Detection of Astrovirus in a Cow with Neurological Signs by Nanopore Technology, Italy


 , , , , , and
, , , , , and 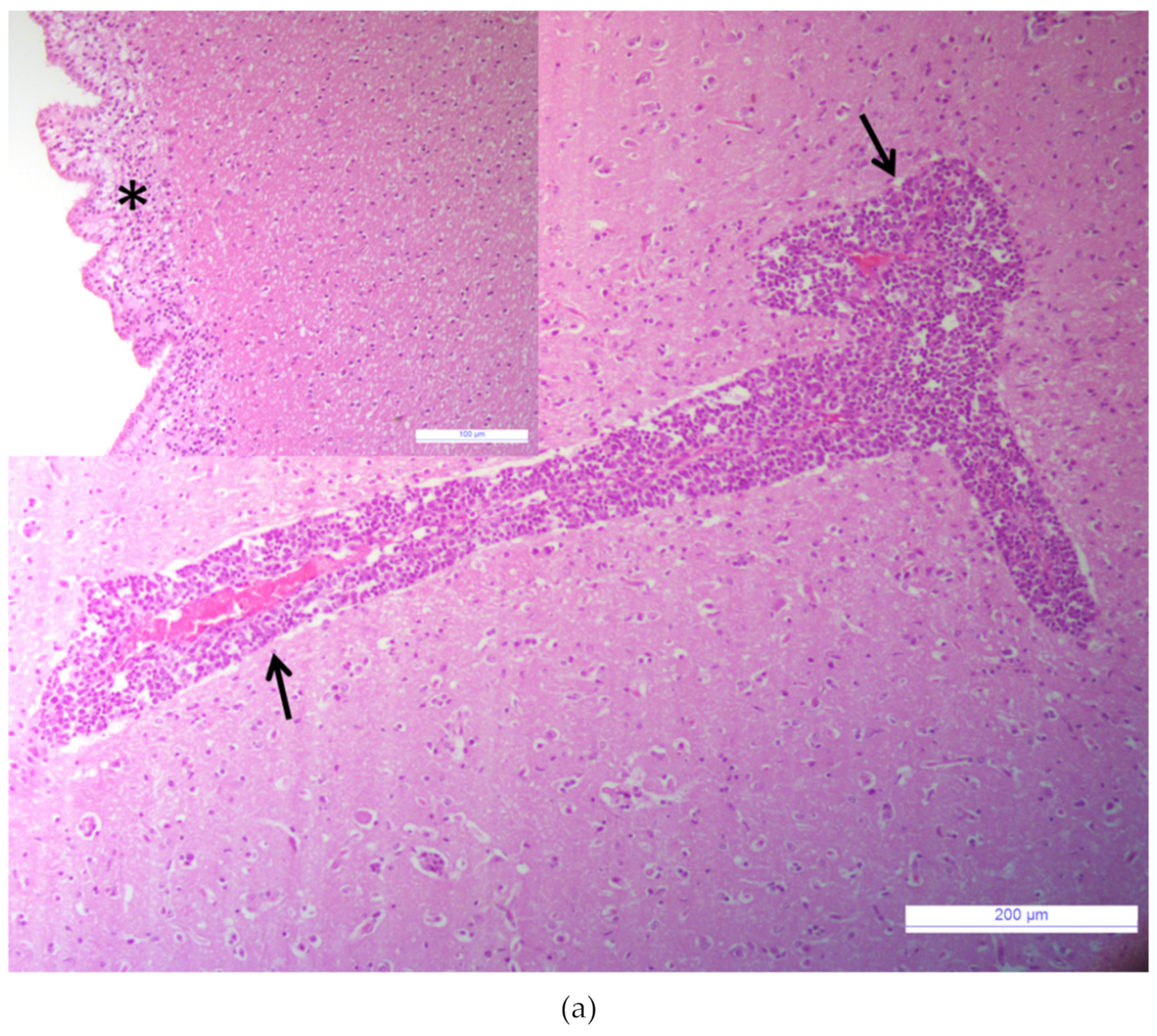{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Case Description
2.2. Samples Collection and Diagnostic Investigation
2.3. Shotgun Metagenomics by MinION
2.4. Real-Time RT-qPCRBoAstV
2.5. Shotgun Metagenomics by Illumina and Bioinformatic Analysis
2.6. Genome Characterization and Phylogeny
2.7. In Situ Investigation
3. Results
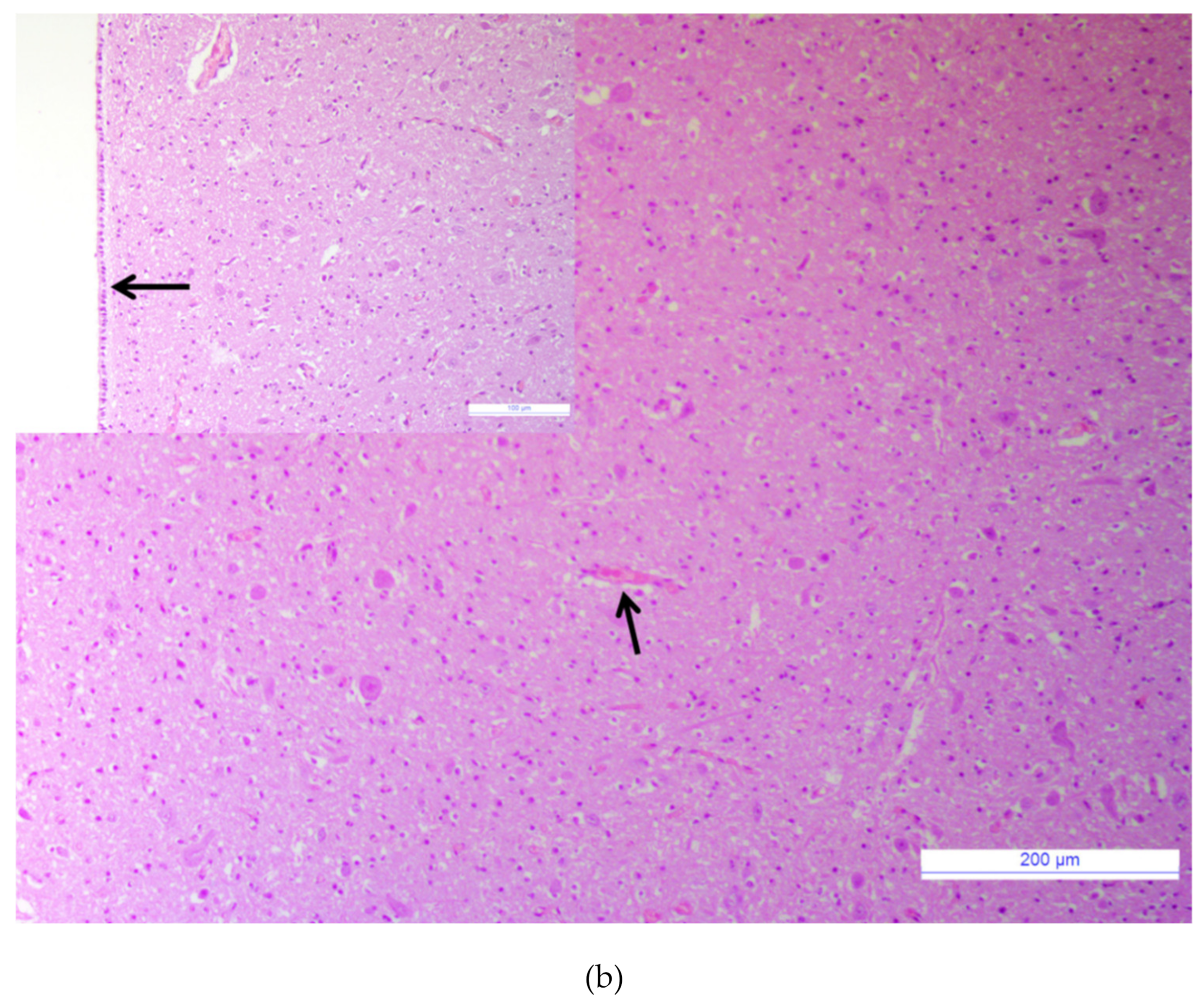
3.1. Major Neurotropic Pathogens Were Absent and Acute Non-Suppurative Encephalomyelitis Was Evidenced
3.2. Bovine Astrovirus RNA Was Identified by MinION and Confirmed by RT-qPCRBoAstV
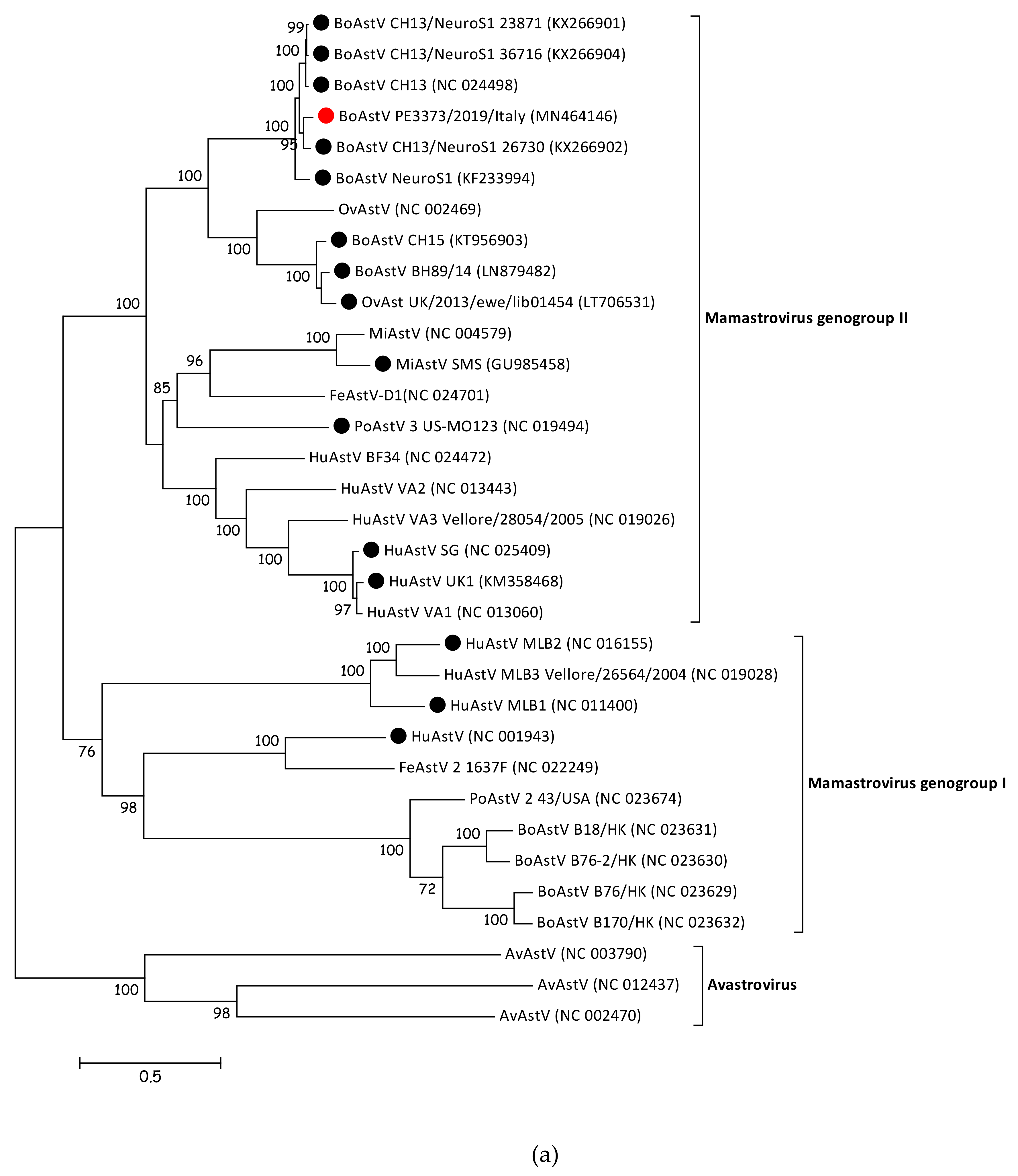
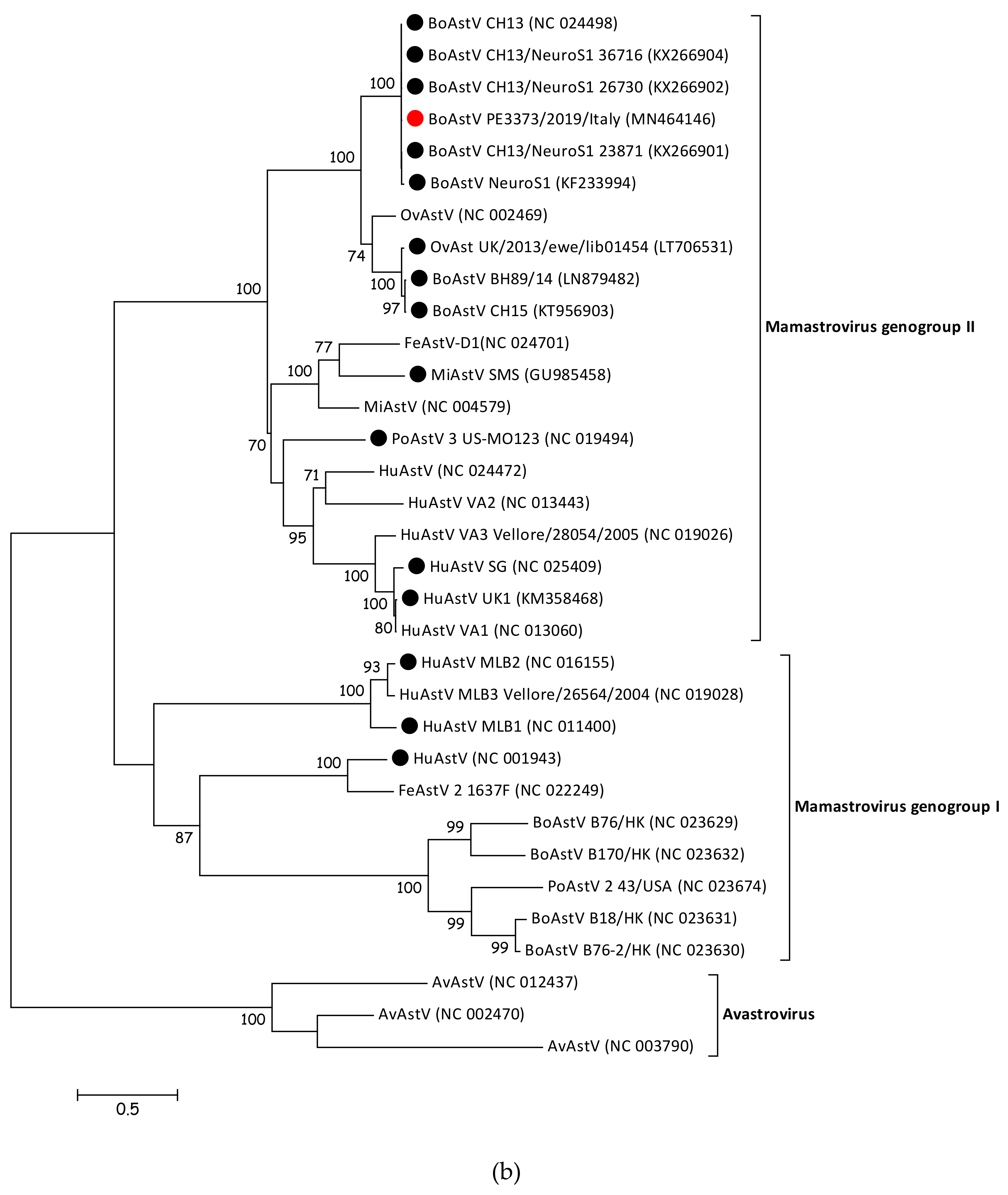
3.3. BoAstV PE3373/2019/Italy Showed the Highest nt Sequence Identity with BoAstV CH13/NeuroS1
3.4. BoAstV PE3373/2019/Italy Groups with BoAstV CH13/NeuroS1 Strains
3.5. BoAstV PE3373/2019/Italy Presence in Cells Was Confirmed by ISH
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Matsui, S.M.; Greenber, H.B. Astroviruses. In Fields Virology, 3rd ed.; Felds, B.N., Knipe, D.M., Howely, P.M., Chanock, R.M., Melnick, J.L., Monath, T.P., Roizman, B., Straus, S.E., Eds.; Lippincott-Raven Publishers: Philadelphia, PA, USA, 1996; pp. 811–824. [Google Scholar]
- De Benedictis, P.; Schultz-Cherry, S.; Burnham, A.; Cattoli, G. Astrovirus infections in humans and animals – Molecular biology, genetic diversity, and interspecies transmissions. Infect. Genet. Evol. 2011, 11, 1529–1544. [Google Scholar] [CrossRef] [PubMed]
- Koci, M.D.; Schultz-Cherry, S. Avian astroviruses. Avian Pathol. 2002, 31, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.; Pintó, R.M.; Guix, S. Human Astroviruses. Clin. Microbiol. Rev. 2014, 27, 1048–1074. [Google Scholar] [CrossRef] [PubMed]
- Lewis, T.L.; Matsui, S.M. Studies of the astrovirus signal that induces (−1) ribosomal frameshifting. Adv. Exp. Med. Biol. 1997, 412, 323–330. [Google Scholar] [CrossRef]
- Kiang, D.; Matsui, S.M. Proteolytic processing of a human astrovirus nonstructural protein. J. Gen. Virol. 2002, 83, 25–34. [Google Scholar] [CrossRef]
- Méndez, E.; Fernández-Luna, T.; López, S.; Méndez-Toss, M.; Arias, C.F. Proteolytic Processing of a Serotype 8 Human Astrovirus ORF2 Polyprotein. J. Virol. 2002, 76, 7996–8002. [Google Scholar] [CrossRef]
- Walter, J.E.; Mitchell, D.K. Astrovirus infection in children. Curr. Opin. Infect. Dis. 2003, 16, 247–253. [Google Scholar] [CrossRef]
- Appleton, H.; Higgins, P.G. Letter: Viruses and gastroenteritis in infants. Lancet 1975, 1, 1297. [Google Scholar] [CrossRef]
- Quan, P.-L.; Wagner, T.A.; Briese, T.; Torgerson, T.R.; Hornig, M.; Tashmukhamedova, A.; Firth, C.; Palacios, G.; Baisre-De-Leon, A.; Paddock, C.D.; et al. Astrovirus Encephalitis in Boy with X-linked Agammaglobulinemia. Emerg. Infect. Dis. 2010, 16, 918–925. [Google Scholar] [CrossRef]
- Brown, J.R.; Morfopoulou, S.; Hubb, J.; Emmett, W.A.; Ip, W.; Shah, D.; Brooks, T.; Paine, S.M.L.; Anderson, G.; Virasami, A.; et al. Astrovirus VA1/HMO-C: An Increasingly Recognized Neurotropic Pathogen in Immunocompromised Patients. Clin. Infect. Dis. 2015, 60, 881–888. [Google Scholar] [CrossRef]
- Frémond, M.-L.; Pérot, P.; Muth, E.; Cros, G.; Dumarest, M.; Mahlaoui, N.; Seilhean, D.; Desguerre, I.; Hébert, C.; Corre-Catelin, N.; et al. Next-Generation Sequencing for Diagnosis and Tailored Therapy: A Case Report of Astrovirus-Associated Progressive Encephalitis. J. Pediatr. Infect. Dis. Soc. 2015, 4, e53–e57. [Google Scholar] [CrossRef] [PubMed]
- Naccache, S.N.; Peggs, K.S.; Mattes, F.M.; Phadke, R.; Garson, J.A.; Grant, P.; Samayoa, E.; Federman, S.; Miller, S.; Lunn, M.P.; et al. Diagnosis of Neuroinvasive Astrovirus Infection in an Immunocompromised Adult With Encephalitis by Unbiased Next-Generation Sequencing. Clin. Infect. Dis. 2015, 60, 919–923. [Google Scholar] [CrossRef] [PubMed]
- Lum, S.H.; Turner, A.; Guiver, M.; Bonney, D.; Martland, T.; Davies, E.; Newbould, M.; Brown, J.; Morfopoulou, S.; Breuer, J.; et al. An emerging opportunistic infection: Fatal astrovirus (VA1/HMO-C) encephalitis in a pediatric stem cell transplant recipient. Transpl. Infect. Dis. 2016, 18, 960–964. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Diab, S.; McGraw, S.; Barr, B.; Traslavina, R.; Higgins, R.; Talbot, T.; Blanchard, P.; Rimoldi, G.; Fahsbender, E.; et al. Divergent Astrovirus Associated with Neurologic Disease in Cattle. Emerg. Infect. Dis. 2013, 19, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Bouzalas, I.G.; Wüthrich, D.; Walland, J.; Drögemüller, C.; Zurbriggen, A.; Vandevelde, M.; Oevermann, A.; Bruggmann, R.; Seuberlich, T. Neurotropic Astrovirus in Cattle with Nonsuppurative Encephalitis in Europe. J. Clin. Microbiol. 2014, 52, 3318–3324. [Google Scholar] [CrossRef] [PubMed]
- Bouzalas, I.G.; Wüthrich, D.; Selimovic-Hamza, S.; Drögemüller, C.; Bruggmann, R.; Seuberlich, T. Full-genome based molecular characterization of encephalitis-associated bovine astroviruses. Infect. Genet. Evol. 2016, 44, 162–168. [Google Scholar] [CrossRef]
- Schlottau, K.; Schulze, C.; Bilk, S.; Hanke, D.; Höper, D.; Beer, M.; Hoffmann, B. Detection of a Novel Bovine Astrovirus in a Cow with Encephalitis. Transbound. Emerg. Dis. 2016, 63, 253–259. [Google Scholar] [CrossRef]
- Seuberlich, T.; Wüthrich, D.; Selimovic-Hamza, S.; Drögemüller, C.; Oevermann, A.; Bruggmann, R.; Bouzalas, I. Identification of a second encephalitis-associated astrovirus in cattle. Emerg. Microbes Infect. 2016, 5, 1–3. [Google Scholar] [CrossRef]
- Blomström, A.-L.; Widén, F.; Hammer, A.-S.; Belák, S.; Berg, M. Detection of a Novel Astrovirus in Brain Tissue of Mink Suffering from Shaking Mink Syndrome by Use of Viral Metagenomics. J. Clin. Microbiol. 2010, 48, 4392–4396. [Google Scholar] [CrossRef]
- Boujon, C.L.; Koch, M.C.; Kauer, R.V.; Keller-Gautschi, E.; Hierweger, M.M.; Hoby, S.; Seuberlich, T. Novel encephalomyelitis-associated astrovirus in a muskox (Ovibos moschatus): A surprise from the archives. Acta Vet. Scand. 2019, 61, 31. [Google Scholar] [CrossRef]
- Boujon, C.L.; Koch, M.C.; Wüthrich, D.; Werder, S.; Jakupovic, D.; Bruggmann, R.; Seuberlich, T. Indication of Cross-Species Transmission of Astrovirus Associated with Encephalitis in Sheep and Cattle. Emerg. Infect. Dis. 2017, 23, 1604–1606. [Google Scholar] [CrossRef] [PubMed]
- Pfaff, F.; Schlottau, K.; Scholes, S.; Courtenay, A.; Hoffmann, B.; Höper, D.; Beer, M. A novel astrovirus associated with encephalitis and ganglionitis in domestic sheep. Transbound. Emerg. Dis. 2017, 64, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Selimovic-Hamza, S.; Boujon, C.L.; Hilbe, M.; Oevermann, A.; Seuberlich, T. Frequency and pathological phenotype of bovine astrovirus CH13/NeuroS1 infection in neurologically-diseased cattle: Towards assessment of causality. Viruses 2017, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Reuter, G.; Pankovics, P.; Boros, A. Nonsuppurative (Aseptic) Meningoencephalomyelitis Associated with Neurovirulent Astrovirus Infections in Humans and Animals. Clin. Microbiol. Rev. 2018, 31, e00040-18. [Google Scholar] [CrossRef] [PubMed]
- Vandevelde, M.; Higgins, R.; Oevermann, A. Veterinary Neuropathology: Essentials of Theory and Practice; John Wiley & Sons: Hoboken, NJ, USA, 2012; ISBN 978-1-118-34949-6. [Google Scholar]
- Peserico, A.; Marcacci, M.; Malatesta, D.; Di Domenico, M.; Pratelli, A.; Mangone, I.; D’Alterio, N.; Pizzurro, F.; Cirone, F.; Zaccaria, G.; et al. Diagnosis and characterization of canine distemper virus through sequencing by MinION nanopore technology. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Del Amo, J.; Sotelo, E.; Fernández-Pinero, J.; Gallardo, C.; Llorente, F.; Agüero, M.; Jiménez-Clavero, M.A. A novel quantitative multiplex real-time RT-PCR for the simultaneous detection and differentiation of West Nile virus lineages 1 and 2, and of Usutu virus. J. Virol. Methods 2013, 189, 321–327. [Google Scholar] [CrossRef]
- Cavrini, F.; Pepa, M.E.D.; Gaibani, P.; Pierro, A.M.; Rossini, G.; Landini, M.P.; Sambri, V. A rapid and specific real-time RT-PCR assay to identify Usutu virus in human plasma, serum, and cerebrospinal fluid. J. Clin. Virol. 2011, 50, 221–223. [Google Scholar] [CrossRef]
- Beato, M.S.; Marcacci, M.; Schiavon, E.; Bertocchi, L.; Di Domenico, M.; Peserico, A.; Mion, M.; Zaccaria, G.; Cavicchio, L.; Mangone, I.; et al. Identification and genetic characterization of bovine enterovirus by combination of two next generation sequencing platforms. J. Virol. Methods 2018, 260, 21–25. [Google Scholar] [CrossRef]
- Marcacci, M.; De Luca, E.; Zaccaria, G.; Di Tommaso, M.; Mangone, I.; Aste, G.; Savini, G.; Boari, A.; Lorusso, A. Genome characterization of feline morbillivirus from Italy. J. Virol. Methods 2016, 234, 160–163. [Google Scholar] [CrossRef]
- Marcacci, M.; Sant, S.; Mangone, I.; Goria, M.; Dondo, A.; Zoppi, S.; van Gennip, R.G.P.; Radaelli, M.C.; Cammà, C.; van Rijn, P.A.; et al. One after the other: A novel Bluetongue virus strain related to Toggenburg virus detected in the Piedmont region (North-western Italy), extends the panel of novel atypical BTV strains. Transbound. Emerg. Dis. 2018, 65, 370–374. [Google Scholar] [CrossRef]
- Lüthi, R.; Boujon, C.L.; Kauer, R.; Koch, M.C.; Bouzalas, I.G.; Seuberlich, T. Accurate and precise real-time RT-PCR assays for the identification of astrovirus associated encephalitis in cattle. Sci. Rep. 2018, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.A.; Korobeynikov, A.; Lapidus, A.; Prjibelski, A.D.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; et al. Assembling single-cell genomes and mini-metagenomes from chimeric MDA products. J. Comput. Biol. 2013, 20, 714–737. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Deiss, R.; Selimovic-Hamza, S.; Seuberlich, T.; Meylan, M. Neurologic Clinical Signs in Cattle with Astrovirus-Associated Encephalitis. J. Vet. Intern. Med. 2017, 31, 1209–1214. [Google Scholar] [CrossRef]
- Xu, Y.; Lewandowski, K.; Lumley, S.; Pullan, S.; Vipond, R.; Carroll, M.; Foster, D.; Matthews, P.C.; Peto, T.; Crook, D. Detection of Viral Pathogens with Multiplex Nanopore MinION Sequencing: Be Careful with Cross-Talk. Front. Microbiol. 2018, 9, 2225. [Google Scholar] [CrossRef]
- Bányai, K.; Meleg, E.; Moschidou, P.; Martella, V. Detection of Newly Described Astrovirus MLB1 in Stool Samples from Children. Emerg. Infect. Dis. 2010, 16, 169–170. [Google Scholar] [CrossRef]
- Martella, V.; Moschidou, P.; Pinto, P.; Catella, C.; Desario, C.; Larocca, V.; Circella, E.; Bànyai, K.; Lavazza, A.; Magistrali, C.; et al. Astroviruses in Rabbits. Emerg. Infect. Dis. 2011, 17, 2287–2293. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Moschidou, P.; Lorusso, E.; Mari, V.; Camero, M.; Bellacicco, A.; Losurdo, M.; Pinto, P.; Desario, C.; Bányai, K.; et al. Detection and characterization of canine astroviruses. J. Gen. Virol. 2011, 92, 1880–1887. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Moschidou, P.; Catella, C.; Larocca, V.; Pinto, P.; Losurdo, M.; Corrente, M.; Lorusso, E.; Bànyai, K.; Decaro, N.; et al. Enteric Disease in Dogs Naturally Infected by a Novel Canine Astrovirus. J. Clin. Microbiol. 2012, 50, 1066–1069. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Decaro, N.; Buonavoglia, C. Enteric viral infections in lambs or kids. Vet. Microbiol. 2015, 181, 154–160. [Google Scholar] [CrossRef]
- Jiang, H.; Holtz, L.R.; Bauer, I.; Franz, C.J.; Zhao, G.; Bodhidatta, L.; Shrestha, S.K.; Kang, G.; Wang, D. Comparison of novel MLB-clade, VA-clade and classic human astroviruses highlights constrained evolution of the classic human astrovirus nonstructural genes. Virology 2013, 436, 8–14. [Google Scholar] [CrossRef]
- Tse, H.; Chan, W.-M.; Tsoi, H.-W.; Fan, R.Y.Y.; Lau, C.C.Y.; Lau, S.K.P.; Woo, P.C.Y.; Yuen, K.-Y. Rediscovery and genomic characterization of bovine astroviruses. J. Gen. Virol. 2011, 92, 1888–1898. [Google Scholar] [CrossRef]
- Finkbeiner, S.R.; Le, B.-M.; Holtz, L.R.; Storch, G.A.; Wang, D. Detection of Newly Described Astrovirus MLB1 in Stool Samples from Children. Emerg. Infect. Dis. 2009, 15, 441–444. [Google Scholar] [CrossRef]
- Kapoor, A.; Li, L.; Victoria, J.; Oderinde, B.; Mason, C.; Pandey, P.; Zaidi, S.Z.; Delwart, E. Multiple novel astrovirus species in human stool. J. Gen. Virol. 2009, 90, 2965–2972. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaccaria, G.; Lorusso, A.; Hierweger, M.M.; Malatesta, D.; Defourny, S.V.; Ruggeri, F.; Cammà, C.; Ricci, P.; Di Domenico, M.; Rinaldi, A.; et al. Detection of Astrovirus in a Cow with Neurological Signs by Nanopore Technology, Italy. Viruses 2020, 12, 530. https://doi.org/10.3390/v12050530
Zaccaria G, Lorusso A, Hierweger MM, Malatesta D, Defourny SV, Ruggeri F, Cammà C, Ricci P, Di Domenico M, Rinaldi A, et al. Detection of Astrovirus in a Cow with Neurological Signs by Nanopore Technology, Italy. Viruses. 2020; 12(5):530. https://doi.org/10.3390/v12050530
Chicago/Turabian StyleZaccaria, Guendalina, Alessio Lorusso, Melanie M. Hierweger, Daniela Malatesta, Sabrina VP Defourny, Franco Ruggeri, Cesare Cammà, Pasquale Ricci, Marco Di Domenico, Antonio Rinaldi, and et al. 2020. "Detection of Astrovirus in a Cow with Neurological Signs by Nanopore Technology, Italy" Viruses 12, no. 5: 530. https://doi.org/10.3390/v12050530
APA StyleZaccaria, G., Lorusso, A., Hierweger, M. M., Malatesta, D., Defourny, S. V., Ruggeri, F., Cammà, C., Ricci, P., Di Domenico, M., Rinaldi, A., Decaro, N., D’Alterio, N., Petrini, A., Seuberlich, T., & Marcacci, M. (2020). Detection of Astrovirus in a Cow with Neurological Signs by Nanopore Technology, Italy. Viruses, 12(5), 530. https://doi.org/10.3390/v12050530