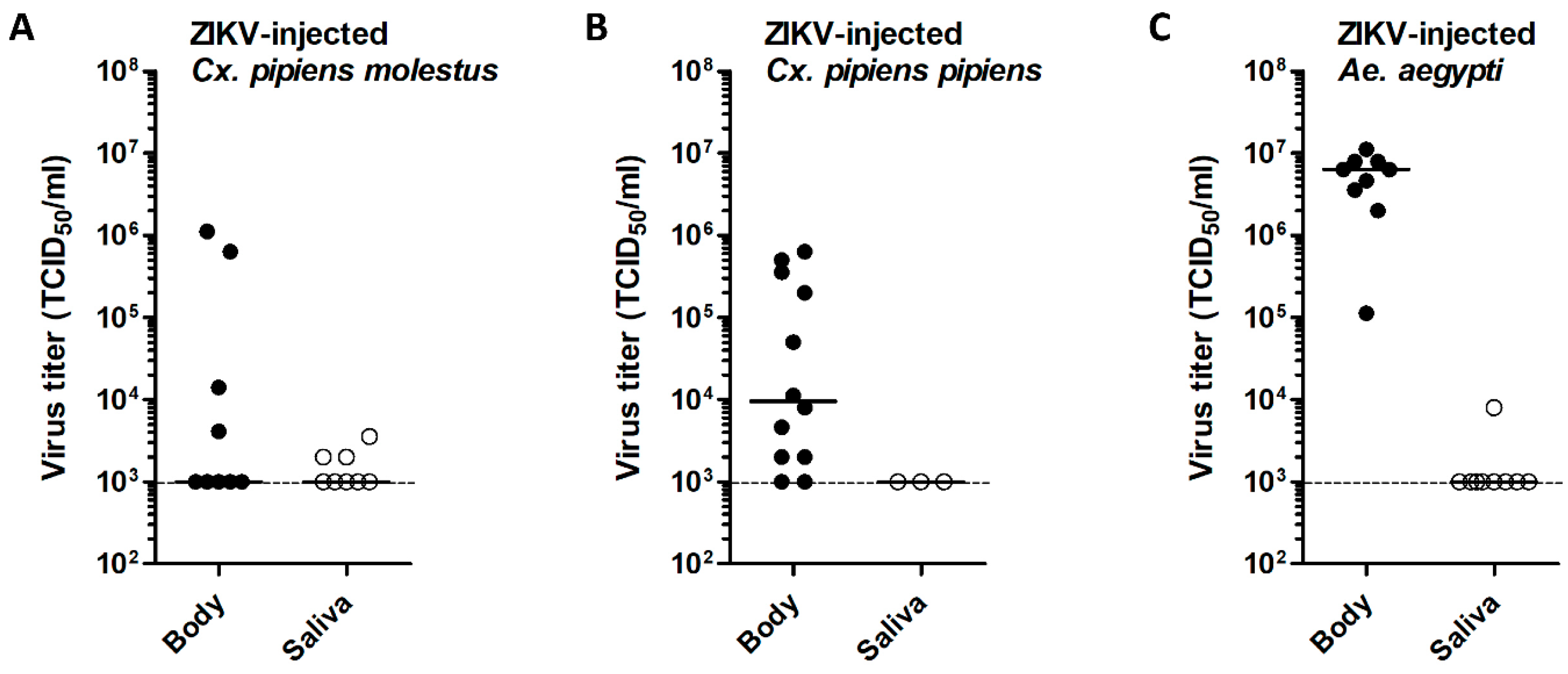
3.3. Intrathoracic ZIKV Injection Leads to Virus Replication in Cx. pipiens with Limited Dissemination to the Mosquito Saliva
3.4. Effect of Injected Viral Dose on ZIKV Infection of Cx. pipiens
3.5. Variability of Viral Titers in ZIKV-Injected Cx. pipiens
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weaver, S.C.; Charlier, C.; Vasilakis, N.; Lecuit, M. Zika, Chikungunya, and Other Emerging Vector-Borne Viral Diseases. Annu. Rev. Med. 2018, 69, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Musso, D.; Gubler, D.J. Zika Virus. Clin. Microbiol. Rev. 2016, 29, 487–524. [Google Scholar] [CrossRef] [PubMed]
- Dick, G.W.; Kitchen, S.F.; Haddow, A.J. Zika virus. I. Isolations and serological specificity. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- MacNamara, F.N. Zika virus: A report on three cases of human infection during an epidemic of jaundice in Nigeria. Trans. R. Soc. Trop. Med. Hyg. 1954, 48, 139–145. [Google Scholar] [CrossRef]
- Zanluca, C.; Melo, V.C.; Mosimann, A.L.; Santos, G.I.; Santos, C.N.; Luz, K. First report of autochthonous transmission of Zika virus in Brazil. Mem. Inst. Oswaldo. Cruz. 2015, 110, 569–572. [Google Scholar] [CrossRef] [PubMed]
- Campos, G.S.; Bandeira, A.C.; Sardi, S.I. Zika Virus Outbreak, Bahia, Brazil. Emerg. Infect. Dis. 2015, 21, 1885–1886. [Google Scholar] [CrossRef]
- Miner, J.J.; Diamond, M.S. Zika Virus Pathogenesis and Tissue Tropism. Cell Host Microbe. 2017, 21, 134–142. [Google Scholar] [CrossRef]
- Moore, C.A.; Staples, J.E.; Dobyns, W.B.; Pessoa, A.; Ventura, C.V.; Fonseca, E.B.; Ribeiro, E.M.; Ventura, L.O.; Neto, N.N.; Arena, J.F.; et al. Characterizing the Pattern of Anomalies in Congenital Zika Syndrome for Pediatric Clinicians. JAMA Pediatr. 2017, 171, 288–295. [Google Scholar] [CrossRef]
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. Elife 2015, 4, e08347. [Google Scholar] [CrossRef]
- Haddow, A.J.; Williams, M.C.; Woodall, J.P.; Simpson, D.I.; Goma, L.K. Twelve Isolations of Zika Virus from Aedes (Stegomyia) Africanus (Theobald) Taken in and above a Uganda Forest. Bull. World Health Organ. 1964, 31, 57–69. [Google Scholar]
- Marchette, N.J.; Garcia, R.; Rudnick, A. Isolation of Zika virus from Aedes aegypti mosquitoes in Malaysia. Am. J. Trop. Med. Hyg. 1969, 18, 411–415. [Google Scholar] [CrossRef]
- Ferreira-de-Brito, A.; Ribeiro, I.P.; Miranda, R.M.; Fernandes, R.S.; Campos, S.S.; Silva, K.A.; Castro, M.G.; Bonaldo, M.C.; Brasil, P.; Lourenco-de-Oliveira, R. First detection of natural infection of Aedes aegypti with Zika virus in Brazil and throughout South America. Mem. Inst. Oswaldo. Cruz. 2016, 111, 655–658. [Google Scholar] [CrossRef] [PubMed]
- Guerbois, M.; Fernandez-Salas, I.; Azar, S.R.; Danis-Lozano, R.; Alpuche-Aranda, C.M.; Leal, G.; Garcia-Malo, I.R.; Diaz-Gonzalez, E.E.; Casas-Martinez, M.; Rossi, S.L.; et al. Outbreak of Zika Virus Infection, Chiapas State, Mexico, 2015, and First Confirmed Transmission by Aedes aegypti Mosquitoes in the Americas. J. Infect. Dis. 2016, 214, 1349–1356. [Google Scholar] [CrossRef] [PubMed]
- Ledermann, J.P.; Guillaumot, L.; Yug, L.; Saweyog, S.C.; Tided, M.; Machieng, P.; Pretrick, M.; Marfel, M.; Griggs, A.; Bel, M.; et al. Aedes hensilli as a potential vector of Chikungunya and Zika viruses. PLoS Negl. Trop. Dis. 2014, 8, e3188. [Google Scholar]
- Chouin-Carneiro, T.; Vega-Rua, A.; Vazeille, M.; Yebakima, A.; Girod, R.; Goindin, D.; Dupont-Rouzeyrol, M.; Lourenco-de-Oliveira, R.; Failloux, A.B. Differential Susceptibilities of Aedes aegypti and Aedes albopictus from the Americas to Zika Virus. PLoS Negl. Trop. Dis. 2016, 10, e0004543. [Google Scholar] [CrossRef]
- Gendernalik, A.; Weger-Lucarelli, J.; Garcia Luna, S.M.; Fauver, J.R.; Ruckert, C.; Murrieta, R.A.; Bergren, N.; Samaras, D.; Nguyen, C.; Kading, R.C.; et al. American Aedes vexans Mosquitoes are Competent Vectors of Zika Virus. Am. J. Trop. Med. Hyg. 2017, 96, 1338–1340. [Google Scholar] [CrossRef]
- Diallo, D.; Sall, A.A.; Diagne, C.T.; Faye, O.; Faye, O.; Ba, Y.; Hanley, K.A.; Buenemann, M.; Weaver, S.C.; Diallo, M. Zika virus emergence in mosquitoes in southeastern Senegal, 2011. PLoS ONE 2014, 9, e109442. [Google Scholar] [CrossRef]
- Elizondo-Quiroga, D.; Medina-Sanchez, A.; Sanchez-Gonzalez, J.M.; Eckert, K.A.; Villalobos-Sanchez, E.; Navarro-Zuniga, A.R.; Sanchez-Tejeda, G.; Correa-Morales, F.; Gonzalez-Acosta, C.; Arias, C.F.; et al. Zika Virus in Salivary Glands of Five Different Species of Wild-Caught Mosquitoes from Mexico. Sci. Rep. 2018, 8, 809. [Google Scholar] [CrossRef]
- Fu, S.; Song, S.; Liu, H.; Li, Y.; Li, X.; Gao, X.; Xu, Z.; Liu, G.; Wang, D.; Tian, Z.; et al. ZIKA virus isolated from mosquitoes: A field and laboratory investigation in China, 2016. Sci. China Life Sci. 2017, 60, 1364–1371. [Google Scholar] [CrossRef]
- Guedes, D.R.; Paiva, M.H.; Donato, M.M.; Barbosa, P.P.; Krokovsky, L.; Rocha, S.; Saraiva, K.; Crespo, M.M.; Rezende, T.M.; Wallau, G.L.; et al. Zika virus replication in the mosquito Culex quinquefasciatus in Brazil. Emerg. Microbes. Infect. 2017, 6, e69. [Google Scholar] [CrossRef]
- Guo, X.X.; Li, C.X.; Deng, Y.Q.; Xing, D.; Liu, Q.M.; Wu, Q.; Sun, A.J.; Dong, Y.D.; Cao, W.C.; Qin, C.F.; et al. Culex pipiens quinquefasciatus: A potential vector to transmit Zika virus. Emerg. Microbes. Infect. 2016, 5, e102. [Google Scholar]
- Smartt, C.T.; Shin, D.; Kang, S.; Tabachnick, W.J. Culex quinquefasciatus (Diptera: Culicidae) From Florida Transmitted Zika Virus. Front. Microbiol. 2018, 9, 768. [Google Scholar] [CrossRef] [PubMed]
- Farajollahi, A.; Fonseca, D.M.; Kramer, L.D.; Marm Kilpatrick, A. “Bird biting” mosquitoes and human disease: A review of the role of Culex pipiens complex mosquitoes in epidemiology. Infect. Genet. Evol. 2011, 11, 1577–1585. [Google Scholar] [CrossRef] [PubMed]
- Ciota, A.T.; Kramer, L.D. Vector-virus interactions and transmission dynamics of West Nile virus. Viruses 2013, 5, 3021–3047. [Google Scholar] [CrossRef] [PubMed]
- Alaniz, A.J.; Carvajal, M.A.; Bacigalupo, A.; Cattan, P.E. Global spatial assessment of Aedes aegypti and Culex quinquefasciatus: A scenario of Zika virus exposure. Epidemiol. Infect. 2018, 147, 1–11. [Google Scholar] [CrossRef]
- Byrne, K.; Nichols, R.A. Culex pipiens in London Underground tunnels: Differentiation between surface and subterranean populations. Heredity 1999, 82, 7–15. [Google Scholar] [CrossRef]
- Fros, J.J.; Geertsema, C.; Vogels, C.B.; Roosjen, P.P.; Failloux, A.B.; Vlak, J.M.; Koenraadt, C.J.; Takken, W.; Pijlman, G.P. West Nile Virus: High Transmission Rate in North-Western European Mosquitoes Indicates Its Epidemic Potential and Warrants Increased Surveillance. PLoS Negl. Trop. Dis. 2015, 9, e0003956. [Google Scholar] [CrossRef]
- Fros, J.J.; Miesen, P.; Vogels, C.B.; Gaibani, P.; Sambri, V.; Martina, B.E.; Koenraadt, C.J.; van Rij, R.P.; Vlak, J.M.; Takken, W.; et al. Comparative Usutu and West Nile virus transmission potential by local Culex pipiens mosquitoes in north-western Europe. One Health 2015, 1, 31–36. [Google Scholar] [CrossRef]
- Vogels, C.B.; Fros, J.J.; Goertz, G.P.; Pijlman, G.P.; Koenraadt, C.J. Vector competence of northern European Culex pipiens biotypes and hybrids for West Nile virus is differentially affected by temperature. Parasit. Vectors 2016, 9, 393. [Google Scholar] [CrossRef]
- Amraoui, F.; Atyame-Nten, C.; Vega-Rua, A.; Lourenco-de-Oliveira, R.; Vazeille, M.; Failloux, A.B. Culex mosquitoes are experimentally unable to transmit Zika virus. Euro. Surveill. 2016, 21, 30333. [Google Scholar] [CrossRef]
- Kenney, J.L.; Romo, H.; Duggal, N.K.; Tzeng, W.P.; Burkhalter, K.L.; Brault, A.C.; Savage, H.M. Transmission Incompetence of Culex quinquefasciatus and Culex pipiens pipiens from North America for Zika Virus. Am. J. Trop. Med. Hyg. 2017, 96, 1235–1240. [Google Scholar] [CrossRef] [PubMed]
- Boccolini, D.; Toma, L.; Di Luca, M.; Severini, F.; Romi, R.; Remoli, M.E.; Sabbatucci, M.; Venturi, G.; Rezza, G.; Fortuna, C. Experimental investigation of the susceptibility of Italian Culex pipiens mosquitoes to Zika virus infection. Euro. Surveill. 2016, 21, 30328. [Google Scholar] [CrossRef]
- Aliota, M.T.; Peinado, S.A.; Osorio, J.E.; Bartholomay, L.C. Culex pipiens and Aedes triseriatus Mosquito Susceptibility to Zika Virus. Emerg. Infect. Dis. 2016, 22, 1857–1859. [Google Scholar] [CrossRef] [PubMed]
- Weger-Lucarelli, J.; Ruckert, C.; Chotiwan, N.; Nguyen, C.; Garcia Luna, S.M.; Fauver, J.R.; Foy, B.D.; Perera, R.; Black, W.C.; Kading, R.C.; et al. Vector Competence of American Mosquitoes for Three Strains of Zika Virus. PLoS Negl. Trop. Dis. 2016, 10, e0005101. [Google Scholar] [CrossRef]
- Heitmann, A.; Jansen, S.; Luhken, R.; Leggewie, M.; Badusche, M.; Pluskota, B.; Becker, N.; Vapalahti, O.; Schmidt-Chanasit, J.; Tannich, E. Experimental transmission of Zika virus by mosquitoes from central Europe. Euro. Surveill. 2017, 22, 30437. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.J.; Ayers, V.B.; Lyons, A.C.; Unlu, I.; Alto, B.W.; Cohnstaedt, L.W.; Higgs, S.; Vanlandingham, D.L. Culex Species Mosquitoes and Zika Virus. Vector Borne Zoonotic Dis. 2016, 16, 673–676. [Google Scholar] [CrossRef] [PubMed]
- Goertz, G.P.; Vogels, C.B.F.; Geertsema, C.; Koenraadt, C.J.M.; Pijlman, G.P. Mosquito co-infection with Zika and chikungunya virus allows simultaneous transmission without affecting vector competence of Aedes aegypti. PLoS Negl. Trop. Dis. 2017, 11, e0005654. [Google Scholar] [CrossRef]
- Armbruster, P.; Hutchinson, R.A. Pupal mass and wing length as indicators of fecundity in Aedes albopictus and Aedes geniculatus (Diptera: Culicidae). J. Med. Entomol. 2002, 39, 699–704. [Google Scholar] [CrossRef]
- Vogels, C.B.; Goertz, G.P.; Pijlman, G.P.; Koenraadt, C.J. Vector competence of European mosquitoes for West Nile virus. Emerg. Microbes. Infect. 2017, 6, e96. [Google Scholar] [CrossRef]
- Gaunt, M.W.; Sall, A.A.; de Lamballerie, X.; Falconar, A.K.; Dzhivanian, T.I.; Gould, E.A. Phylogenetic relationships of flaviviruses correlate with their epidemiology, disease association and biogeography. J. Gen. Virol. 2001, 82, 1867–1876. [Google Scholar] [CrossRef]
- Grard, G.; Moureau, G.; Charrel, R.N.; Holmes, E.C.; Gould, E.A.; de Lamballerie, X. Genomics and evolution of Aedes-borne flaviviruses. J. Gen. Virol. 2010, 91, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Tsetsarkin, K.A.; Vanlandingham, D.L.; McGee, C.E.; Higgs, S. A single mutation in chikungunya virus affects vector specificity and epidemic potential. PLoS Pathog. 2007, 3, e201. [Google Scholar] [CrossRef] [PubMed]
- Stapleford, K.A.; Coffey, L.L.; Lay, S.; Borderia, A.V.; Duong, V.; Isakov, O.; Rozen-Gagnon, K.; Arias-Goeta, C.; Blanc, H.; Beaucourt, S.; et al. Emergence and transmission of arbovirus evolutionary intermediates with epidemic potential. Cell Host Microbe. 2014, 15, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Lourenco-de-Oliveira, R.; Marques, J.T.; Sreenu, V.B.; Atyame Nten, C.; Aguiar, E.; Varjak, M.; Kohl, A.; Failloux, A.B. Culex quinquefasciatus mosquitoes do not support replication of Zika virus. J. Gen. Virol. 2018, 99, 258–264. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abbo, S.R.; Vogels, C.B.F.; Visser, T.M.; Geertsema, C.; van Oers, M.M.; Koenraadt, C.J.M.; Pijlman, G.P. Forced Zika Virus Infection of Culex pipiens Leads to Limited Virus Accumulation in Mosquito Saliva. Viruses 2020, 12, 659. https://doi.org/10.3390/v12060659
Abbo SR, Vogels CBF, Visser TM, Geertsema C, van Oers MM, Koenraadt CJM, Pijlman GP. Forced Zika Virus Infection of Culex pipiens Leads to Limited Virus Accumulation in Mosquito Saliva. Viruses. 2020; 12(6):659. https://doi.org/10.3390/v12060659
Chicago/Turabian StyleAbbo, Sandra R., Chantal B. F. Vogels, Tessa M. Visser, Corinne Geertsema, Monique M. van Oers, Constantianus J. M. Koenraadt, and Gorben P. Pijlman. 2020. "Forced Zika Virus Infection of Culex pipiens Leads to Limited Virus Accumulation in Mosquito Saliva" Viruses 12, no. 6: 659. https://doi.org/10.3390/v12060659
APA StyleAbbo, S. R., Vogels, C. B. F., Visser, T. M., Geertsema, C., van Oers, M. M., Koenraadt, C. J. M., & Pijlman, G. P. (2020). Forced Zika Virus Infection of Culex pipiens Leads to Limited Virus Accumulation in Mosquito Saliva. Viruses, 12(6), 659. https://doi.org/10.3390/v12060659