
First Evidence of Carp Edema Virus Infection of Koi Cyprinus carpio in Chiang Mai Province, Thailand

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Disease History and Clinical Examination
2.2. Histopathological Study
2.3. Electron Microscopic Study
2.4. Primers and PCR Amplification of CEV
2.5. DNA Sequence and Analysis of Sequence Data
3. Results
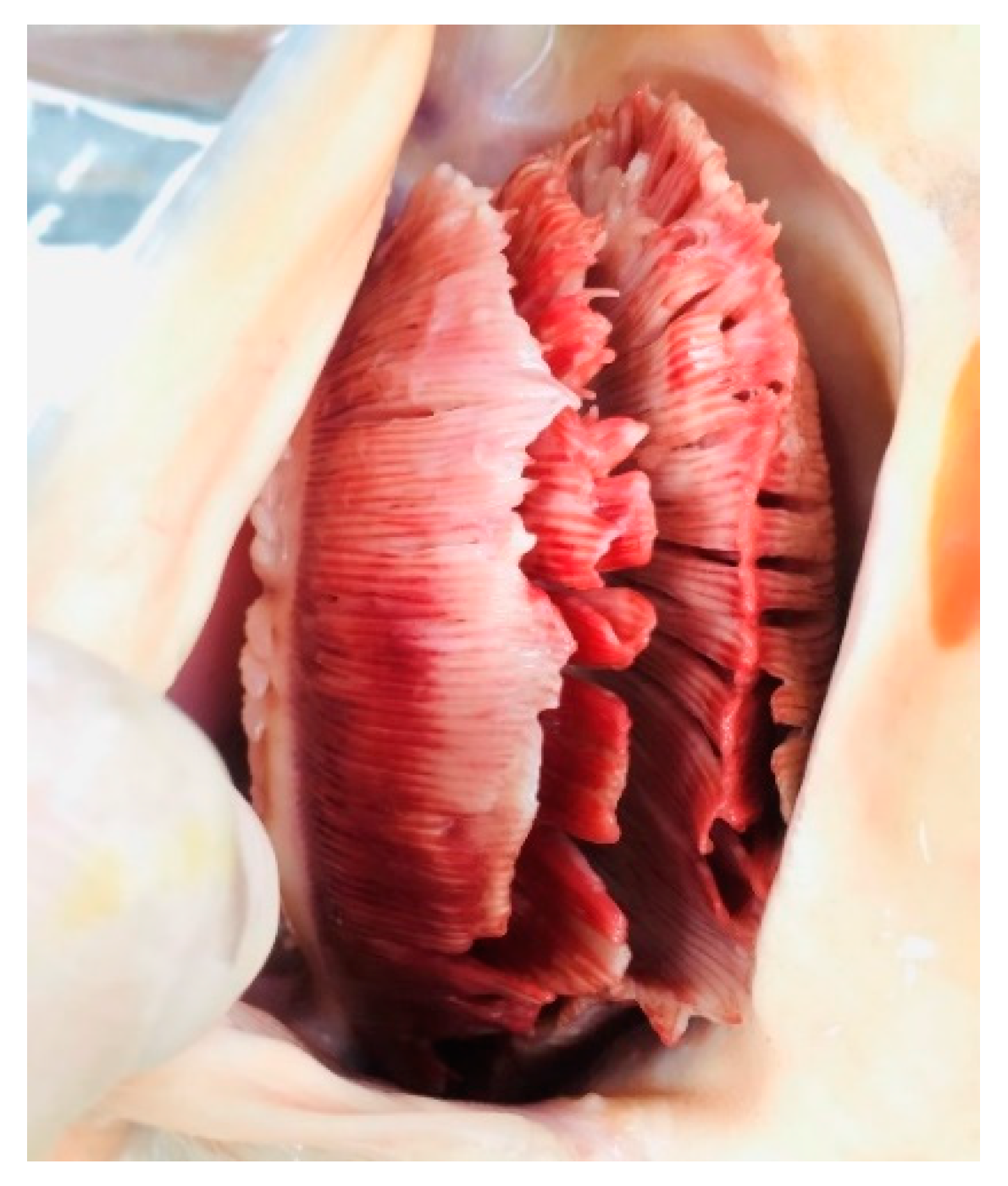
3.1. Disease History and Clinical Examination
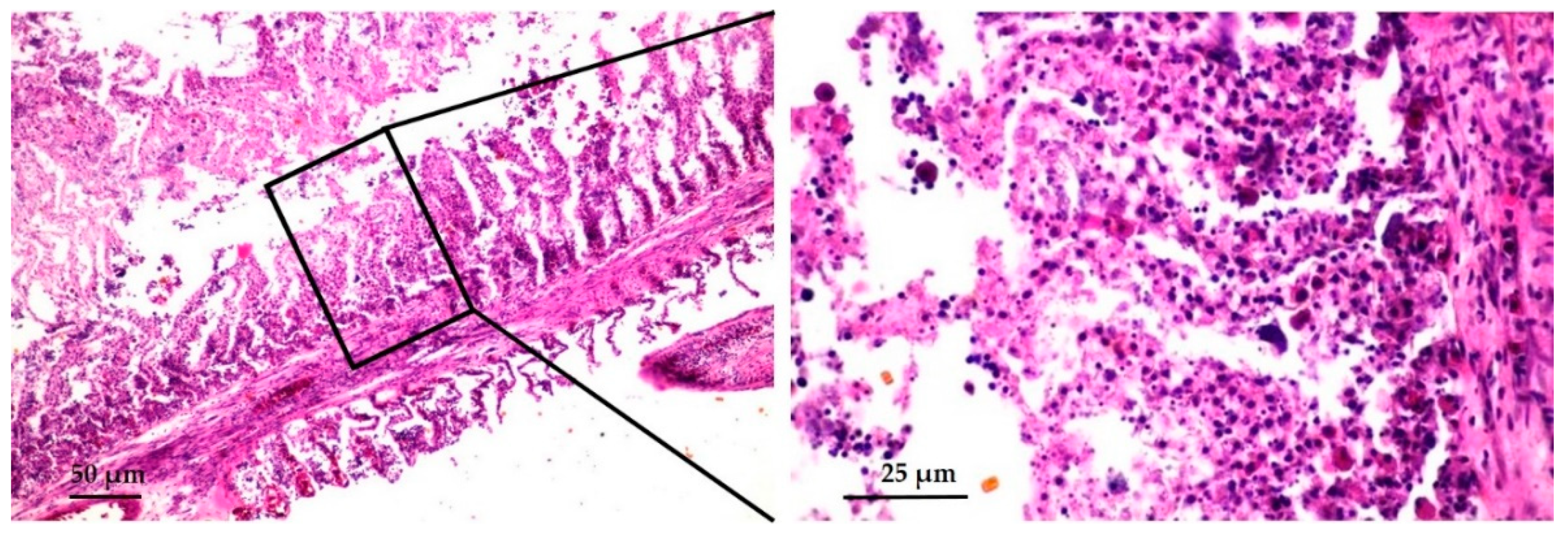
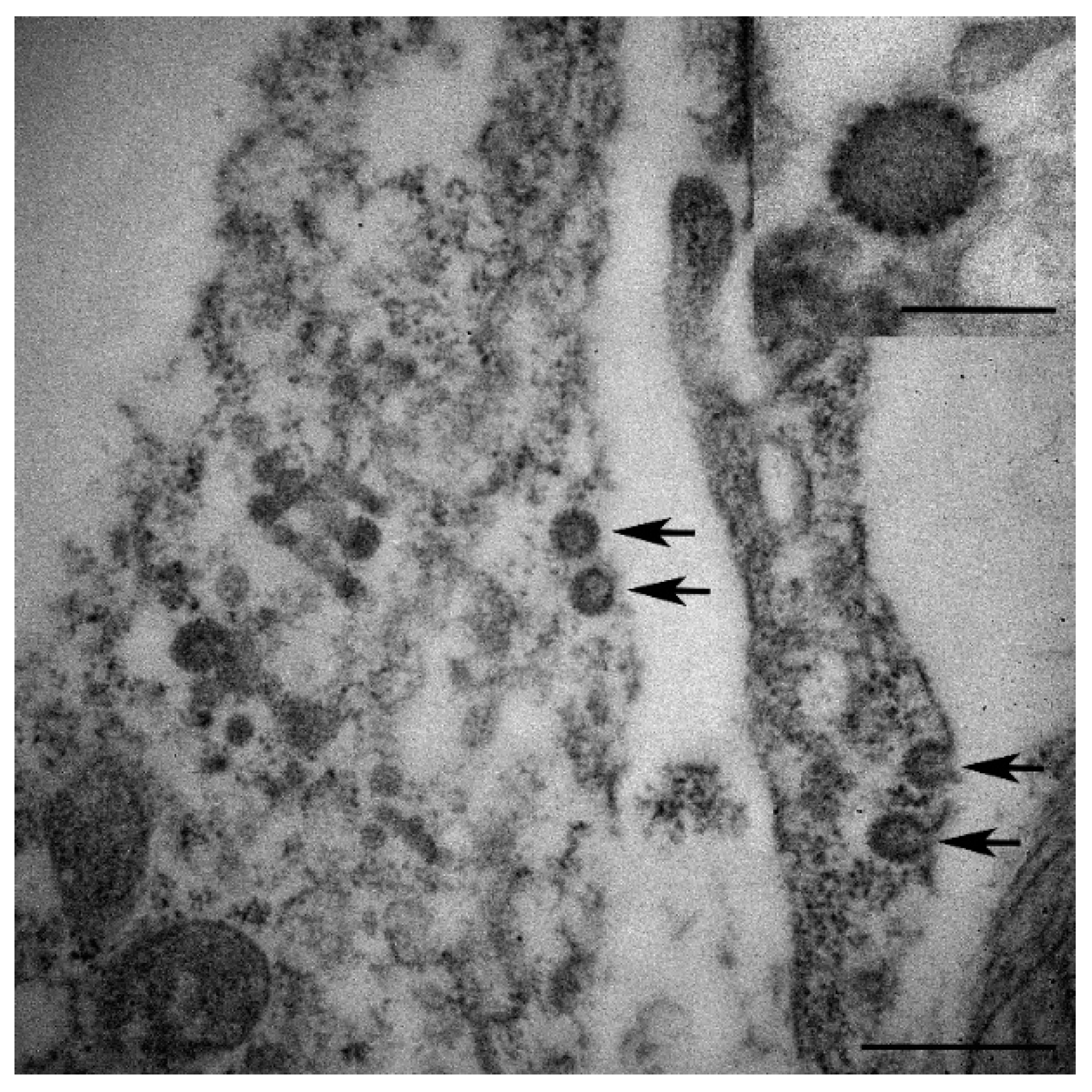
3.2. Histopathological and Electron Microscopic Study
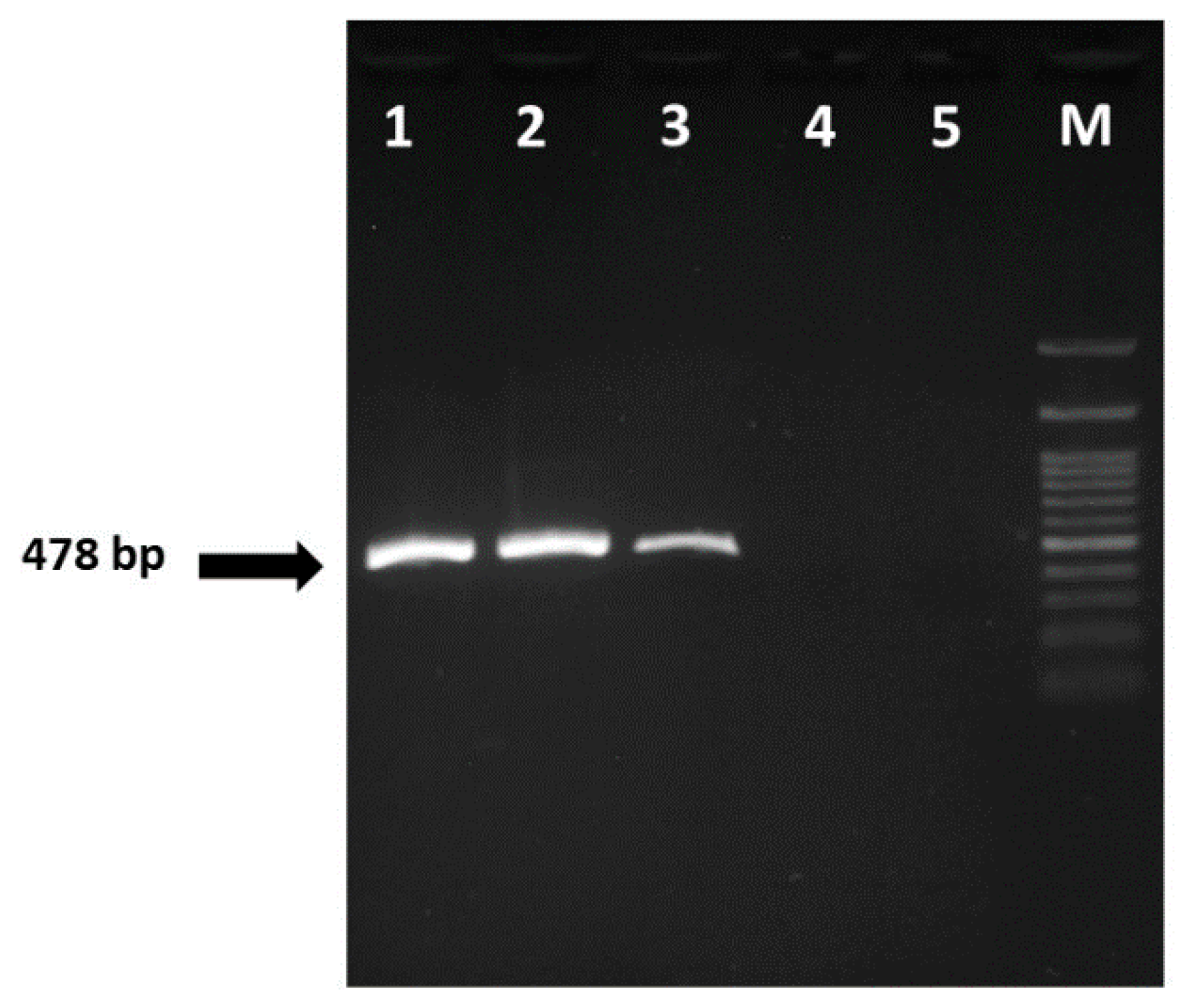
3.3. CEV Confirmation by PCR
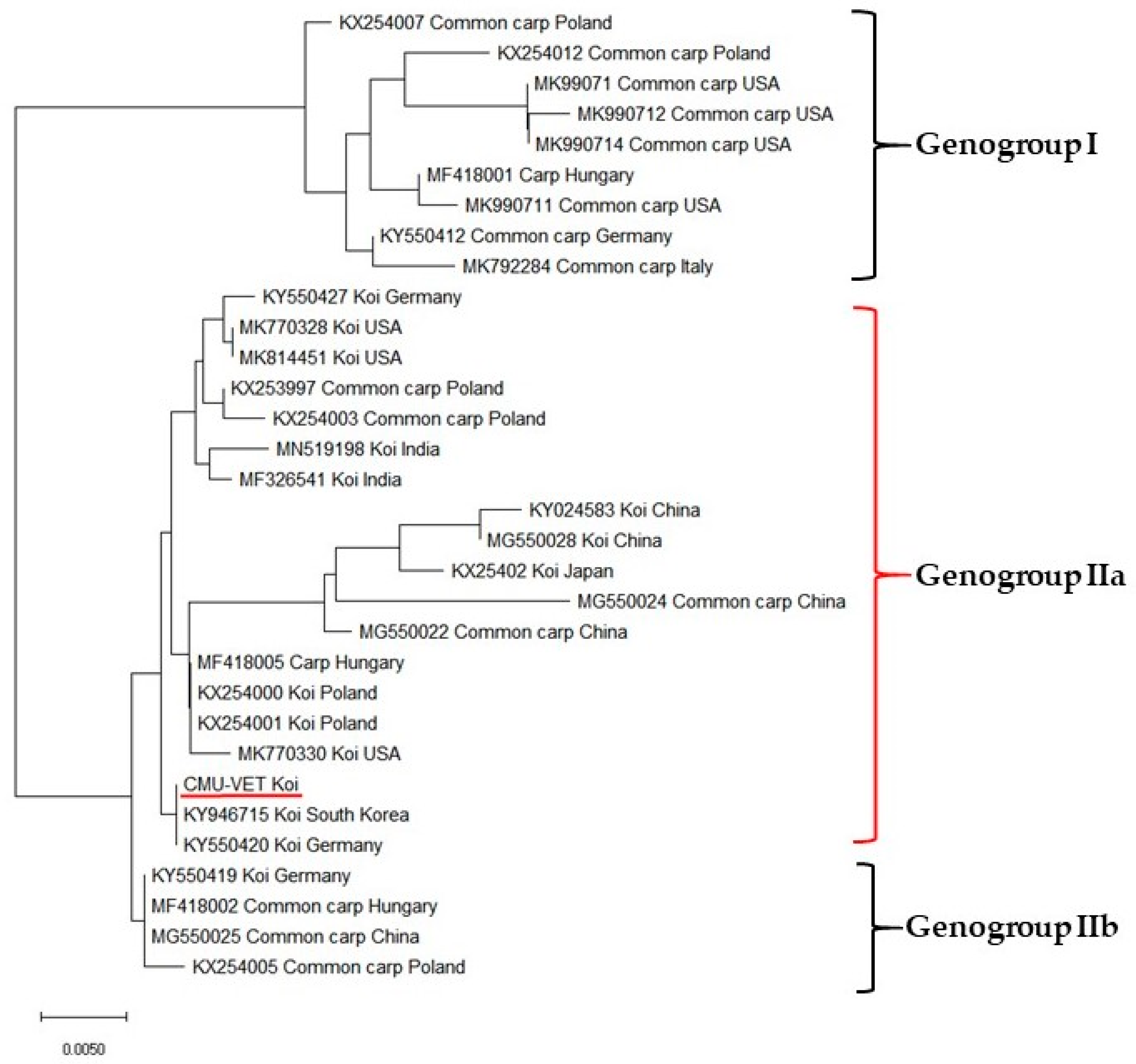
3.4. Sequence Analysis and Phylogenetic Study
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2018—Meeting the Sustainable Development Goals; FAO: Rome, Italy, 2018; Licence: CC BY-NC-SA 3.0 IGO. [Google Scholar]
- David, L.; Rothbard, S.; Rubinstein, I.; Katzman, H.; Hulata, G.; Hillel, J.; Lavi, U. Aspects of red and black color inheritance in the Japanese ornamental (Koi) carp (Cyprinus carpio L.). Aquaculture 2004, 233, 129–147. [Google Scholar] [CrossRef]
- Su, H.; Su, J. Cyprinid viral diseases and vaccine development. Fish Shellfish Immunol. 2018, 83, 84–95. [Google Scholar] [CrossRef]
- Jung, S.J.; Miyazaki, T. Herpesviral haematopoietic necrosis of goldfish, Carassius auratus (L.). J. Fish Dis. 1995, 18, 211–220. [Google Scholar] [CrossRef]
- Bercovier, H.; Fishman, Y.; Nahary, R.; Sinai, S.; Zlotkin, A.; Eyngor, M.; Gilad, O.; Eldar, A.; Hedrick, R.P. Cloning of the koi herpesvirus (KHV) gene encoding thymidine kinase and its use for a highly sensitive PCR based diagnosis. BMC Microbiol. 2005, 5, 13. [Google Scholar] [CrossRef] [PubMed]
- Gilad, O.; Yun, S.; Andree, K.B.; Adkison, M.A.; Zlotkin, A.; Bercovier, H.; Eldar, A.; Hedrick, R.P. Initial characteristics of koi herpesvirus and development of polymerase chain reaction assay to detect the virus in koi, Cyprinus carpio koi. Dis. Aquat. Organ. 2002, 48, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Pikulkaew, S.; Meeyam, T.; Banlunara, W. The outbreak of Koi herpesvirus (KHV) in Koi (Cyprinus carpio koi) from Chiang Mai Province, Thailand. Thai J. Vet. Med. 2009, 39, 53–58. [Google Scholar]
- Office International des Epizooties (OIE). Chapter 2.3.9. Spring viraemia of carp. In Manual of Diagnostic Tests for Aquatic Animals; OIE Press: Rome, Italy, 2019; Available online: https://www.oie.int/index.php?id=2439&L=0&htmfile=chapitre_svc.htm (accessed on 1 October 2020).
- Ahne, W.; Björklund, H.V.; Essbauer, S.; Fijan, N.; Kurath, G.; Winton, J.R. Spring viremia of carp (SVC). Dis. Aquat. Organ. 2002, 52, 261–272. [Google Scholar] [CrossRef]
- Murakami, Y.; Shitanaka, M.; Toshida, S.; Matsuzato, T. Studies on mass mortality of juvenile carp: About mass mortality showing edema. Bull. Hiroshima. Fresh. Water. Fish. Exp. Stn. 1976, 19–33. [Google Scholar]
- Adamek, M.; Jung-Schroers, V.; Hellmann, J.; Teitge, F.; Bergmann, S.M.; Runge, M.; Kleingeld, D.W.; Way, K.; Stone, D.M.; Steinhagen, D. Concentration of carp edema virus (CEV) DNA in koi tissues affected by koi sleepy disease (KSD). Dis. Aquat. Organ. 2016, 119, 245–251. [Google Scholar] [CrossRef]
- Matras, M.; Borzym, E.; Stone, D.; Way, K.; Stachnik, M.; Maj-Paluch, J.; Palusińska, M.; Reichert, M. Carp edema virus in Polish aquaculture—Evidence of significant sequence divergence and a new lineage in common carp Cyprinus carpio (L.). J. Fish Dis. 2017, 40, 319–325. [Google Scholar] [CrossRef]
- Lewisch, E.; Gorgoglione, B.; Way, K.; El-Matbouli, M. Carp edema virus/Koi sleepy disease: An emerging disease in Central-East Europe. Transbound. Emerg. Dis. 2015, 62, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Jung-Schroers, V.; Adamek, M.; Teitge, F.; Hellmann, J.; Bergmann, S.M.; Schütze, H.; Kleingeld, D.W.; Way, K.; Stone, D.; Runge, M.; et al. Another potential carp killer?: Carp Edema Virus disease in Germany. BMC Vet. Res. 2015, 11, 114. [Google Scholar] [CrossRef] [PubMed]
- Way, K.; Haenen, O.; Stone, D.; Adamek, M.; Bergmann, S.M.; Bigarré, L.; Diserens, N.; El-Matbouli, M.; Gjessing, M.C.; Jung-Schroers, V.; et al. Emergence of carp edema virus (CEV) and its significance to European common carp and koi Cyprinus carpio. Dis. Aquat. Organ. 2017, 126, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Adamek, M.; Baska, F.; Vincze, B.; Steinhagen, D. Carp edema virus from three genogroups is present in common carp in Hungary. J. Fish Dis. 2018, 41, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Lovy, J.; Friend, S.E.; Al-Hussinee, L.; Waltzek, T.B. First report of carp edema virus in the mortality of wild common carp Cyprinus carpio in North America. Dis. Aquat. Organ. 2018, 131, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Stevens, B.N.; Michel, A.; Liepnieks, M.L.; Kenelty, K.; Gardhouse, S.M.; Groff, J.M.; Waltzek, T.B.; Soto, E. Outbreak and treatment of carp edema virus in koi (Cyprinus carpio) from northern California. J. Zoo Wildl. Med. 2018, 49, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, T.R.; Kumar, R.; Dharmaratnam, A.; Basheer, V.S.; Sood, N.; Pradhan, P.K.; Sanil, N.K.; Vijayagopal, P.; Jena, J.K. Emergence of carp edema virus in cultured ornamental koi carp, Cyprinus carpio koi, in India. J. Gen. Virol. 2016, 97, 3392–3399. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Giri, S.S.; Kim, S.G.; Kwon, J.; Oh, W.T.; Park, S.C. Carp Edema Virus and Cyprinid Herpesvirus-3 Coinfection is Associated with Mass Mortality of Koi (Cyprinus carpio haematopterus) in the Republic of Korea. Pathogens 2020, 9, 222. [Google Scholar] [CrossRef]
- Luo, F.; Lian, Z.; Niu, Y.; Lü, A.; Hu, X.; Xie, X.; Sun, J. Molecular characterization of carp edema virus disease: An emerging threat to koi Cyprinus carpio in China. Microb. Pathog. 2020, 149, 104551. [Google Scholar] [CrossRef]
- Matějíčková, K.; Pojezdal, Ľ.; Pokorová, D.; Reschová, S.; Piačková, V.; Palíková, M.; Veselý, T.; Papežíková, I. Carp oedema virus disease outbreaks in Czech and Slovak aquaculture. J. Fish Dis. 2020, 43, 971–978. [Google Scholar] [CrossRef]
- Miyazaki, T.; Isshiki, T.; Katsuyuki, H. Histopathological and electron microscopy studies on sleepy disease of koi Cyprinus carpio koi in Japan. Dis. Aquat. Organ. 2005, 65, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Padhi, S.K.; Tolo, I.; McEachran, M.; Primus, A.; Mor, S.K.; Phelps, N.B.D. Koi herpesvirus and carp oedema virus: Infections and coinfections during mortality events of wild common carp in the United States. J. Fish Dis. 2019, 42, 1609–1621. [Google Scholar] [CrossRef] [PubMed]
- Adamek, M.; Matras, M.; Jung-Schroers, V.; Teitge, F.; Heling, M.; Bergmann, S.M.; Reichert, M.; Way, K.; Stone, D.M.; Steinhagen, D. Comparison of PCR methods for the detection of genetic variants of carp edema virus. Dis. Aquat. Organ. 2017, 126, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Soliman, H.; El-Matbouli, M. Rapid detection and differentiation of carp oedema virus and cyprinid herpes virus-3 in koi and common carp. J. Fish Dis. 2018, 41, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Grasshoff, K.; Kremling, K.; Ehrhardt, M. Methods of Seawater Analysis; Verlag Chemie GmbH: Weinheim, Germany, 1983. [Google Scholar]
- Emerson, K.; Russo, R.C.; Lund, R.E.; Thurston, R.V. Aqueous ammonia equilibrium calculations: Effect of pH and temperature. J. Fish. Res. Board. Can. 1975, 32, 2379–2383. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Hwang, J.Y.; Markkandan, K.; Han, K.; Kwon, M.G.; Seo, J.S.; Yoo, S.I.; Hwang, S.D.; Ji, B.Y.; Son, M.H.; Park, J.H. Temperature-dependent immune response of olive flounder (Paralichthys olivaceus) infected with viral hemorrhagic septicemia virus (VHSV). Genes Genom. 2018, 40, 315–320. [Google Scholar] [CrossRef]
- Adamek, M.; Teitge, F.; Steinhagen, D. Quantitative diagnostics of gill diseases in common carp: Not as simple as it seems. Dis. Aquat. Organ. 2019, 134, 197–207. [Google Scholar] [CrossRef]
- Adamek, M.; Teitge, F.; Jung-Schroers, V.; Heling, M.; Gela, D.; Piackova, V.; Kocour, M.; Steinhagen, D. Flavobacteria as secondary pathogens in carp suffering from koi sleepy disease. J. Fish Dis. 2018, 41, 1631–1642. [Google Scholar] [CrossRef]
- Haenen, O.; Way, K.; Gorgoglione, B.; Ito, T.; Paley, R.; Bigarré, L.; Waltzek, T. Novel viral infections threatening cyprinid fish. Bull. Eur. Assoc. Fish. Pathol. 2016, 36, 11–23. [Google Scholar]
- Reite, O.B.; Evensen, O. Inflammatory cells of teleostean fish: A review focusing on mast cells/eosinophilic granule cells and rodlet cells. Fish. Shellfish Immunol. 2006, 20, 192–208. [Google Scholar] [CrossRef]
- Ouyang, P.; Yang, R.; Chen, J.; Wang, K.; Geng, Y.; Lai, W.; Huang, X.; Chen, D.; Fang, J.; Chen, Z.; et al. First detection of carp edema virus in association with cyprinid herpesvirus 3 in cultured ornamental koi, Cyprinus carpio L., in China. Aquacultutre 2018, 490, 162–168. [Google Scholar] [CrossRef]
- Guh, Y.J.; Lin, C.H.; Hwang, P.P. Osmoregulation in zebrafish: Ion transport mechanisms and functional regulation. EXCLI J. 2015, 14, 627–659. [Google Scholar] [PubMed]
- Way, K.; Stone, D. Emergence of carp edema viruslike (CEV-like) disease in the UK. CEFAS Finfish News 2013, 15, 32–34. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Primer Sequence (5′-3′) | Primer Length (bp) | Reference |
|---|---|---|---|
| KHV-TK-F | GGG TTA CCT GTA CGA G | 16 | Bercovier et al. (2005) [5] |
| KHV-TK-R | CAC CCA GTA GAT TAT GC | 17 | |
| KHV9/5F | GAC GAC GCC GGA GAC CTT GTG | 21 | Gilad et al. (2002) [6] |
| KHV9/5R | CAC AAG TTC AGT CTG TTC CTC AAC | 24 | |
| CEV-For-B | ATG GAG TAT CCA AAG TAC TTA G | 22 | Matras et al. (2017) [12] |
| CEV-Rev-J | CTC TTC ACT ATT GTG ACT TTG | 21 | |
| CEV-For-B Internal | GTT ATC AAT GAA ATT TGT GTA TTG | 24 | |
| CEV-Rev-J Internal | TAG CAA AGT ACT ACC TCA TCC | 21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pikulkaew, S.; Phatwan, K.; Banlunara, W.; Intanon, M.; Bernard, J.K. First Evidence of Carp Edema Virus Infection of Koi Cyprinus carpio in Chiang Mai Province, Thailand. Viruses 2020, 12, 1400. https://doi.org/10.3390/v12121400
Pikulkaew S, Phatwan K, Banlunara W, Intanon M, Bernard JK. First Evidence of Carp Edema Virus Infection of Koi Cyprinus carpio in Chiang Mai Province, Thailand. Viruses. 2020; 12(12):1400. https://doi.org/10.3390/v12121400
Chicago/Turabian StylePikulkaew, Surachai, Khathawat Phatwan, Wijit Banlunara, Montira Intanon, and John K. Bernard. 2020. "First Evidence of Carp Edema Virus Infection of Koi Cyprinus carpio in Chiang Mai Province, Thailand" Viruses 12, no. 12: 1400. https://doi.org/10.3390/v12121400
APA StylePikulkaew, S., Phatwan, K., Banlunara, W., Intanon, M., & Bernard, J. K. (2020). First Evidence of Carp Edema Virus Infection of Koi Cyprinus carpio in Chiang Mai Province, Thailand. Viruses, 12(12), 1400. https://doi.org/10.3390/v12121400