Structural Characterization of Cuta- and Tusavirus: Insight into Protoparvoviruses Capsid Morphology

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
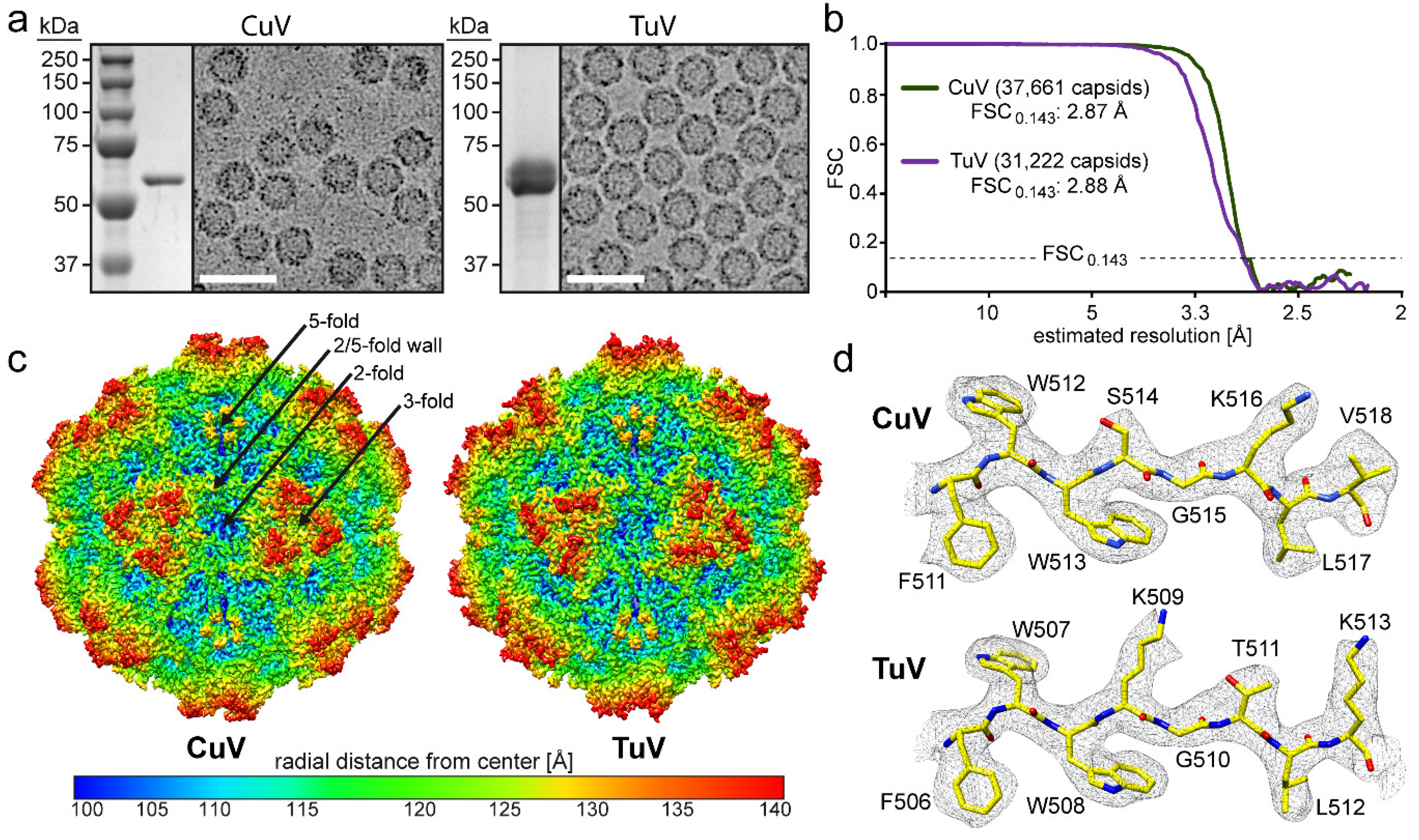
2.1. Production and Purification of CuV and TuV Virus-Like Particles
2.2. VLP Sample Purity and Integrity
2.3. Cryo-Electron Microscopy (Cryo-EM) and Image Reconstruction
2.4. Sequence and Structural Comparison
2.5. Fluorescent Labeling of VLPs
2.6. Cell Binding Assay
2.7. Glycan Array Screening
2.8. Protein Data Bank Accession Numbers
3. Results and Discussion
3.1. CuV and TuV VLPs Were Generated at Levels Suitable for Structure Determination
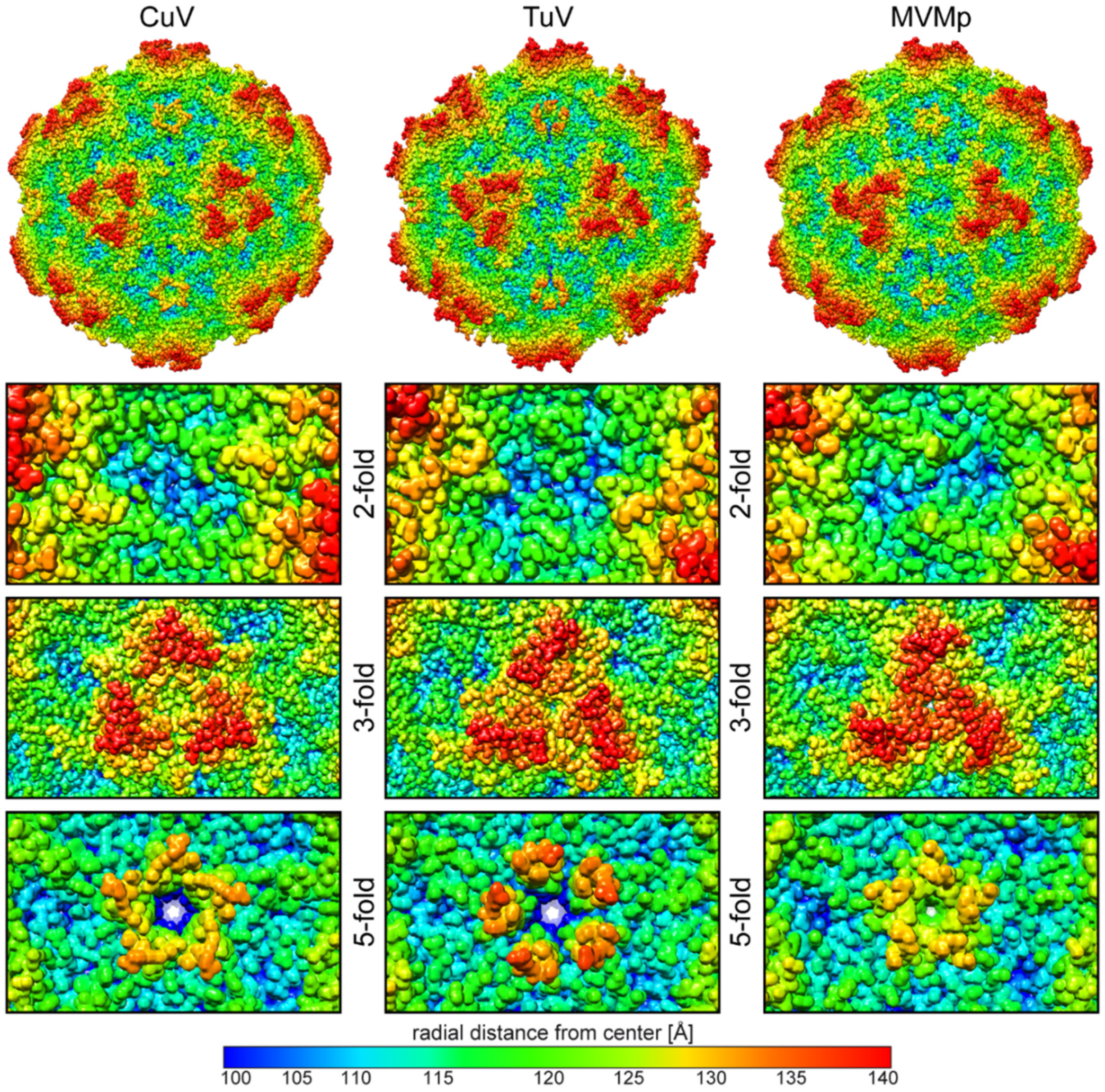
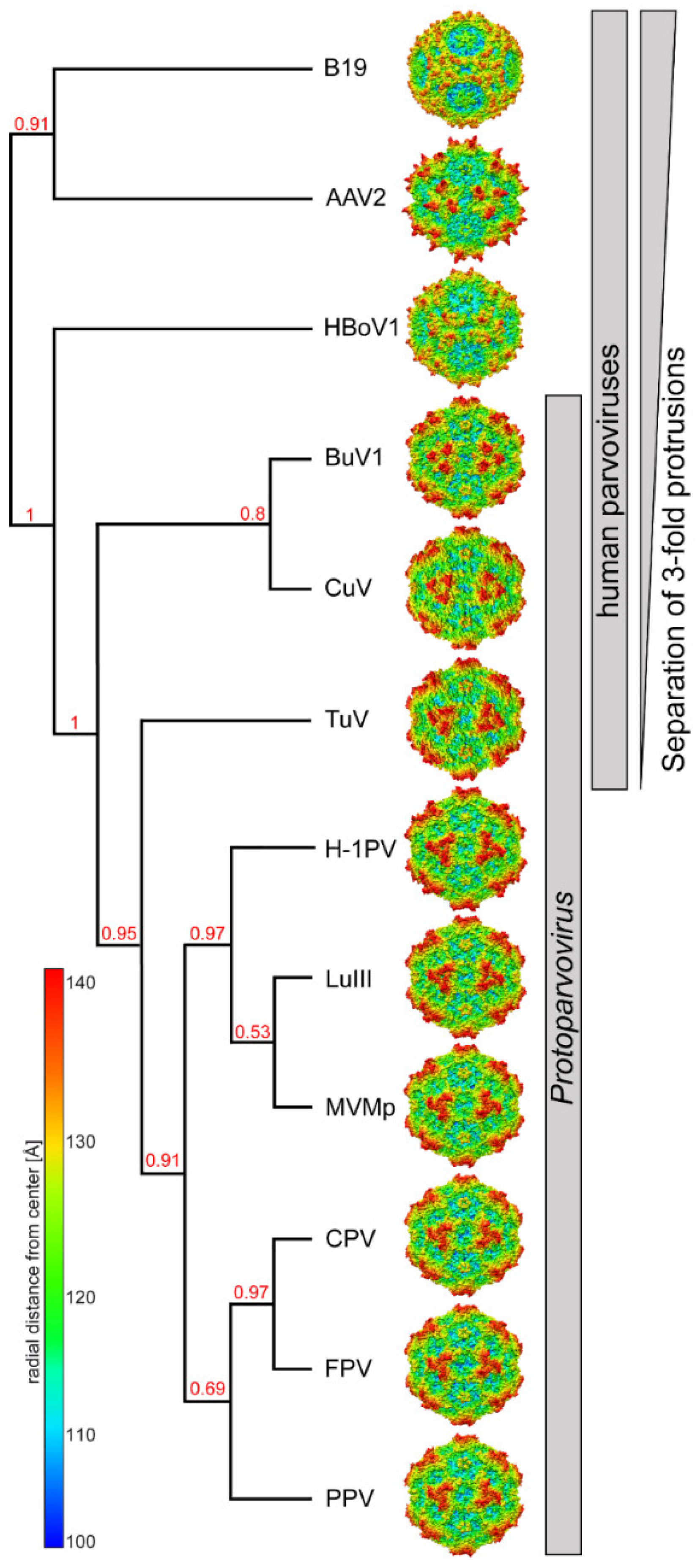
3.2. Comparison of CuV and TuV to other Protoparvoviruses Suggest Host-Driven Capsid Surface Morphologies
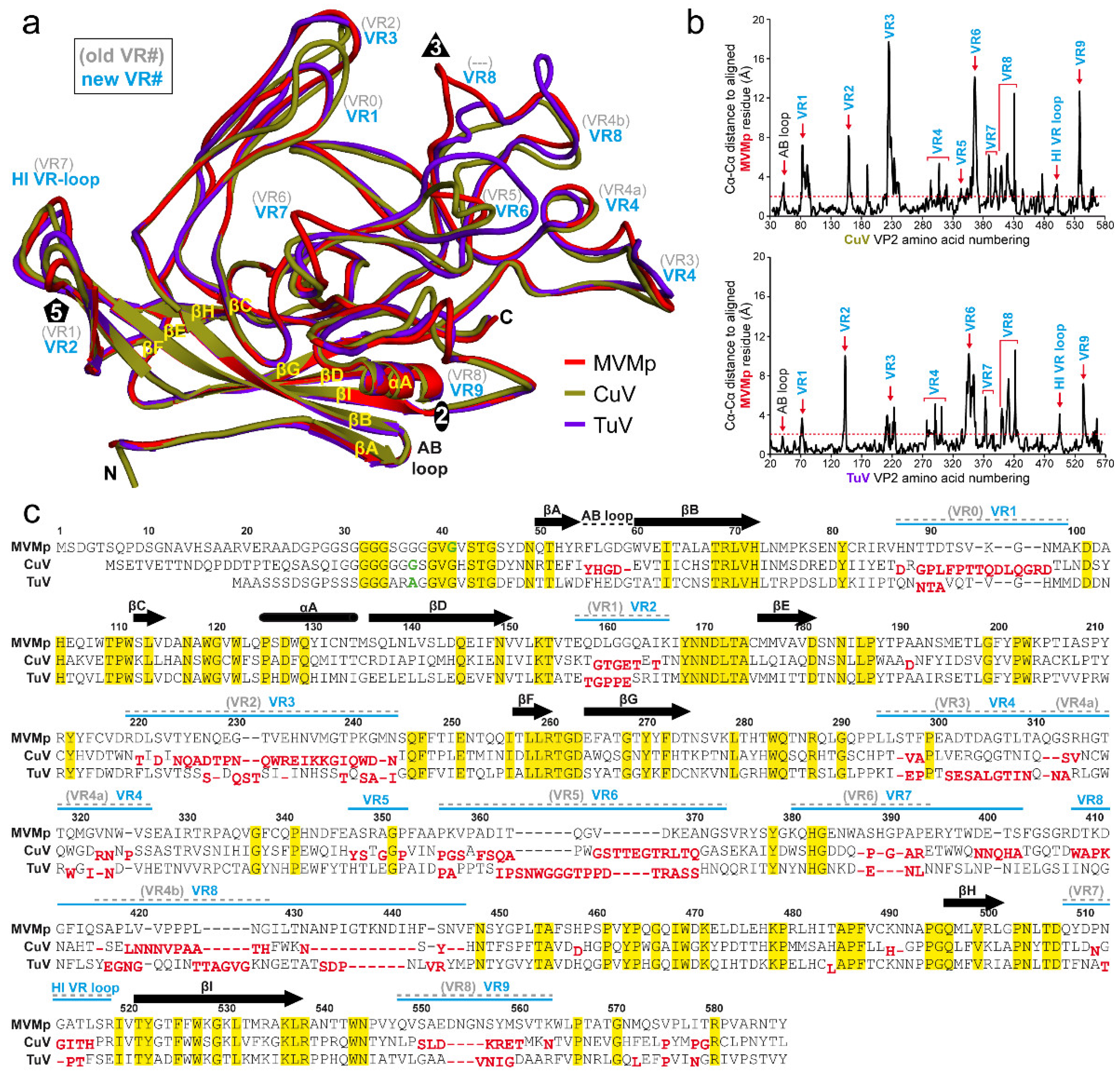
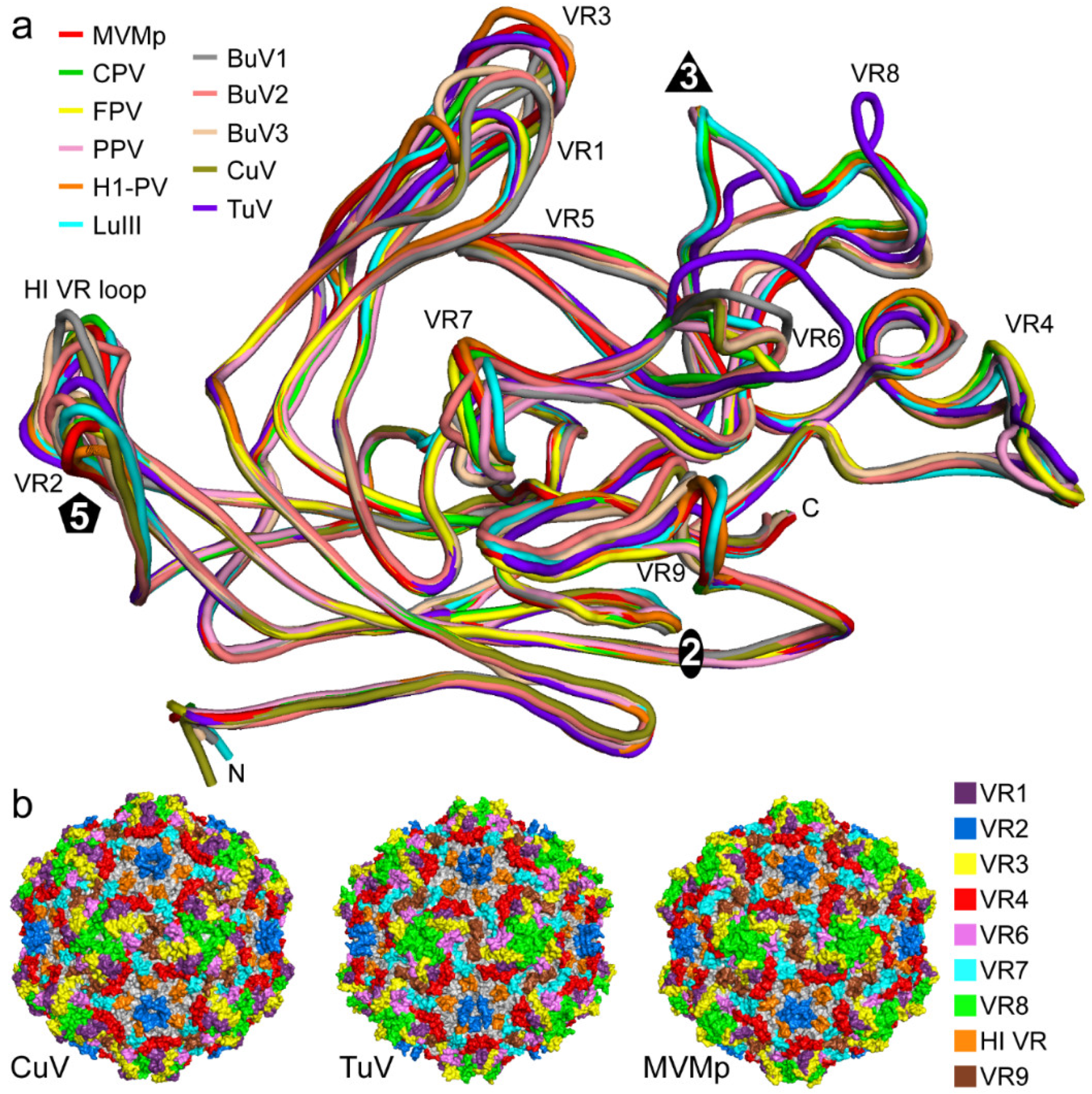
3.3. The CuV and TuV Capsids Exhibit the Most Differences in the Variable Regions of the Protoparvoviruses
3.4. BuV and CuV Are Closely Related
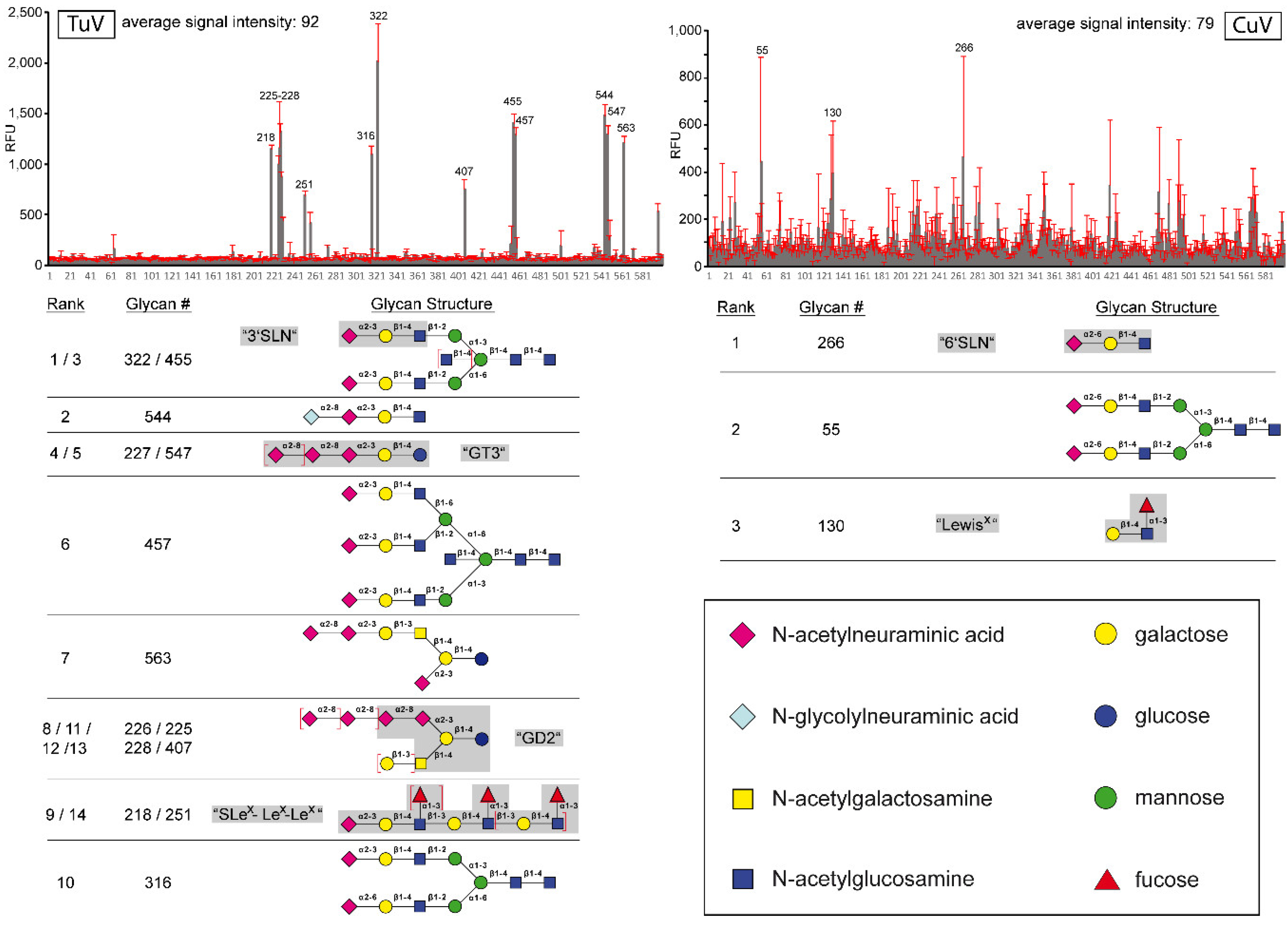
3.5. TuV Binds to α2-3-Linked and Poly SIA while CuV Potentially Binds α2-6-Linked SIA
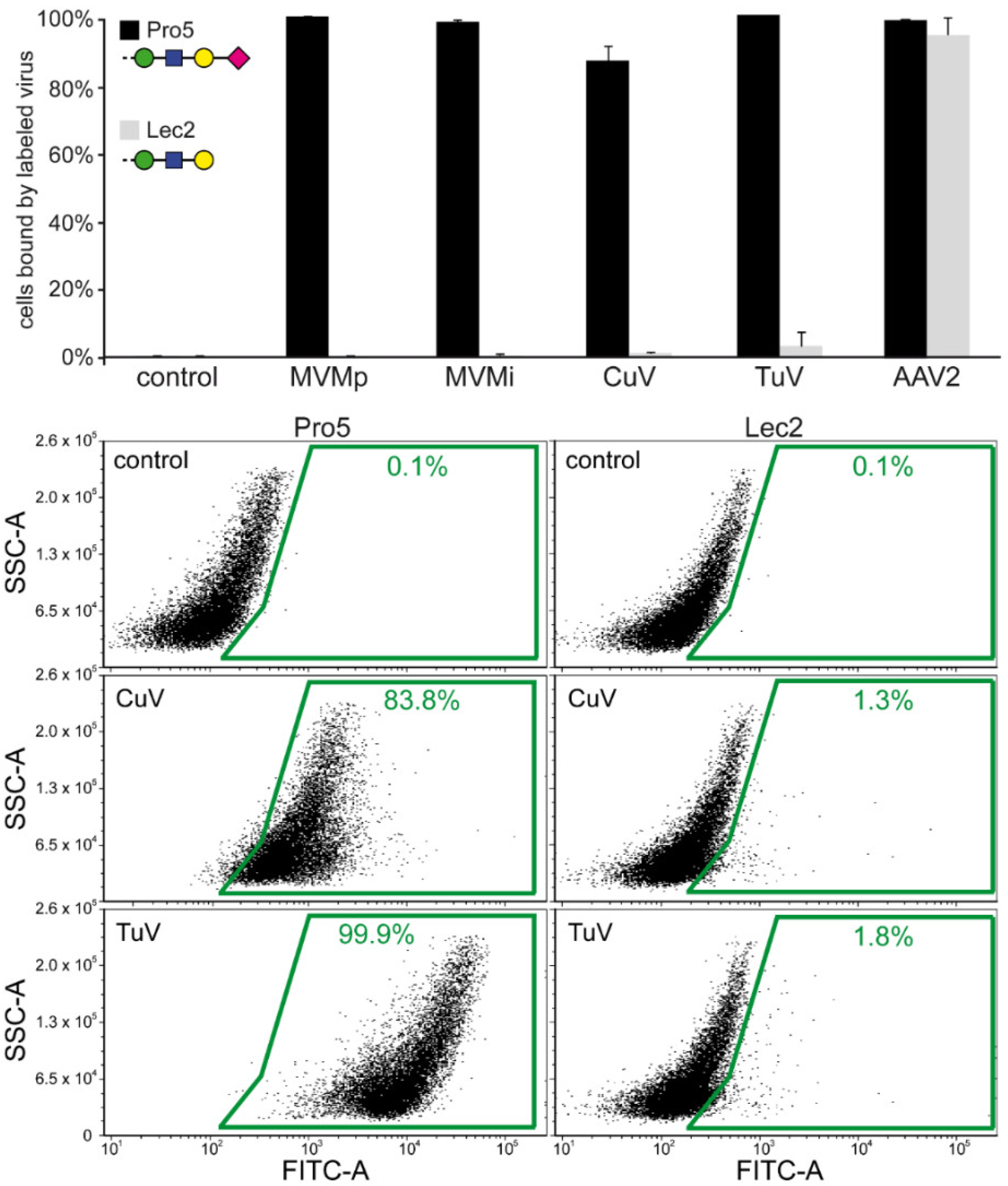
3.6. Cell Binding Assays Confirm that SIA Serves as a Cellular Attachment Receptor for CuV and TuV
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cotmore, S.F.; Agbandje-McKenna, M.; Canuti, M.; Chiorini, J.A.; Eis-Hubinger, A.-M.; Hughes, J.; Mietzsch, M.; Modha, S.; Ogliastro, M.; Pénzes, J.J.; et al. ICTV Virus Taxonomy Profile: Parvoviridae. J. Gen. Virol. 2019, 100, 367–368. [Google Scholar] [CrossRef] [PubMed]
- Pénzes, J.J.; Söderlund-Venermo, M.; Canuti, M.; Eis-Hübinger, A.M.; Hughes, J.; Cotmore, S.F.; Harrach, B. Reorganizing the family Parvoviridae: A revised taxonomy independent of the canonical approach based on host association. Arch. Virol. 2020, 1–14. [Google Scholar]
- Phan, T.G.; Sdiri-Loulizi, K.; Aouni, M.; Ambert-Balay, K.; Pothier, P.; Deng, X.; Delwart, E. New Parvovirus in Child with Unexplained Diarrhea, Tunisia. Emerg. Infect. Dis. 2014, 20, 1911–1913. [Google Scholar] [CrossRef]
- Phan, T.G.; Vo, N.P.; Bonkoungou, I.J.; Kapoor, A.; Barro, N.; O’Ryan, M.; Kapusinszky, B.; Wang, C.; Delwart, E. Acute diarrhea in West African children: Diverse enteric viruses and a novel parvovirus genus. J. Virol. 2012, 86, 11024–11030. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.G.; Dréno, B.; Da Costa, A.C.; Li, L.; Orlandi, P.; Deng, X.; Kapusinszky, B.; Siqueira, J.; Knol, A.-C.; Halary, F.; et al. A new protoparvovirus in human fecal samples and cutaneous T cell lymphomas (mycosis fungoides). Virology 2016, 496, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Mietzsch, M.; Pénzes, J.J.; Mc Kenna, A.-; Agbandje-McKenna, M. Twenty-Five Years of Structural Parvovirology. Viruses 2019, 11, 362. [Google Scholar] [CrossRef]
- Farr, G.A.; Cotmore, S.F.; Tattersall, P. VP2 cleavage and the leucine ring at the base of the fivefold cylinder control pH-dependent externalization of both the VP1 N terminus and the genome of minute virus of mice. J. Virol. 2006, 80, 161–171. [Google Scholar] [CrossRef]
- Bennett, A.; Mietzsch, M.; Agbandje-McKenna, M. Understanding capsid assembly and genome packaging for adeno-associated viruses. Future Virol. 2017, 12, 283–297. [Google Scholar] [CrossRef]
- Ilyas, M.; Mietzsch, M.; Kailasan, S.; Vaisanen, E.; Luo, M.; Chipman, P.; Smith, J.K.; Kurian, J.; Sousa, D.; McKenna, R.; et al. Atomic Resolution Structures of Human Bufaviruses Determined by Cryo-Electron Microscopy. Viruses 2018, 10, 22. [Google Scholar] [CrossRef]
- Agbandje-McKenna, M.; Llamas-Saiz, A.L.; Wang, F.; Tattersall, P.; Rossmann, M.G. Functional implications of the structure of the murine parvovirus, minute virus of mice. Structure 1998, 6, 1369–1381. [Google Scholar] [CrossRef]
- Mollerup, S.; Fridholm, H.; Vinner, L.; Kjartansdóttir, K.R.; Friis-Nielsen, J.; Asplund, M.; Herrera, J.A.R.; Steiniche, T.; Mourier, T.; Brunak, S.; et al. Cutavirus in Cutaneous Malignant Melanoma. Emerg. Infect. Dis. 2017, 23, 363–365. [Google Scholar] [CrossRef] [PubMed]
- Smits, S.L.; Schapendonk, C.M.; van Beek, J.; Vennema, H.; Schurch, A.C.; Schipper, D.; Bodewes, R.; Haagmans, B.L.; Osterhaus, A.D.; Koopmans, M.P. New viruses in idiopathic human diarrhea cases, the Netherlands. Emerg. Infect. Dis. 2014, 20, 1218–1222. [Google Scholar] [CrossRef] [PubMed]
- Vaisanen, E.; Kuisma, I.; Phan, T.G.; Delwart, E.; Lappalainen, M.; Tarkka, E.; Hedman, K.; Soderlund-Venermo, M. Bufavirus in feces of patients with gastroenteritis, Finland. Emerg. Infect. Dis. 2014, 20, 1077–1079. [Google Scholar] [CrossRef] [PubMed]
- Väisänen, E.; Mohanraj, U.; Kinnunen, P.M.; Jokelainen, P.; Al-Hello, H.; Barakat, A.M.; Sadeghi, M.; Jalilian, F.A.; Majlesi, A.; Masika, M.; et al. Global Distribution of Human Protoparvoviruses. Emerg. Infect. Dis. 2018, 24, 1292–1299. [Google Scholar] [CrossRef] [PubMed]
- Väisänen, E.; Paloniemi, M.; Kuisma, I.; Lithovius, V.; Kumar, A.; Franssila, R.; Ahmed, K.; Delwart, E.; Vesikari, T.; Hedman, K.; et al. Epidemiology of two human protoparvoviruses, bufavirus and tusavirus. Sci. Rep. 2016, 6, 39267. [Google Scholar] [CrossRef]
- Kreuter, A.; Nasserani, N.; Tigges, C.; Oellig, F.; Silling, S.; Akgul, B.; Wieland, U. Cutavirus Infection in Primary Cutaneous B- and T-Cell Lymphoma. JAMA Dermatol. 2018, 154, 965–967. [Google Scholar] [CrossRef]
- Väisänen, E.; Fu, Y.; Koskenmies, S.; Fyhrquist, N.; Wang, Y.; Keinonen, A.; Mäkisalo, H.; Väkevä, L.; Pitkänen, S.; Ranki, A.; et al. Cutavirus DNA in Malignant and Nonmalignant Skin of Cutaneous T-Cell Lymphoma and Organ Transplant Patients but Not of Healthy Adults. Clin. Infect. Dis. 2018, 68, 1904–1910. [Google Scholar] [CrossRef]
- Wieland, U.; Silling, S.; Hufbauer, M.; Mauch, C.; Zigrino, P.; Oellig, F.; Kreuter, A.; Akgul, B. No Evidence for Role of Cutavirus in Malignant Melanoma. Emerg. Infect. Dis. 2019, 25, 1600–16002. [Google Scholar] [CrossRef]
- Allaume, X.; El-Andaloussi, N.; Leuchs, B.; Bonifati, S.; Kulkarni, A.; Marttila, T.; Kaufmann, J.K.; Nettelbeck, D.M.; Kleinschmidt, J.; Rommelaere, J.; et al. Retargeting of rat parvovirus H-1PV to cancer cells through genetic engineering of the viral capsid. J. Virol. 2012, 86, 3452–3465. [Google Scholar] [CrossRef]
- Lofling, J.; Lyi, S.M.; Parrish, C.R.; Varki, A. Canine and feline parvoviruses preferentially recognize the non-human cell surface sialic acid N-glycolylneuraminic acid. Virology 2013, 440, 89–96. [Google Scholar] [CrossRef]
- Lopez-Bueno, A.; Rubio, M.P.; Bryant, N.; McKenna, R.; Agbandje-McKenna, M.; Almendral, J.M. Host-selected amino acid changes at the sialic acid binding pocket of the parvovirus capsid modulate cell binding affinity and determine virulence. J. Virol. 2006, 80, 1563–1573. [Google Scholar] [CrossRef] [PubMed]
- Boisvert, M.; Fernandes, S.; Tijssen, P. Multiple pathways involved in porcine parvovirus cellular entry and trafficking toward the nucleus. J. Virol. 2010, 84, 7782–7792. [Google Scholar] [CrossRef]
- Berger, I.; Poterszman, A. Baculovirus expression: Old dog, new tricks. Bioengineered 2015, 6, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.Q.; Palovcak, E.; Armache, J.P.; Verba, K.A.; Cheng, Y.; Agard, D.A. MotionCor2: Anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nat. Methods 2017, 14, 331–332. [Google Scholar] [CrossRef]
- Grant, T.; Rohou, A.; Grigorieff, N. cisTEM, user-friendly software for single-particle image processing. Elife 2018, 7, e35383. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Gallo Cassarino, T.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: Modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, W252–W258. [Google Scholar] [CrossRef]
- Ho, P.T.; Montiel-Garcia, D.J.; Wong, J.J.; Carrillo-Tripp, M.; Brooks, C.L.; Johnson, J.E.; Reddy, V.S. VIPERdb: A Tool for Virus Research. Annu. Rev. Virol. 2018, 5, 477–488. [Google Scholar] [CrossRef]
- Tang, G.; Peng, L.; Baldwin, P.R.; Mann, D.S.; Jiang, W.; Rees, I.; Ludtke, S.J. EMAN2: An extensible image processing suite for electron microscopy. J. Struct. Biol. 2007, 157, 38–46. [Google Scholar] [CrossRef]
- Adams, P.D.D.; Afonine, P.V.; Bunkóczi, G.; Chen, V.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.-W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. Sect. D Boil. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Mietzsch, M.; Broecker, F.; Reinhardt, A.; Seeberger, P.H.; Heilbronn, R.; Imperiale, M.J. Differential adeno-associated virus serotype-specific interaction patterns with synthetic heparins and other glycans. J. Virol. 2014, 88, 2991–3003. [Google Scholar] [CrossRef] [PubMed]
- Mietzsch, M.; Kailasan, S.; Garrison, J.; Ilyas, M.; Chipman, P.; Kantola, K.; Janssen, M.E.; Spear, J.; Sousa, D.; McKenna, R.; et al. Structural Insights into Human Bocaparvoviruses. J. Virol. 2017, 91, e00261-17. [Google Scholar] [CrossRef] [PubMed]
- Venkatakrishnan, B.; Yarbrough, J.; Domsic, J.; Bennett, A.; Bothner, B.; Kozyreva, O.G.; Samulski, R.J.; Muzyczka, N.; McKenna, R.; Agbandje-McKenna, M. Structure and dynamics of adeno-associated virus serotype 1 VP1-unique N-terminal domain and its role in capsid trafficking. J. Virol. 2013, 87, 4974–4984. [Google Scholar] [CrossRef]
- Mietzsch, M.; Barnes, C.; Hull, J.A.; Chipman, P.; Xie, J.; Bhattacharya, N.; Sousa, D.; McKenna, R.; Gao, G.; Agbandje-McKenna, M. Comparative Analysis of the Capsid Structures of AAVrh.10, AAVrh.39, and AAV8. J. Virol. 2020, 94, 6. [Google Scholar] [CrossRef]
- Burg, M.; Rosebrough, C.; Drouin, L.M.; Bennett, A.; Mietzsch, M.; Chipman, P.; McKenna, R.; Sousa, D.; Potter, M.; Byrne, B.; et al. Atomic structure of a rationally engineered gene delivery vector, AAV2.5. J. Struct. Biol. 2018, 203, 236–241. [Google Scholar] [CrossRef]
- Pittman, N.; Misseldine, A.; Geilen, L.; Halder, S.; Smith, J.K.; Kurian, J.; Chipman, P.; Janssen, M.; McKenna, R.; Baker, T.S.; et al. Atomic Resolution Structure of the Oncolytic Parvovirus LuIII by Electron Microscopy and 3D Image Reconstruction. Viruses 2017, 9, 321. [Google Scholar] [CrossRef]
- Wu, H.; Rossmann, M.G. The canine parvovirus empty capsid structure. J. Mol. Biol. 1993, 233, 231–244. [Google Scholar] [CrossRef]
- Halder, S.; Nam, H.J.; Govindasamy, L.; Vogel, M.; Dinsart, C.; Salome, N.; McKenna, R.; Agbandje-McKenna, M. Structural characterization of H-1 parvovirus: Comparison of infectious virions to empty capsids. J. Virol. 2013, 87, 5128–5140. [Google Scholar] [CrossRef]
- Simpson, A.A.; Hebert, B.; Sullivan, G.M.; Parrish, C.R.; Zadori, Z.; Tijssen, P.; Rossmann, M.G. The structure of porcine parvovirus: Comparison with related viruses. J. Mol. Biol. 2002, 315, 1189–1198. [Google Scholar] [CrossRef]
- Kontou, M.; Govindasamy, L.; Nam, H.J.; Bryant, N.; Llamas-Saiz, A.L.; Foces-Foces, C.; Hernando, E.; Rubio, M.P.; McKenna, R.; Almendral, J.M.; et al. Structural determinants of tissue tropism and in vivo pathogenicity for the parvovirus minute virus of mice. J. Virol. 2005, 79, 10931–10943. [Google Scholar] [CrossRef] [PubMed]
- Agbandje, M.; McKenna, R.; Rossmann, M.G.; Strassheim, M.L.; Parrish, C.R. Structure determination of feline panleukopenia virus empty particles. Proteins Struct. Funct. Bioinform. 1993, 16, 155–171. [Google Scholar] [CrossRef]
- Kaufmann, B.; Lopez-Bueno, A.; Mateu, M.G.; Chipman, P.R.; Nelson, C.D.; Parrish, C.R.; Almendral, J.M.; Rossmann, M.G. Minute virus of mice, a parvovirus, in complex with the Fab fragment of a neutralizing monoclonal antibody. J. Virol. 2007, 81, 9851–9858. [Google Scholar] [CrossRef] [PubMed]
- Plevka, P.; Hafenstein, S.; Li, L.; D’Abrgamo, A., Jr.; Cotmore, S.F.; Rossmann, M.G.; Tattersall, P. Structure of a packaging-defective mutant of minute virus of mice indicates that the genome is packaged via a pore at a 5-fold axis. J. Virol. 2011, 85, 4822–4827. [Google Scholar] [CrossRef]
- DeLano, W.L. The PyMOL Molecular Graphics Syste; DeLano Scientific: San Carlos, CA, USA, 2002. [Google Scholar]
- Farr, G.A.; Tattersall, P. A conserved leucine that constricts the pore through the capsid fivefold cylinder plays a central role in parvoviral infection. Virology 2004, 323, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Reguera, J.; Carreira, A.; Riolobos, L.; Almendral, J.M.; Mateu, M.G. Role of interfacial amino acid residues in assembly, stability, and conformation of a spherical virus capsid. Proc. Natl. Acad. Sci. USA 2004, 101, 2724–2729. [Google Scholar] [CrossRef] [PubMed]
- Hafenstein, S.; Bowman, V.D.; Sun, T.; Nelson, C.D.; Palermo, L.M.; Chipman, P.R.; Battisti, A.J.; Parrish, C.R.; Rossmann, M.G. Structural comparison of different antibodies interacting with parvovirus capsids. J. Virol. 2009, 83, 5556–5566. [Google Scholar] [CrossRef]
- Emmanuel, S.N.; Mietzsch, M.; Tseng, Y.S.; Smith, J.K.; Agbandje-McKenna, M. Parvovirus Capsid-Antibody Complex Structures Reveal Conservation of Antigenic Epitopes across the Family. Viral Immunol. 2020. [Google Scholar] [CrossRef]
- Goodman, L.B.; Lyi, S.M.; Johnson, N.C.; Cifuente, J.O.; Hafenstein, S.L.; Parrish, C.R. Binding site on the transferrin receptor for the parvovirus capsid and effects of altered affinity on cell uptake and infection. J. Virol. 2010, 84, 4969–4978. [Google Scholar] [CrossRef] [PubMed]
- Hafenstein, S.; Palermo, L.M.; Kostyuchenko, V.A.; Xiao, C.; Morais, M.C.; Nelson, C.D.; Bowman, V.D.; Battisti, A.J.; Chipman, P.R.; Parrish, C.R.; et al. Asymmetric binding of transferrin receptor to parvovirus capsids. Proc. Natl. Acad. Sci. USA 2007, 104, 6585–6589. [Google Scholar] [CrossRef] [PubMed]
- Tsao, J.; Chapman, M.S.; Agbandje, M.; Keller, W.; Smith, K.; Wu, H.; Luo, M.; Smith, T.J.; Rossmann, M.G.; Compans, R.W.; et al. The three-dimensional structure of canine parvovirus and its functional implications. Science 1991, 251, 1456–1464. [Google Scholar] [CrossRef] [PubMed]
- Llamas-Saiz, A.L.; Agbandje-McKenna, M.; Wikoff, W.R.; Bratton, J.; Tattersall, P.; Rossmann, M.G. Structure determination of minute virus of mice. Acta Crystallogr. Sect. D Biol. Crystallogr. 1997, 53, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Nam, H.J.; Gurda-Whitaker, B.; Gan, W.Y.; Ilaria, S.; McKenna, R.; Mehta, P.; Alvarez, R.A.; Agbandje-McKenna, M. Identification of the sialic acid structures recognized by minute virus of mice and the role of binding affinity in virulence adaptation. J. Biol. Chem. 2006, 281, 25670–25677. [Google Scholar] [CrossRef] [PubMed]
- Stanley, P.; Cummings, R.D. Structures Common to Different Glycans. In Essentials of Glycobiology, 3rd ed.; Varki, A., Cummings, R.D., Esko, J.D., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2017. [Google Scholar]
- Fujita, T.; Murayama, K.; Hanamura, T.; Okada, T.; Ito, T.; Harada, M.; Komatsu, A.; Koyama, H.; Kanai, T.; Maeno, K.; et al. CSLEX (Sialyl Lewis X) is a Useful Tumor Marker for Monitoring of Breast Cancer Patients. Jpn. J. Clin. Oncol. 2010, 41, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Miller, E.; Agbandje-McKenna, M.; Samulski, R.J. Alpha2,3 and alpha2,6 N-linked sialic acids facilitate efficient binding and transduction by adeno-associated virus types 1 and 6. J. Virol. 2006, 80, 9093–9103. [Google Scholar] [CrossRef] [PubMed]
- Kaludov, N.; Brown, K.E.; Walters, R.W.; Zabner, J.; Chiorini, J.A. Adeno-associated virus serotype 4 (AAV4) and AAV5 both require sialic acid binding for hemagglutination and efficient transduction but differ in sialic acid linkage specificity. J. Virol. 2001, 75, 6884–6893. [Google Scholar] [CrossRef]
- North, S.J.; Huang, H.H.; Sundaram, S.; Jang-Lee, J.; Etienne, A.T.; Trollope, A.; Chalabi, S.; Dell, A.; Stanley, P.; Haslam, S.M. Glycomics profiling of Chinese hamster ovary cell glycosylation mutants reveals N-glycans of a novel size and complexity. J. Biol. Chem. 2010, 285, 5759–5775. [Google Scholar] [CrossRef]
- Barbis, D.P.; Chang, S.F.; Parrish, C.R. Mutations adjacent to the dimple of the canine parvovirus capsid structure affect sialic acid binding. Virology 1992, 191, 301–308. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cryo-EM Data and Refinement Parameter | CuV | TuV |
| Total number of micrographs | 738 | 580 |
| Defocus range (µm) | 0.90–3.84 | 0.83–4.27 |
| Electron dose (e-/Å2) | 75 | 75 |
| Frames / micrograph | 50 | 50 |
| Pixel size (Å/pixel) | 1.05 | 1.07 |
| Capsids used for final map | 15,296 | 33,191 |
| Resolution of final map (Å) | 2.87 | 2.88 |
| PHENIX model refinement statistics | ||
| Residue range | 32–569 | 19–565 |
| Map CC | 0.874 | 0.866 |
| RMSD bonds (Å) | 0.01 | 0.01 |
| RMSD angles (°) | 0.86 | 1.01 |
| All-atom clash score | 8.10 | 7.66 |
| Ramachandran plot | ||
| Favored (%) | 97.8 | 95.6 |
| Allowed (%) | 2.2 | 4.4 |
| Outliers (%) | 0 | 0 |
| Rotamer outliers (%) | 0.2 | 0.2 |
| C-β deviations | 0 | 0 |
| RMSD [Å] | Overall | w/o VRs | AB- loop | VR1 | VR2 | VR3 | VR4 | VR5 | VR6 | VR7 | VR8 | HI- loop | VR9 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CuV vs. MVMp | 3.0 | 1.0 | 2.5 | 4.1 | 4.1 | 6.6 | 2.2 | 2.0 | 5.9 | 3.1 | 5.2 | 2.0 | 6.3 |
| TuV vs. MVMp | 2.2 | 0.8 | 1.1 | 1.6 | 4.4 | 2.2 | 2.4 | 0.9 | 5.4 | 2.6 | 4.0 | 2.2 | 3.8 |
| CuV vs. TuV | 2.7 | 1.0 | 2.9 | 3.3 | 2.3 | 6.7 | 1.9 | 1.4 | 6.9 | 1.4 | 3.7 | 2.3 | 3.8 |
| BuV1 | BuV2 | BuV3 | CuV | TuV | MVMp | LuIII | H-1PV | CPV | FPV | PPV | Structural aligned amino acids: Cα distance < 2Å (in %) | |
| BuV1 | - | 91.4 | 98.0 | 95.6 | 79.8 | 70.8 | 72.2 | 71.4 | 69.7 | 70.4 | 71.5 | |
| BuV2 | 66.0 | - | 92.0 | 93.9 | 74.4 | 65.6 | 67.8 | 65.1 | 65.8 | 65.2 | 65.1 | |
| BuV3 | 74.4 | 65.2 | - | 95.4 | 78.1 | 70.0 | 69.0 | 67.8 | 68.5 | 70.4 | 70.1 | |
| CuV | 67.0 | 83.6 | 64.7 | - | 79.3 | 72.1 | 71.8 | 71.4 | 71.5 | 70.8 | 69.9 | |
| TuV | 35.1 | 32.5 | 33.6 | 33.3 | - | 80.4 | 81.9 | 79.9 | 80.4 | 80.7 | 80.7 | |
| MVMp | 31.2 | 32.1 | 31.9 | 31.8 | 41.4 | - | 96.9 | 97.1 | 88.9 | 91.2 | 90.8 | |
| LuIII | 32.6 | 32.2 | 31.6 | 33.3 | 43.0 | 72.2 | - | 96.1 | 87.7 | 89.4 | 90.8 | |
| H-1PV | 32.9 | 31.2 | 32.9 | 30.4 | 41.6 | 67.4 | 68.4 | - | 88.7 | 91.6 | 90.9 | |
| CPV | 33.6 | 33.2 | 33.3 | 32.2 | 40.4 | 52.2 | 53.3 | 52.9 | - | 96.2 | 89.9 | |
| FPV | 32.3 | 32.3 | 32.6 | 31.8 | 40.4 | 52.4 | 53.1 | 52.5 | 98.0 | - | 90.0 | |
| PPV | 33.3 | 34.6 | 34.0 | 32.3 | 42.6 | 51.3 | 51.1 | 51.4 | 59.2 | 58.8 | - | |
| Amino acid sequence identity in ordered VP structure (in %) | ||||||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mietzsch, M.; McKenna, R.; Väisänen, E.; Yu, J.C.; Ilyas, M.; Hull, J.A.; Kurian, J.; Smith, J.K.; Chipman, P.; Lasanajak, Y.; et al. Structural Characterization of Cuta- and Tusavirus: Insight into Protoparvoviruses Capsid Morphology. Viruses 2020, 12, 653. https://doi.org/10.3390/v12060653
Mietzsch M, McKenna R, Väisänen E, Yu JC, Ilyas M, Hull JA, Kurian J, Smith JK, Chipman P, Lasanajak Y, et al. Structural Characterization of Cuta- and Tusavirus: Insight into Protoparvoviruses Capsid Morphology. Viruses. 2020; 12(6):653. https://doi.org/10.3390/v12060653
Chicago/Turabian StyleMietzsch, Mario, Robert McKenna, Elina Väisänen, Jennifer C. Yu, Maria Ilyas, Joshua A. Hull, Justin Kurian, J. Kennon Smith, Paul Chipman, Yi Lasanajak, and et al. 2020. "Structural Characterization of Cuta- and Tusavirus: Insight into Protoparvoviruses Capsid Morphology" Viruses 12, no. 6: 653. https://doi.org/10.3390/v12060653
APA StyleMietzsch, M., McKenna, R., Väisänen, E., Yu, J. C., Ilyas, M., Hull, J. A., Kurian, J., Smith, J. K., Chipman, P., Lasanajak, Y., Smith, D., Söderlund-Venermo, M., & Agbandje-McKenna, M. (2020). Structural Characterization of Cuta- and Tusavirus: Insight into Protoparvoviruses Capsid Morphology. Viruses, 12(6), 653. https://doi.org/10.3390/v12060653