
Identification of Novel Feline Paramyxoviruses in Guignas (Leopardus guigna) from Chile

 , , , , ,
, , , , ,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
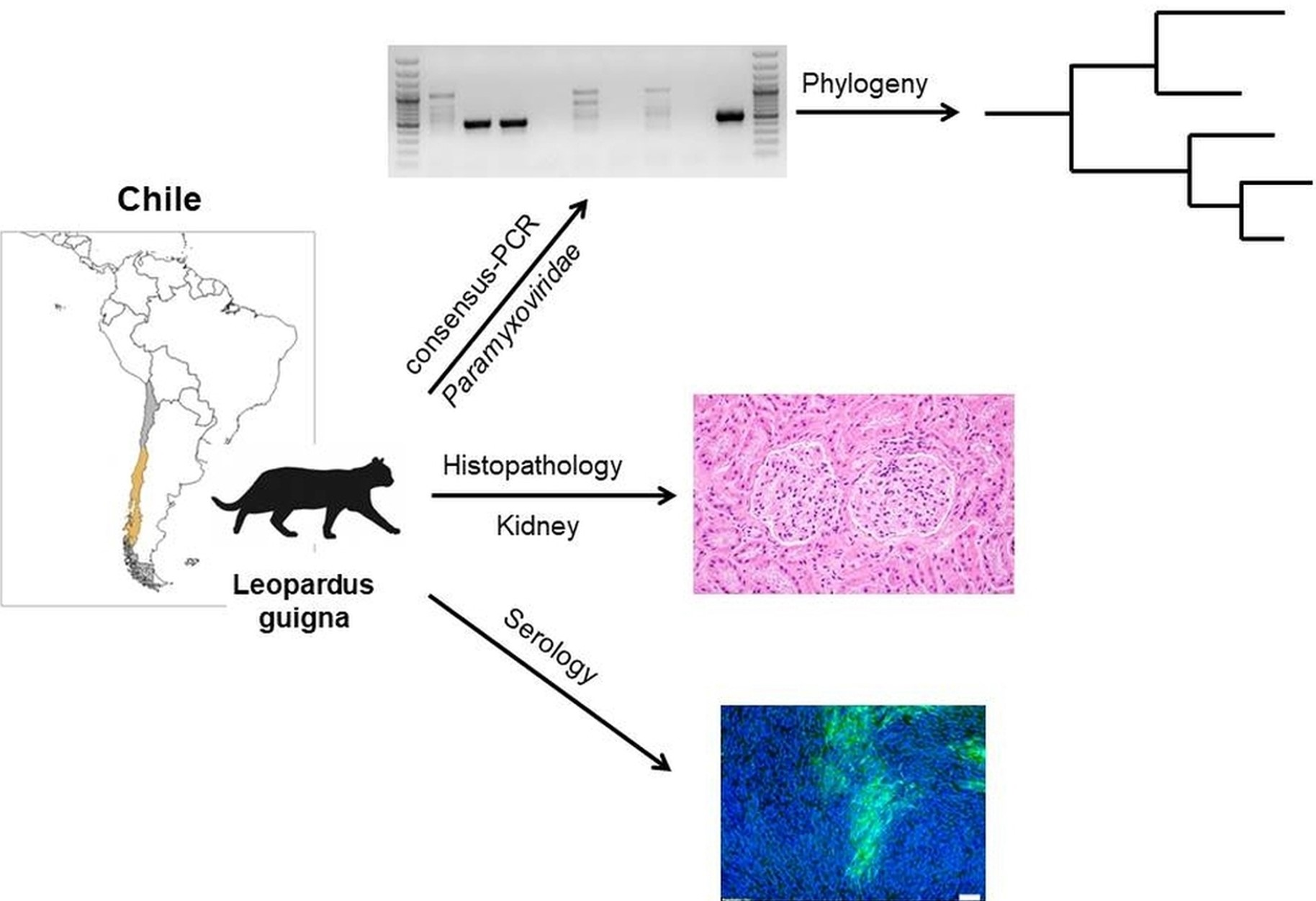
2.1. Sample Collection, RNA-Isolation, PCR Amplification and Histopathology
2.2. Phylogenetic Analysis
2.3. Serological Analysis
2.4. Statistical Analysis
2.5. Virus Isolation Attempts
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Johnson, W.E.; Eizirik, E.; Pecon-Slattery, J.; Murphy, W.J.; Antunes, A.; Teeling, E.; O’Brien, S.J. The Late Miocene Radiation of Modern Felidae: A Genetic Assessment. Science 2006, 311, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Nowell, K.; Jackson, P. Wild Cats: Status Survey and Conservation Action Plan; International Union for Conservation of Nature: Gland, Switzerland, 1996; ISBN 2-8317-0045-0. [Google Scholar]
- Johnson, W.E.; Slattery, J.P.; Eizirik, E.; Kim, J.-H.; Raymond, M.M.; Bonacic, C.; Cambre, R.; Crawshaw, P.; Nunes, A.; Seuanez, H.N.; et al. Disparate phylogeographic patterns of molecular genetic variation in four closely related South American small cat species. Mol. Ecol. 1999, 8, S79–S94. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, C.; Johnson, W.E.; Sanderson, J.; O’Brien, S.J.; Hoelzel, A.R.; Freer, R.; Dunstone, N.; Ritland, K.; Ritland, C.E.; Poulin, E. Phylogeography and population history of Leopardus guigna, the smallest American felid. Conserv. Genet. 2014, 15, 631–653. [Google Scholar] [CrossRef]
- Napolitano, C.; Gálvez, N.; Bennett, M.; Acosta-Jamett, G.; Sanderson, J. Leopardus guigna. IUCN Red List Threat. Species 2015 2015. [Google Scholar] [CrossRef]
- Mora, M.; Napolitano, C.; Ortega, R.; Poulin, E.; Pizarro-Lucero, J. Feline immunodeficiency virus and feline leukemia virus infection in free-ranging guignas (Leopardus guigna) and sympatric domestic cats in human perturbed landscapes on Chiloé Island, Chile. J. Wildl. Dis. 2015, 51, 199–208. [Google Scholar] [CrossRef]
- Ortega, R.; Mena, J.; Grecco, S.; Pérez, R.; Panzera, Y.; Napolitano, C.; Zegpi, N.; Sandoval, A.; Sandoval, D.; González-Acuña, D.; et al. Domestic dog origin of Carnivore Protoparvovirus 1 infection in a rescued free-ranging guiña (Leopardus guigna) in Chile. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- O’Brien, S.; Troyer, J.L.; Brown, M.A.; Johnson, W.E.; Antunes, A.; Roelke-Parker, M.E.; Pecon-Slattery, J. Emerging Viruses in the Felidae: Shifting Paradigms. Viruses 2012, 4, 236–257. [Google Scholar] [CrossRef]
- Roelke, M.; Munson, L.; Packer, C.; Kock, R.; Cleaveland, S.; Carpenter, M.; O’Brien, S.; Pospischil, A.; Hofmann-Lehmann, R.; Lutz, H.; et al. A canine distemper virus epidemic in Serengeti lions (Panthera leo). Nature 1996, 379, 441–445. [Google Scholar] [CrossRef]
- Seimon, T.A.; Miquelle, D.G.; Chang, T.Y.; Newton, A.L.; Korotkova, I.; Ivanchuk, G.; Lyubchenko, E.; Tupikov, A.; Slabe, E.; McAloose, D. Canine Distemper Virus: An Emerging Disease in Wild Endangered Amur Tigers (Panthera tigris altaica). mBio 2013, 4, e00410-13. [Google Scholar] [CrossRef]
- Beineke, A.; Baumgärtner, W.; Wohlsein, P. Cross-species transmission of canine distemper virus—An update. One Health 2015, 1, 49–59. [Google Scholar] [CrossRef]
- Terio, K.A.; Craft, M.E. Canine Distemper Virus (CDV) in Another Big Cat: Should CDV Be Renamed Carnivore Distemper Virus? mBio 2013, 4, e00702-13. [Google Scholar] [CrossRef] [PubMed]
- Knipe, D.M.; Howley, P.M. (Eds.) Fields Virology, 6th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; ISBN 9781451105636. [Google Scholar]
- Woo, P.C.Y.; Lau, S.K.P.; Wong, B.H.L.; Fan, R.Y.Y.; Wong, A.Y.P.; Zhang, A.J.X.; Wu, Y.; Choi, G.K.Y.; Li, K.S.M.; Hui, J.; et al. Feline morbillivirus, a previously undescribed paramyxovirus associated with tubulointerstitial nephritis in domestic cats. Proc. Natl. Acad. Sci. USA 2012, 109, 5435–5440. [Google Scholar] [CrossRef] [PubMed]
- Sieg, M.; Heenemann, K.; Rückner, A.; A Burgener, I.; Oechtering, G.; Vahlenkamp, T.W. Discovery of new feline paramyxoviruses in domestic cats with chronic kidney disease. Virus Genes 2015, 51, 294–297. [Google Scholar] [CrossRef] [PubMed]
- Lorusso, A.; Di Tommaso, M.; Di Felice, E.; Zaccaria, G.; Luciani, A.; Marcacci, M.; Aste, G.; Boari, A.; Savini, G. First report of feline morbillivirus in Europe. Vet. Ital. 2015, 51, 235–237. [Google Scholar] [PubMed]
- McCallum, K.E.; Stubbs, S.; Hope, N.; Mickleburgh, I.; Dight, D.; Tiley, L.; Williams, T.L. Detection and seroprevalence of morbillivirus and other paramyxoviruses in geriatric cats with and without evidence of azotemic chronic kidney disease. J. Vet. Intern. Med. 2018, 32, 1100–1108. [Google Scholar] [CrossRef]
- Furuya, T.; Sassa, Y.; Omatsu, T.; Nagai, M.; Fukushima, R.; Shibutani, M.; Yamaguchi, T.; Uematsu, Y.; Shirota, K.; Mizutani, T. Existence of feline morbillivirus infection in Japanese cat populations. Arch. Virol. 2014, 159, 371–373. [Google Scholar] [CrossRef]
- Yilmaz, H.; Tekelioglu, B.K.; Gurel, A.; Bamac, O.E.; Ozturk, G.Y.; Cizmecigil, U.Y.; Altan, E.; Aydin, O.; Yilmaz, A.; Berriatua, E.; et al. Frequency, clinicopathological features and phylogenetic analysis of feline morbillivirus in cats in Istanbul, Turkey. J. Feline Med. Surg. 2017, 19, 1206–1214. [Google Scholar] [CrossRef]
- Isa, N.H.M.; Selvarajah, G.T.; Khor, K.H.; Tan, S.W.; Manoraj, H.; Omar, N.H.; Omar, A.R.; Kamal, F.M. Molecular detection and characterisation of feline morbillivirus in domestic cats in Malaysia. Vet. Microbiol. 2019, 236, 108382. [Google Scholar] [CrossRef]
- Chaiyasak, S.; Piewbang, C.; Rungsipipat, A.; Techangamsuwan, S. Molecular epidemiology and genome analysis of feline morbillivirus in household and shelter cats in Thailand. BMC Vet. Res. 2020, 16, 1–9. [Google Scholar] [CrossRef]
- Sharp, C.R.; Nambulli, S.; Acciardo, A.S.; Rennick, L.J.; Drexler, J.F.; Rima, B.K.; Williams, T.; Duprex, W.P. Chronic Infection of Domestic Cats with Feline Morbillivirus, United States. Emerg. Infect. Dis. 2016, 22, 760–762. [Google Scholar] [CrossRef]
- Darold, G.M.; Alfieri, A.A.; Muraro, L.S.; Amude, A.M.; Zanatta, R.; Yamauchi, K.C.I.; Alfieri, A.F.; Lunardi, M. First report of feline morbillivirus in South America. Arch. Virol. 2017, 162, 469–475. [Google Scholar] [CrossRef]
- Sieg, M.; Busch, J.; Eschke, M.; Böttcher, D.; Heenemann, K.; Vahlenkamp, A.; Reinert, A.; Seeger, J.; Heilmann, R.M.; Scheffler, K.; et al. A New Genotype of Feline Morbillivirus Infects Primary Cells of the Lung, Kidney, Brain and Peripheral Blood. Viruses 2019, 11, 146. [Google Scholar] [CrossRef]
- Sakaguchi, S.; Nakagawa, S.; Mitsuhashi, S.; Ogawa, M.; Sugiyama, K.; Tamukai, K.; Koide, R.; Katayama, Y.; Nakano, T.; Makino, S.; et al. Molecular characterization of feline paramyxovirus in Japanese cat populations. Arch. Virol. 2019, 165, 413–418. [Google Scholar] [CrossRef]
- Tong, S.; Chern, S.-W.W.; Li, Y.; Pallansch, M.A.; Anderson, L.J. Sensitive and Broadly Reactive Reverse Transcription-PCR Assays To Detect Novel Paramyxoviruses. J. Clin. Microbiol. 2008, 46, 2652–2658. [Google Scholar] [CrossRef]
- Sutummaporn, K.; Suzuki, K.; Machida, N.; Mizutani, T.; Park, E.-S.; Morikawa, S.; Furuya, T.; Shigeru, M. Association of feline morbillivirus infection with defined pathological changes in cat kidney tissues. Vet. Microbiol. 2019, 228, 12–19. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Busch, J.; Sacristán, I.; Cevidanes, A.; Millán, J.; Vahlenkamp, T.W.; Napolitano, C.; Sieg, M. High seroprevalence of feline morbilliviruses in free-roaming domestic cats in Chile. Arch. Virol. 2020. [Google Scholar] [CrossRef]
- Sieg, M.; Vahlenkamp, A.; Baums, C.G.; Vahlenkamp, T.W. First Complete Genome Sequence of a Feline Morbillivirus Isolate from Germany. Genome Announc. 2018, 6, e00244-18. [Google Scholar] [CrossRef] [PubMed]
- Litster, A.; Pressler, B.; Volpe, A.; Dubovi, E. Accuracy of a point-of-care ELISA test kit for predicting the presence of protective canine parvovirus and canine distemper virus antibody concentrations in dogs. Vet. J. 2012, 193, 363–366. [Google Scholar] [CrossRef]
- Mortlock, M.; Kuzmin, I.V.; Weyer, J.; Gilbert, A.T.; Agwanda, B.; Rupprecht, C.E.; Nel, L.H.; Kearney, T.; Malekani, J.M.; Markotter, W. Novel Paramyxoviruses in Bats from Sub-Saharan Africa, 2007–2012. Emerg. Infect. Dis. 2015, 21, 1840–1843. [Google Scholar] [CrossRef]
- Drexler, J.F.; Corman, V.M.; Müller, M.A.; Maganga, G.D.; Vallo, P.; Binger, T.; Gloza-Rausch, F.; Cottontail, V.M.; Rasche, A.; Yordanov, S.; et al. Bats host major mammalian paramyxoviruses. Nat. Commun. 2012, 3, 796. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Muleya, W.; Ishii, A.; Orba, Y.; Hang’Ombe, B.M.; Mweene, A.S.; Moonga, L.; Thomas, Y.; Kimura, T.; Sawa, H. Molecular epidemiology of paramyxoviruses in Zambian wild rodents and shrews. J. Gen. Virol. 2014, 95, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Prada, D.; Boyd, V.; Baker, M.L.; O’Dea, M.; Jackson, B. Viral Diversity of Microbats within the South West Botanical Province of Western Australia. Viruses 2019, 11, 1157. [Google Scholar] [CrossRef]
- Kurth, A.; Kohl, C.; Brinkmann, A.; Ebinger, A.; Harper, J.A.; Wang, L.-F.; Mühldorfer, K.; Wibbelt, G. Novel Paramyxoviruses in Free-Ranging European Bats. PLoS ONE 2012, 7, e38688. [Google Scholar] [CrossRef]
- De Luca, E.; Crisi, P.E.; Marcacci, M.; Malatesta, D.; Di Sabatino, D.; Cito, F.; D’Alterio, N.; Puglia, I.; Berjaoui, S.; Colaianni, M.L.; et al. Epidemiology, pathological aspects and genome heterogeneity of feline morbillivirus in Italy. Vet. Microbiol. 2019, 240, 108484. [Google Scholar] [CrossRef]
- Piewbang, C.; Chaiyasak, S.; Kongmakee, P.; Sanannu, S.; Khotapat, P.; Ratthanophart, J.; Banlunara, W.; Techangamsuwan, S. Feline Morbillivirus Infection Associated with Tubulointerstitial Nephritis in Black Leopards (Panthera pardus). Vet. Pathol. 2020, 57, 871–879. [Google Scholar] [CrossRef]
- Sacristán, I.; Esperon, F.; Pérez, R.; Acuña, F.; Aguilar, E.; Garcia, S.; López, M.J.; Neves, E.; Cabello, J.; Hidalgo-Hermoso, E.; et al. Epidemiology, molecular characterization and risk factors of Carnivore protoparvovirus-1 infection and disease in the wild felid Leopardus guigna in Chile. Authorea Prepr. 2020. [Google Scholar] [CrossRef]
- Gilardi, K.V.K.; Mazet, J.A.K. The United States Agency for International Development Emerging Pandemic Threats PREDICT Project—Global Detection of Emerging Wildlife Viral Zoonoses. In Fowler’s Zoo and Wild Animal Medicine Current Therapy; Saunders: Philadelphia, PA, USA, 2019; pp. 110–116. [Google Scholar] [CrossRef]
- Troyer, J.L.; Roelke, M.E.; Jespersen, J.M.; Baggett, N.; Buckley-Beason, V.; MacNulty, D.; Craft, M.; Packer, C.; Pecon-Slattery, J.; O’Brien, S.J. FIV diversity: FIVPle subtype composition may influence disease outcome in African lions. Vet. Immunol. Immunopathol. 2011, 143, 338–346. [Google Scholar] [CrossRef]
- Carpenter, M.A.; Brown, E.W.; Culver, M.; Johnson, W.E.; Pecon-Slattery, J.; Brousset, D.; O’Brien, S.J. Genetic and phylogenetic divergence of feline immunodeficiency virus in the puma (Puma concolor). J. Virol. 1996, 70, 6682–6693. [Google Scholar] [CrossRef]
- Nikolin, V.M.; Olarte-Castillo, X.A.; Osterrieder, N.; Hofer, H.; Dubovi, E.; Mazzoni, C.J.; Brunner, E.; Goller, K.V.; Fyumagwa, R.D.; Moehlman, P.D.; et al. Canine distemper virus in the Serengeti ecosystem: Molecular adaptation to different carnivore species. Mol. Ecol. 2017, 26, 2111–2130. [Google Scholar] [CrossRef]
- Sacristán, I.; Sieg, M.; Acuña, F.; Aguilar, E.; García, S.; López, M.J.; Cevidanes, A.; Hidalgo-Hermoso, E.; Cabello, J.; Vahlenkamp, T.W.; et al. Molecular and serological survey of carnivore pathogens in free-roaming domestic cats of rural communities in southern Chile. J. Vet. Med. Sci. 2019, 81, 1740–1748. [Google Scholar] [CrossRef]
- Arikawa, K.; Wachi, A.; Imura, Y.; Sutummaporn, K.; Kai, C.; Park, E.-S.; Morikawa, S.; Uematsu, Y.; Yamaguchi, T.; Furuya, T. Development of an ELISA for serological detection of feline morbillivirus infection. Arch. Virol. 2017, 162, 2421–2425. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Identity 1 | Animals 1 (Accession Number) | Subspecies 1 | Origin | Highest Nucleotide Homology to (Accession No.) | Relatedness to Other Paramyxoviruses |
|---|---|---|---|---|---|
| FPaV- related | LG 126 (MN850340) | LGG | Chiloe Island | 83% FPaV Felis catus, Japan (LC431581.1) | 76% bat paramyxovirus (KC578587.1) 74% Tenrec paramyxovirus (KF246040.1) 73% rodent paramyxovirus (JF828299.1) |
| LG 127 (MN850341) | LGG | ||||
| LG 184 (MN850343) | LGG | ||||
| LG 182 (MN850342) | LGT | Chilean continent | 82% FPaV (LC431581.1) | ||
| FeMV- related | LG 128 (MN850344) | LGG | Chiloe Island | 75% FeMV Felis catus, Brazil (KX452077.1) | 70% bat paramyxovirus (MN602070.1) |
| LG 137 (MN850346) | LGG | ||||
| LG 136 (MN850345) | LGT | Chilean continent | 74% FeMV Felis catus, China (JQ411016.1) | 70% rodent paramyxovirus (AB844347.1) 70% bat paramyxovirus (MH259211.1) | |
| LG 181 (MN850347) | LGG | ||||
| LG 186 (MN850348) | LGG | ||||
| LG 189 (MN850349) | LGG | ||||
| LG 193 (MN850350) | LGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sieg, M.; Sacristán, I.; Busch, J.; Terio, K.A.; Cabello, J.; Hidalgo-Hermoso, E.; Millán, J.; Böttcher, D.; Heenemann, K.; Vahlenkamp, T.W.; et al. Identification of Novel Feline Paramyxoviruses in Guignas (Leopardus guigna) from Chile. Viruses 2020, 12, 1397. https://doi.org/10.3390/v12121397
Sieg M, Sacristán I, Busch J, Terio KA, Cabello J, Hidalgo-Hermoso E, Millán J, Böttcher D, Heenemann K, Vahlenkamp TW, et al. Identification of Novel Feline Paramyxoviruses in Guignas (Leopardus guigna) from Chile. Viruses. 2020; 12(12):1397. https://doi.org/10.3390/v12121397
Chicago/Turabian StyleSieg, Michael, Irene Sacristán, Johannes Busch, Karen A. Terio, Javier Cabello, Ezequiel Hidalgo-Hermoso, Javier Millán, Denny Böttcher, Kristin Heenemann, Thomas W. Vahlenkamp, and et al. 2020. "Identification of Novel Feline Paramyxoviruses in Guignas (Leopardus guigna) from Chile" Viruses 12, no. 12: 1397. https://doi.org/10.3390/v12121397
APA StyleSieg, M., Sacristán, I., Busch, J., Terio, K. A., Cabello, J., Hidalgo-Hermoso, E., Millán, J., Böttcher, D., Heenemann, K., Vahlenkamp, T. W., & Napolitano, C. (2020). Identification of Novel Feline Paramyxoviruses in Guignas (Leopardus guigna) from Chile. Viruses, 12(12), 1397. https://doi.org/10.3390/v12121397