Experimental Infection of Pregnant Female Sheep with Zika Virus During Early Gestation

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Animal Infection and Evaluation of Clinical Disease
2.3. Post-Mortem Evaluation and Tissue Collection
2.4. Virus Quantification
2.5. Virus Isolation from Animal Tissue
2.6. Real-Time PCR
2.7. Serology
2.7.1. IgM Capture ELISA
2.7.2. Plaque Reduction Neutralization Test
2.8. Fluorescence-Activated Cell Sorting
2.9. Immortalized Sheep Cell Growth and Infection
2.10. Immunofluorescence of Immortalized Sheep Cells
2.11. Statistical Analysis
3. Results
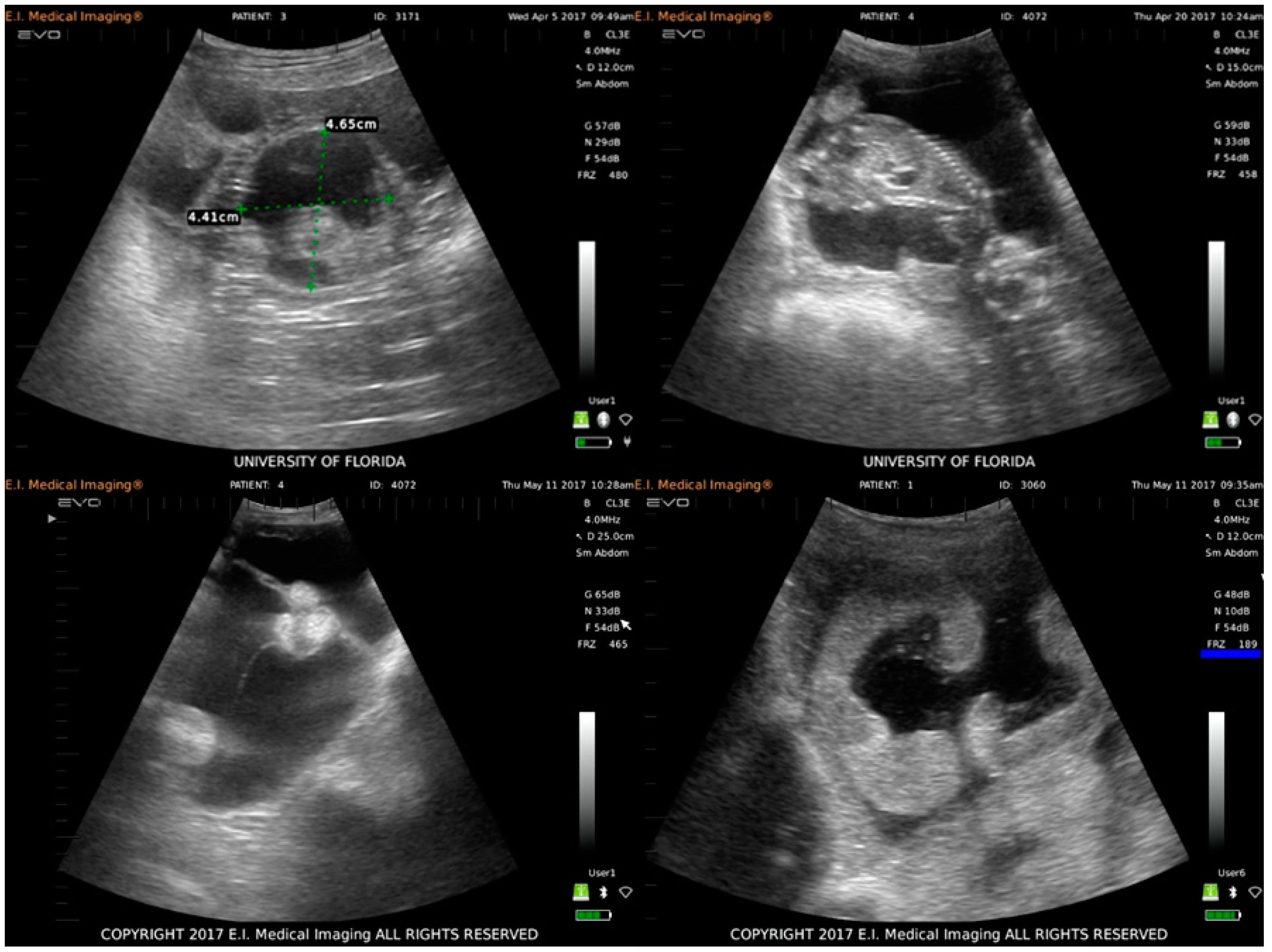
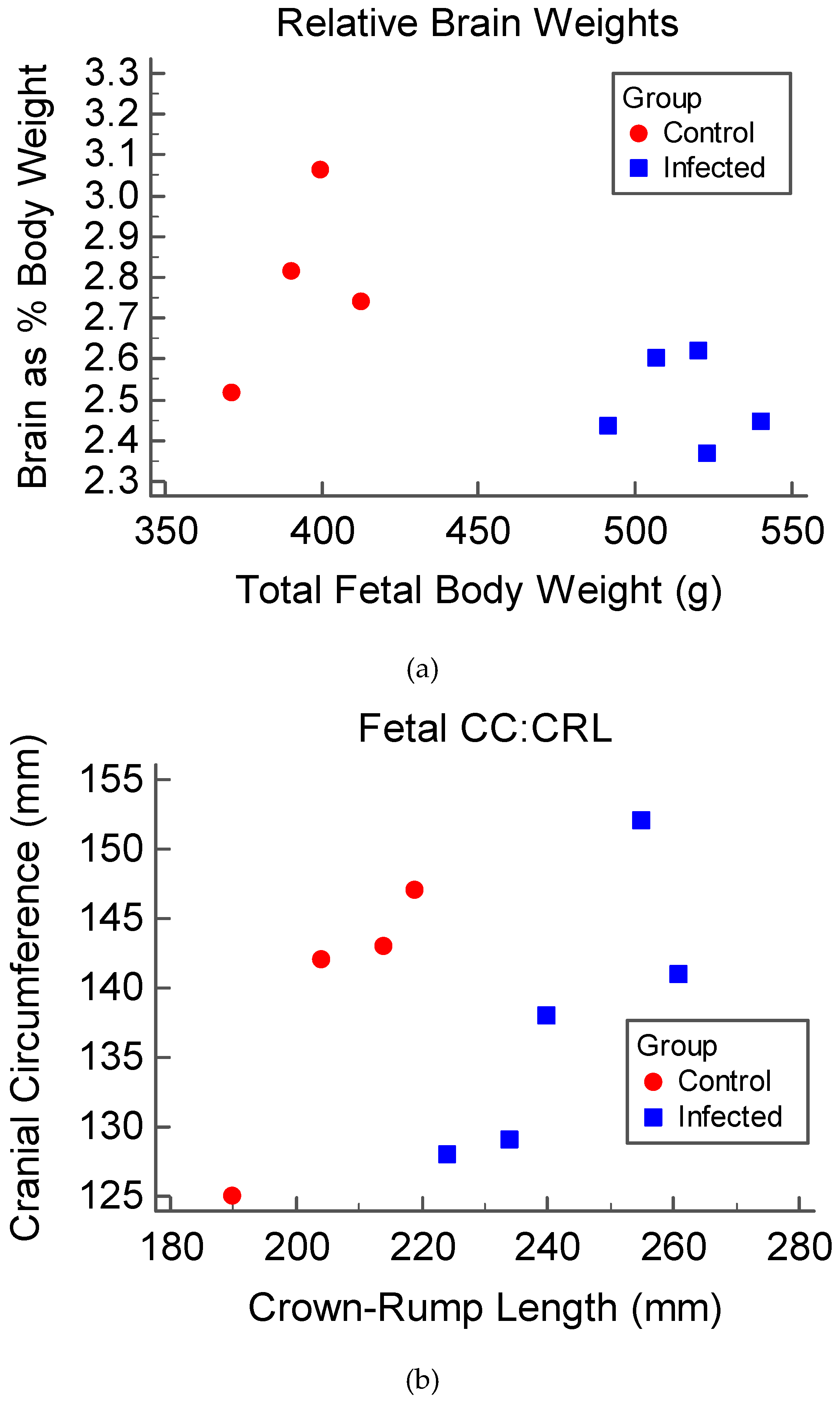
3.1. Clinical Disease and Fetal Development
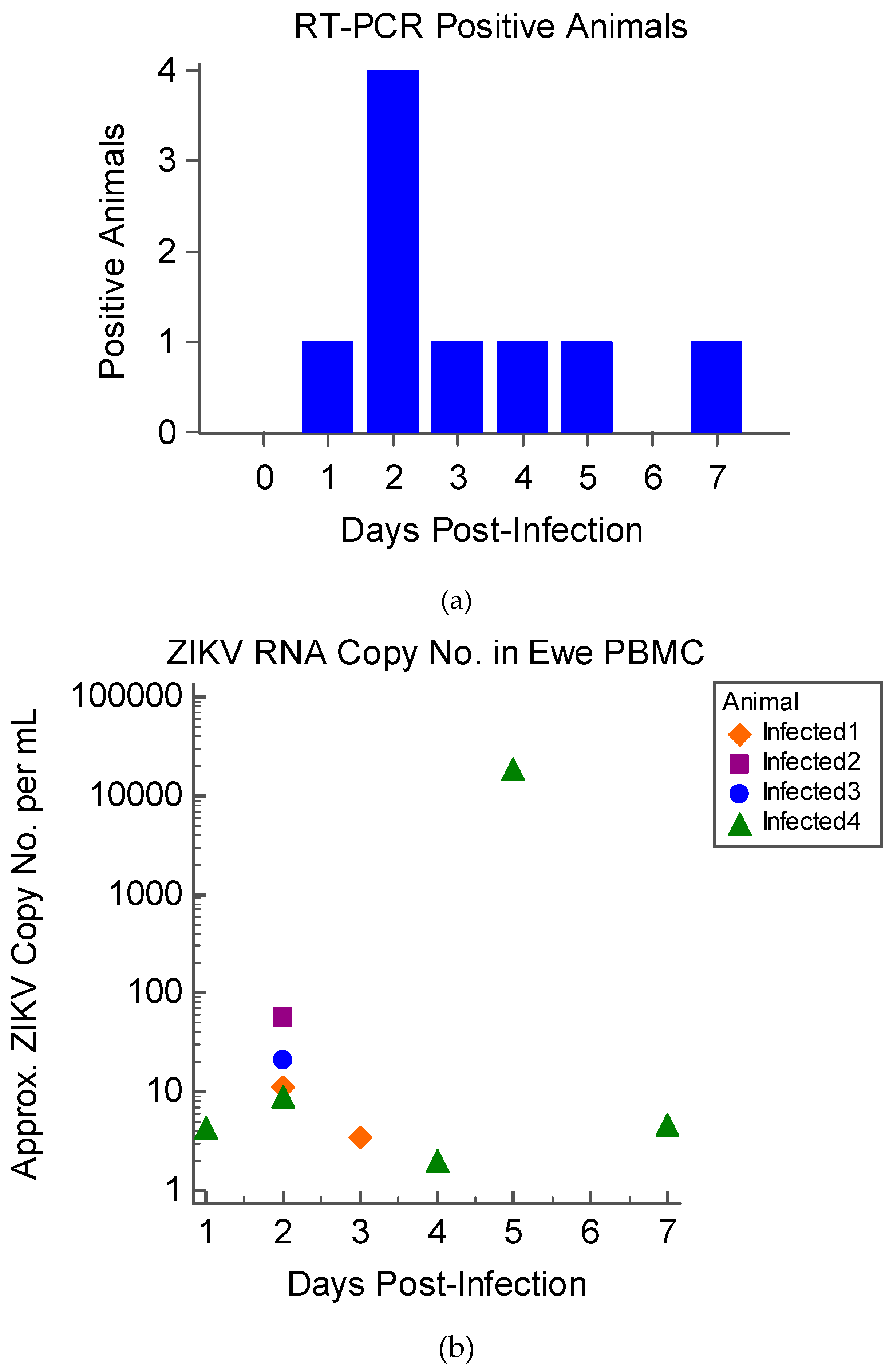
3.2. Virus Isolation and Detection
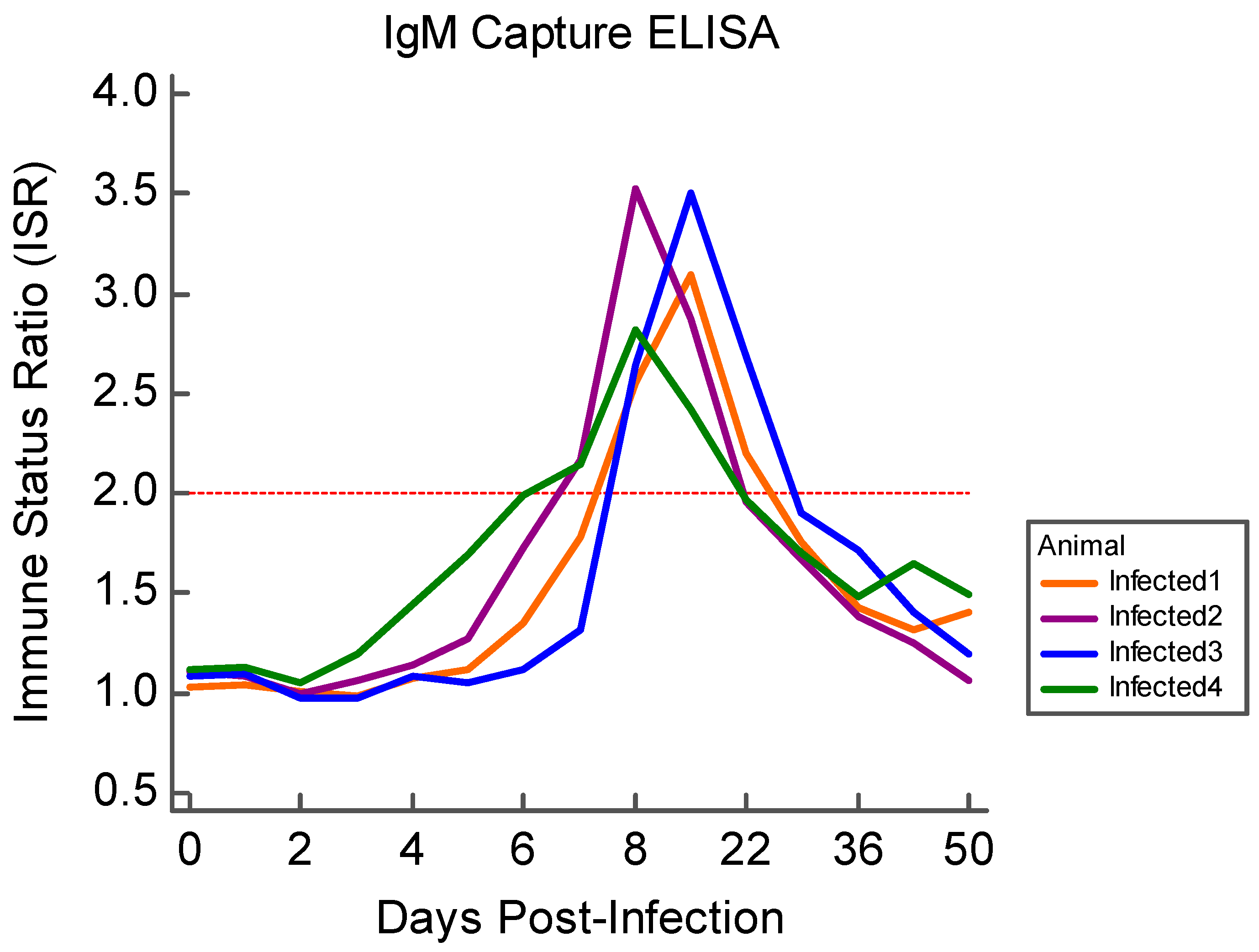
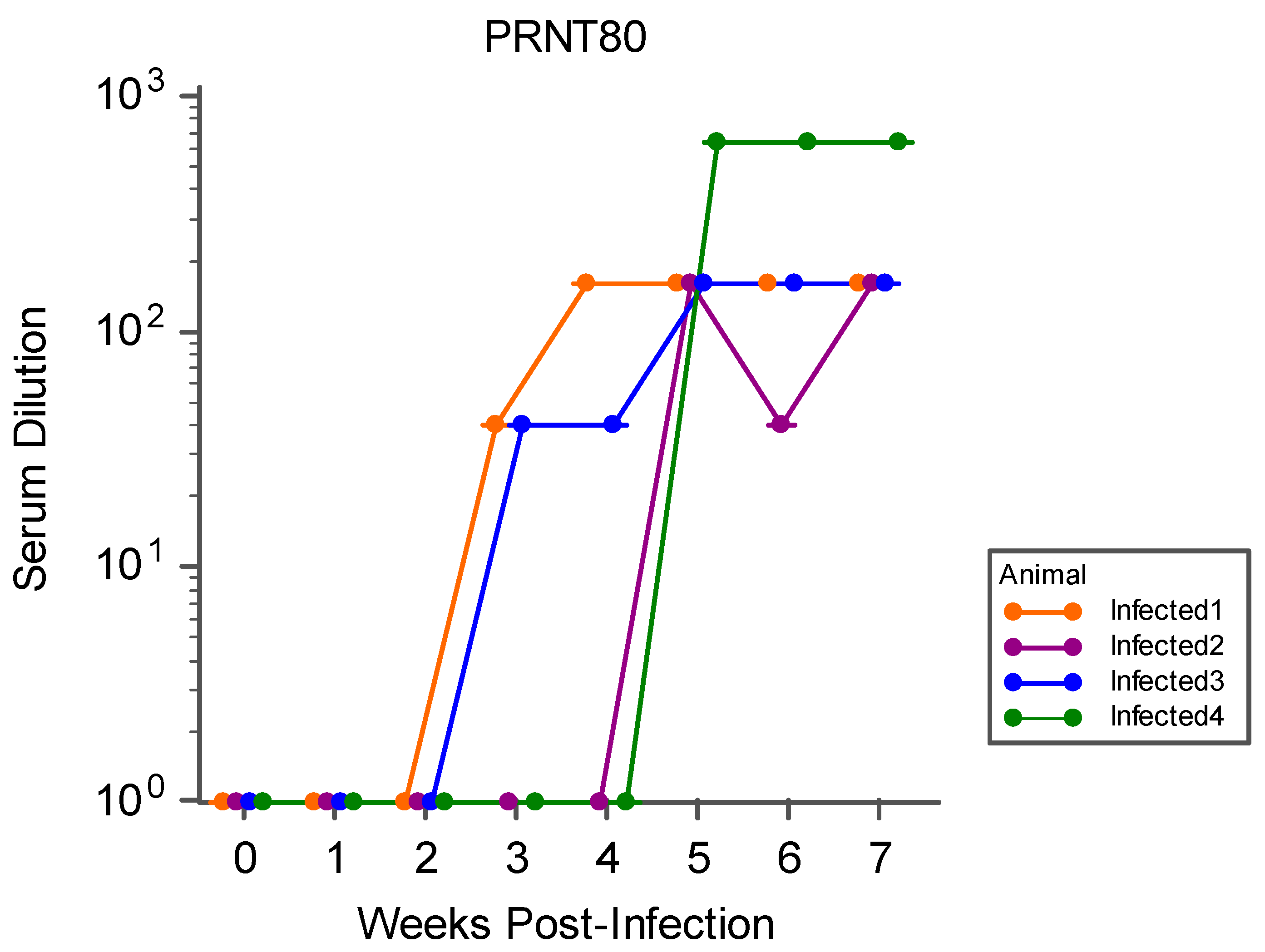
3.3. Serology
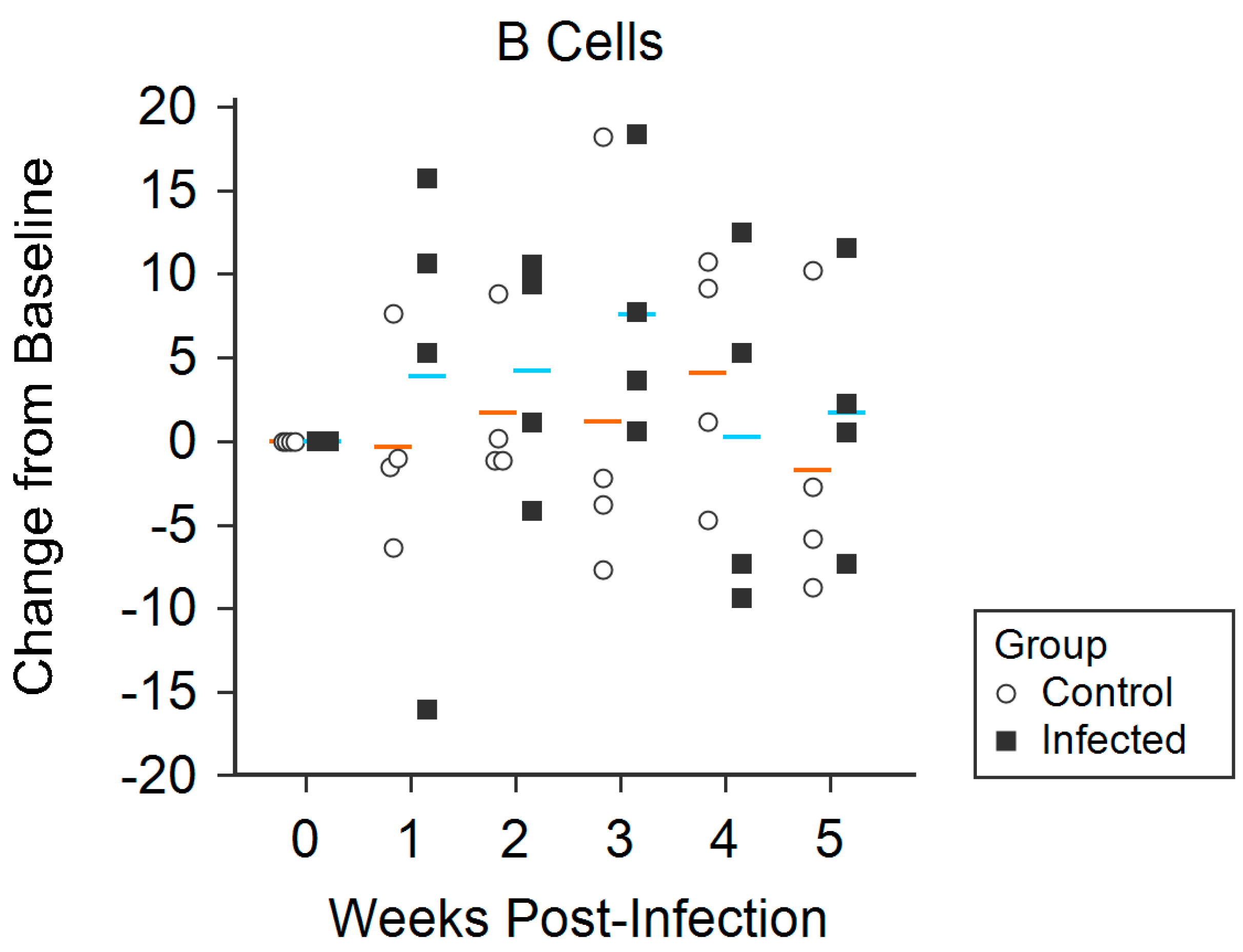
3.4. Flow Cytometry
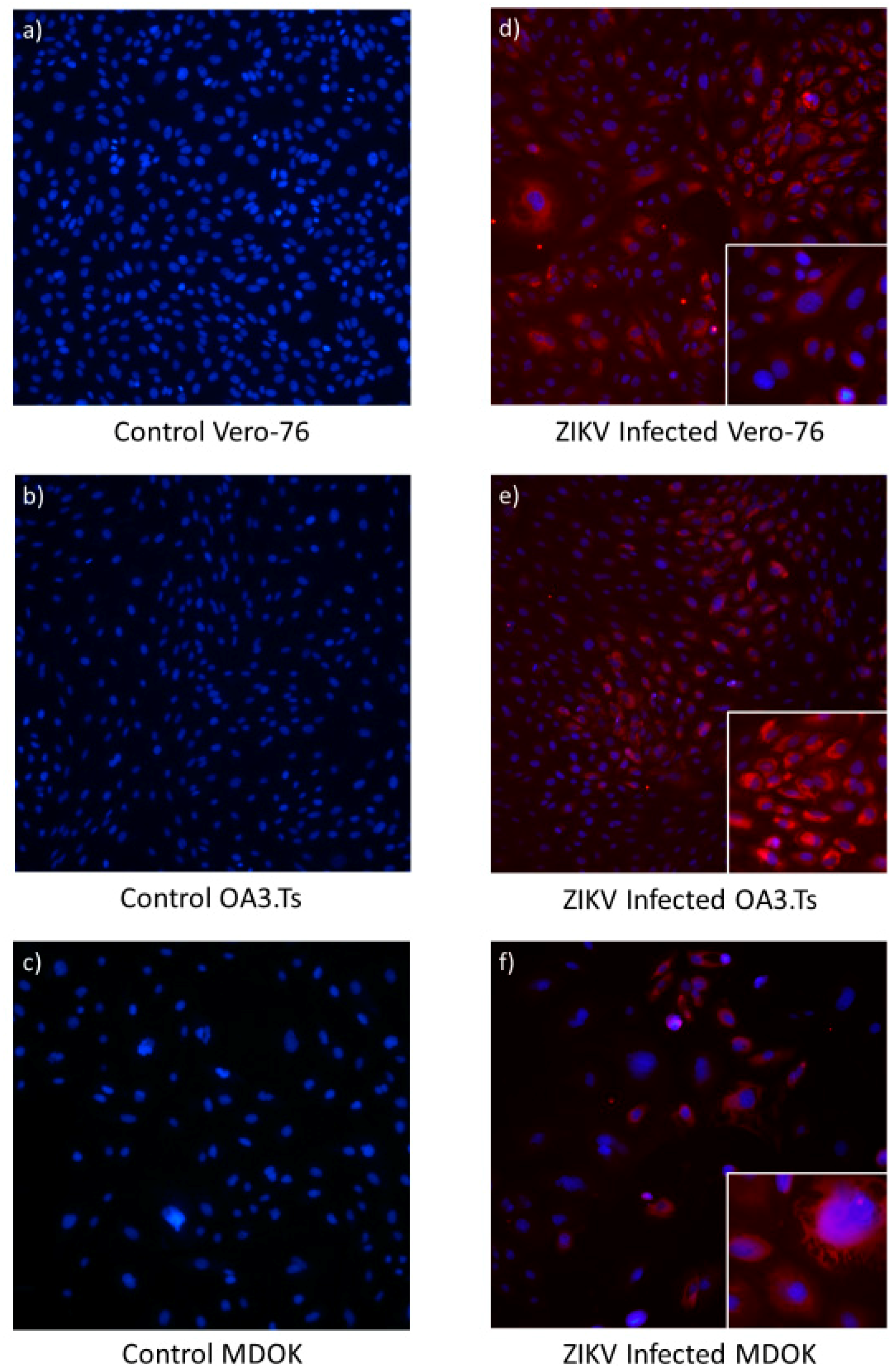
3.5. Immortalized Sheep Cell Permissiveness to ZIKV
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Melo, A.S.; Aguiar, R.S.; Amorim, M.M.; Arruda, M.B.; Melo, F.O.; Ribeiro, S.T.; Batista, A.G.; Ferreira, T.; Dos Santos, M.P.; Sampaio, V.V.; et al. Congenital Zika Virus Infection: Beyond Neonatal Microcephaly. JAMA Neurol. 2016, 73, 1407–1416. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, M.; Feitosa, I.M.; Ribeiro, E.M.; Horovitz, D.D.; Pessoa, A.L.; França, G.V.; García-Alix, A.; Doriqui, M.J.; Wanderley, H.Y.; Sanseverino, M.V.; et al. The phenotypic spectrum of congenital Zika syndrome. Am. J. Med. Genet. A 2017, 173, 841–857. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, W.K.; de França, G.V.A.; Carmo, E.H.; Duncan, B.B.; de Souza Kuchenbecker, R.; Schmidt, M.I. Infection-related microcephaly after the 2015 and 2016 Zika virus outbreaks in Brazil: A surveillance-based analysis. Lancet 2017, 390, 861–870. [Google Scholar] [CrossRef]
- Li, R.; Simmons, K.B.; Bertolli, J.; Rivera-Garcia, B.; Cox, S.; Romero, L.; Koonin, L.M.; Valencia-Prado, M.; Bracero, N.; Jamieson, D.J.; et al. Cost-effectiveness of Increasing Access to Contraception during the Zika Virus Outbreak, Puerto Rico, 2016. Emerg. Infect. Dis. 2017, 23, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Gurung, S.; Preno, A.N.; Dubaut, J.P.; Nadeau, H.; Hyatt, K.; Reuter, N.; Nehete, B.; Wolf, R.F.; Nehete, P.; Dittmer, D.P.; et al. Translational Model of Zika Virus Disease in Baboons. J. Virol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Coffey, L.L.; Keesler, R.I.; Pesavento, P.A.; Woolard, K.; Singapuri, A.; Watanabe, J.; Cruzen, C.; Christe, K.L.; Usachenko, J.; Yee, J.; et al. Intraamniotic Zika virus inoculation of pregnant rhesus macaques produces fetal neurologic disease. Nat. Commun. 2018, 9, 2414. [Google Scholar] [CrossRef] [PubMed]
- Carroll, T.; Lo, M.; Lanteri, M.; Dutra, J.; Zarbock, K.; Silveira, P.; Rourke, T.; Ma, Z.M.; Fritts, L.; O’Connor, S.; et al. Zika virus preferentially replicates in the female reproductive tract after vaginal inoculation of rhesus macaques. PLoS Pathog. 2017, 13, e1006537. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, S.M.; Antony, K.M.; Dudley, D.M.; Kohn, S.; Simmons, H.A.; Wolfe, B.; Salamat, M.S.; Teixeira, L.B.C.; Wiepz, G.J.; Thoong, T.H.; et al. Highly efficient maternal-fetal Zika virus transmission in pregnant rhesus macaques. PLoS Pathog. 2017, 13, e1006378. [Google Scholar] [CrossRef]
- Dudley, D.M.; Aliota, M.T.; Mohr, E.L.; Weiler, A.M.; Lehrer-Brey, G.; Weisgrau, K.L.; Mohns, M.S.; Breitbach, M.E.; Rasheed, M.N.; Newman, C.M.; et al. A rhesus macaque model of Asian-lineage Zika virus infection. Nat. Commun. 2016, 7, 12204. [Google Scholar] [CrossRef]
- Yun, S.I.; Song, B.H.; Frank, J.C.; Julander, J.G.; Olsen, A.L.; Polejaeva, I.A.; Davies, C.J.; White, K.L.; Lee, Y.M. Functional Genomics and Immunologic Tools: The Impact of Viral and Host Genetic Variations on the Outcome of Zika Virus Infection. Viruses 2018, 10, 422. [Google Scholar] [CrossRef]
- Barr, K.L.; Anderson, B.D.; Prakoso, D.; Long, M.T. Working with Zika and Usutu Viruses In Vitro. PLoS Negl. Trop. Dis. 2016, 10, e0004931. [Google Scholar] [CrossRef] [PubMed]
- Ragan, I.K.; Blizzard, E.L.; Gordy, P.; Bowen, R.A. Investigating the Potential Role of North American Animals as Hosts for Zika Virus. Vector Borne Zoonotic Dis. 2017, 17, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.G.; Ksiazek, T.G.; Gubler, D.J.; Lubis, S.I.; Simanjuntak, G.; Lee, V.H.; Nalim, S.; Juslis, K.; See, R. A survey for arboviral antibodies in sera of humans and animals in Lombok, Republic of Indonesia. Ann. Trop. Med. Parasitol. 1983, 77, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Darwish, M.A.; Hoogstraal, H.; Roberts, T.J.; Ahmed, I.P.; Omar, F. A sero-epidemiological survey for certain arboviruses (Togaviridae) in Pakistan. Trans. R. Soc. Trop. Med. Hyg. 1983, 77, 442–445. [Google Scholar] [CrossRef]
- Johnson, B.K.; Chanas, A.C.; Shockley, P.; Squires, E.J.; Gardner, P.; Wallace, C.; Simpson, D.I.; Bowen, E.T.; Platt, G.S.; Way, H.; et al. Arbovirus isolations from, and serological studies on, wild and domestic vertebrates from Kano Plain, Kenya. Trans. R. Soc. Trop. Med. Hyg. 1977, 71, 512–517. [Google Scholar] [CrossRef]
- Henderson, B.E.; Hewitt, L.E.E.; Lule, M. Serology of Wild Mammals; Virus Research Institute Annual Report; East African Printer: Nairobi, Kenya, 1968; pp. 48–51. [Google Scholar]
- Barry, J.S.; Rozance, P.J.; Anthony, R.V. An animal model of placental insufficiency-induced intrauterine growth restriction. Semin. Perinatol. 2008, 32, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Barry, J.S.; Anthony, R.V. The pregnant sheep as a model for human pregnancy. Theriogenology 2008, 69, 55–67. [Google Scholar] [CrossRef]
- Frasch, M.G.; Lobmaier, S.M.; Stampalija, T.; Desplats, P.; Pallarés, M.E.; Pastor, V.; Brocco, M.A.; Wu, H.T.; Schulkin, J.; Herry, C.L.; et al. Non-invasive biomarkers of fetal brain development reflecting prenatal stress: An integrative multi-scale multi-species perspective on data collection and analysis. Neurosci. Biobehav. Rev. 2018. [Google Scholar] [CrossRef]
- Herry, C.L.; Cortes, M.; Wu, H.T.; Durosier, L.D.; Cao, M.; Burns, p.; Desrochers, A.; Fecteau, G.; Seely, A.J.; Frasch, M.G. Temporal Patterns in Sheep Fetal Heart Rate Variability Correlate to Systemic Cytokine Inflammatory Response: A Methodological Exploration of Monitoring Potential Using Complex Signals Bioinformatics. PLoS ONE 2016, 11, e0153515. [Google Scholar] [CrossRef]
- Frasch, M.G.; Baier, C.J.; Antonelli, M.C.; Metz, G.A.S. Perinatal Psychoneuroimmunology: Protocols for the Study of Prenatal Stress and Its Effects on Fetal and Postnatal Brain Development. Methods Mol. Biol. 2018, 1781, 353–376. [Google Scholar] [CrossRef]
- Burns, P.; Liu, H.L.; Kuthiala, S.; Fecteau, G.; Desrochers, A.; Durosier, L.D.; Cao, M.; Frasch, M.G. Instrumentation of Near-term Fetal Sheep for Multivariate Chronic Non-anesthetized Recordings. J. Vis. Exp. 2015, e52581. [Google Scholar] [CrossRef] [PubMed]
- Ghesquière, L.; De Jonckheere, J.; Drumez, E.; Sharma, D.; Aubry, E.; Deruelle, P.; Storme, L.; Houfflin-Debarge, V.; Garabedian, C. Parasympathetic nervous system response to acidosis: Evaluation in an experimental fetal sheep model. Acta Obstet. Gynecol. Scand. 2019, 98, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Anthony, R.V.; Scheaffer, A.N.; Wright, C.D.; Regnault, T.R. Ruminant models of prenatal growth restriction. Reprod. Suppl. 2003, 61, 183–194. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sawyer, M.; Moe, J.; Osburn, B.I. Ontogeny of immunity and leukocytes in the ovine fetus and elevation of immunoglobulins related to congenital infection. Am. J. Vet. Res. 1978, 39, 643–648. [Google Scholar] [PubMed]
- Stonestreet, B.S.; Patlak, C.S.; Pettigrew, K.D.; Reilly, C.B.; Cserr, H.F. Ontogeny of blood-brain barrier function in ovine fetuses, lambs, and adults. Am. J. Physiol. 1996, 271, R1594–R1601. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.L.; Merriam, A.A.; Ohuma, E.O.; Dighe, M.K.; Gale, M.; Rajagopal, L.; Papageorghiou, A.T.; Gyamfi-Bannerman, C.; Adams Waldorf, K.M. Femur-sparing pattern of abnormal fetal growth in pregnant women from New York City after maternal Zika virus infection. Am. J. Obstet. Gynecol. 2018, 219, 187.e181–187.e120. [Google Scholar] [CrossRef] [PubMed]
- Yasri, S.; Wiwanitkit, V. Femur-sparing pattern of fetal growth after maternal Zika virus infection. Am. J. Obstet. Gynecol. 2018, 219, 514. [Google Scholar] [CrossRef] [PubMed]
- Barbera, A.; Jones, O.W.; Zerbe, G.O.; Hobbins, J.C.; Battaglia, F.C.; Meschia, G. Ultrasonographic assessment of fetal growth: Comparison between human and ovine fetus. Am. J. Obstet. Gynecol. 1995, 173, 1765–1769. [Google Scholar] [CrossRef]
- Leiser, R.; Krebs, C.; Ebert, B.; Dantzer, V. Placental vascular corrosion cast studies: A comparison between ruminants and humans. Microsc. Res. Tech. 1997, 38, 76–87. [Google Scholar] [CrossRef]
- Stegeman, J.H.J. Placental development in the sheep and its relation to fetal development. A qualitative and quantitative anatomic and histological study. Bijdr. Tot De Dierkd. 1974, 44, 72. [Google Scholar]
- Steven, D.H. Anatomy of the Placental Barrier; Academic Press: London, UK, 1975. [Google Scholar]
- Enright, F.M.; Osburn, B.I. Ontogeny of host responses in ovine fetuses infected with bluetongue virus. Am. J. Vet. Res. 1980, 41, 224–229. [Google Scholar] [PubMed]
- Parsonson, I.M.; Della-Porta, A.J.; Snowdon, W.A. Developmental disorders of the fetus in some arthropod-borne virus infections. Am. J. Trop. Med. Hyg. 1981, 30, 660–673. [Google Scholar] [CrossRef] [PubMed]
- van der Sluijs, M.; Timmermans, M.; Moulin, V.; Noordegraaf, C.V.; Vrijenhoek, M.; Debyser, I.; de Smit, A.J.; Moormann, R. Transplacental transmission of Bluetongue virus serotype 8 in ewes in early and mid gestation. Vet. Microbiol. 2011, 149, 113–125. [Google Scholar] [CrossRef] [PubMed]
- van der Sluijs, M.T.; Schroer-Joosten, D.P.; Fid-Fourkour, A.; Smit, M.; Vrijenhoek, M.P.; Moulin, V.; de Smit, A.J.; Moormann, R.J. Transplacental transmission of BTV-8 in sheep: BTV viraemia, antibody responses and vaccine efficacy in lambs infected in utero. Vaccine 2013, 31, 3726–3731. [Google Scholar] [CrossRef] [PubMed]
- Worwa, G.; Hilbe, M.; Ehrensperger, F.; Chaignat, V.; Hofmann, M.A.; Griot, C.; Maclachlan, N.J.; Thuer, B. Experimental transplacental infection of sheep with bluetongue virus serotype 8. Vet. Rec. 2009, 164, 499–500. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.I.; Livingston, C.W.; Edwards, J.F.; Crandell, R.W.; Shope, R.E.; Shelton, M.J.; Collisson, E.W. Evidence that Cache Valley virus induces congenital malformations in sheep. Vet. Microbiol. 1990, 21, 297–307. [Google Scholar] [CrossRef]
- Parsonson, I.M.; McPhee, D.A.; Della-Porta, A.J.; McClure, S.; McCullagh, P. Transmission of Akabane virus from the ewe to the early fetus (32 to 53 days). J. Comp. Pathol. 1988, 99, 215–227. [Google Scholar] [CrossRef]
- van den Brom, R.; Luttikholt, S.J.; Lievaart-Peterson, K.; Peperkamp, N.H.; Mars, M.H.; van der Poel, W.H.; Vellema, P. Epizootic of ovine congenital malformations associated with Schmallenberg virus infection. Tijdschr. Diergeneeskd. 2012, 137, 106–111. [Google Scholar]
- Poskin, A.; Verite, S.; Comtet, L.; Van der Stede, Y.; Cay, B.; De Regge, N. Persistence of the protective immunity and kinetics of the isotype specific antibody response against the viral nucleocapsid protein after experimental Schmallenberg virus infection of sheep. Vet. Res. 2015, 46, 119. [Google Scholar] [CrossRef]
- Peperkamp, N.H.; Luttikholt, S.J.; Dijkman, R.; Vos, J.H.; Junker, K.; Greijdanus, S.; Roumen, M.P.; van Garderen, E.; Meertens, N.; van Maanen, C.; et al. Ovine and Bovine Congenital Abnormalities Associated With Intrauterine Infection With Schmallenberg Virus. Vet. Pathol. 2015, 52, 1057–1066. [Google Scholar] [CrossRef]
- Brasil, P.; Pereira, J.P.; Moreira, M.E.; Ribeiro Nogueira, R.M.; Damasceno, L.; Wakimoto, M.; Rabello, R.S.; Valderramos, S.G.; Halai, U.A.; Salles, T.S.; et al. Zika Virus Infection in Pregnant Women in Rio de Janeiro. N. Engl. J. Med. 2016, 375, 2321–2334. [Google Scholar] [CrossRef] [PubMed]
- Calvet, G.; Aguiar, R.S.; Melo, A.S.O.; Sampaio, S.A.; de Filippis, I.; Fabri, A.; Araujo, E.S.M.; de Sequeira, P.C.; de Mendonça, M.C.L.; de Oliveira, L.; et al. Detection and sequencing of Zika virus from amniotic fluid of fetuses with microcephaly in Brazil: A case study. Lancet Infect. Dis. 2016, 16, 653–660. [Google Scholar] [CrossRef]
- Castro, M.C.; Han, Q.C.; Carvalho, L.R.; Victora, C.G.; França, G.V.A. Implications of Zika virus and congenital Zika syndrome for the number of live births in Brazil. Proc. Natl. Acad. Sci. USA 2018. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liang, Y.; Yi, P.; Xu, L.; Hawkins, H.K.; Rossi, S.L.; Soong, L.; Cai, J.; Menon, R.; Sun, J. Outcomes of Congenital Zika Disease Depend on Timing of Infection and Maternal-Fetal Interferon Action. Cell Rep. 2017, 21, 1588–1599. [Google Scholar] [CrossRef] [PubMed]
- Rice, M.E.; Galang, R.R.; Roth, N.M.; Ellington, S.R.; Moore, C.A.; Valencia-Prado, M.; Ellis, E.M.; Tufa, A.J.; Taulung, L.A.; Alfred, J.M.; et al. Vital Signs: Zika-Associated Birth Defects and Neurodevelopmental Abnormalities Possibly Associated with Congenital Zika Virus Infection—U.S. Territories and Freely Associated States, 2018. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 858–867. [Google Scholar] [CrossRef]
- Meaney-Delman, D.; Hills, S.L.; Williams, C.; Galang, R.R.; Iyengar, P.; Hennenfent, A.K.; Rabe, I.B.; Panella, A.; Oduyebo, T.; Honein, M.A.; et al. Zika Virus Infection Among U.S. Pregnant Travelers—August 2015–February 2016. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 211–214. [Google Scholar] [CrossRef]
- Hoen, B.; Schaub, B.; Funk, A.L.; Ardillon, V.; Boullard, M.; Cabié, A.; Callier, C.; Carles, G.; Cassadou, S.; Césaire, R.; et al. Pregnancy Outcomes after ZIKV Infection in French Territories in the Americas. N. Engl. J. Med. 2018, 378, 985–994. [Google Scholar] [CrossRef]
- Magnani, D.M.; Rogers, T.F.; Maness, N.J.; Grubaugh, N.D.; Beutler, N.; Bailey, V.K.; Gonzalez-Nieto, L.; Gutman, M.J.; Pedreño-Lopez, N.; Kwal, J.M.; et al. Fetal demise and failed antibody therapy during Zika virus infection of pregnant macaques. Nat. Commun. 2018, 9, 1624. [Google Scholar] [CrossRef]
- Citil Dogan, A.; Wayne, S.; Bauer, S.; Ogunyemi, D.; Kulkharni, S.K.; Maulik, D.; Carpenter, C.F.; Bahado-Singh, R.O. The Zika virus and pregnancy: Evidence, management, and prevention. J. Matern. Fetal Neonatal Med. 2017, 30, 386–396. [Google Scholar] [CrossRef][Green Version]
- Seino, K.K.; Long, M.T.; Gibbs, E.P.; Bowen, R.A.; Beachboard, S.E.; Humphrey, P.P.; Dixon, M.A.; Bourgeois, M.A. Comparative efficacies of three commercially available vaccines against West Nile Virus (WNV) in a short-duration challenge trial involving an equine WNV encephalitis model. Clin. Vaccine Immunol. 2007, 14, 1465–1471. [Google Scholar] [CrossRef]
- Rio, D.C.; Ares, M.; Hannon, G.J.; Nilsen, T.W. Purification of RNA using TRIzol (TRI reagent). Cold Spring Harb. Protoc. 2010, 2010. [Google Scholar] [CrossRef] [PubMed]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Velez, J.O.; Lambert, A.J.; Johnson, A.J.; Stanfield, S.M.; Duffy, M.R. Genetic and serologic properties of Zika virus associated with an epidemic, Yap State, Micronesia, 2007. Emerg. Infect. Dis. 2008, 14, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Long, M.T.; Jeter, W.; Hernandez, J.; Sellon, D.C.; Gosche, D.; Gillis, K.; Bille, E.; Gibbs, E.P. Diagnostic performance of the equine IgM capture ELISA for serodiagnosis of West Nile virus infection. J. Vet. Intern. Med. 2006, 20, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, V.; Fineschi, N.; Matteucci, G.; Marsili, I.; Nencioni, L.; Puddu, M.; Garelick, H.; Zuckerman, A.J. Preparation and immunogenicity of an inactivated hepatitis A vaccine. Vaccine 1993, 11, 383–387. [Google Scholar] [CrossRef]
- Barr, K.L.; Khan, E.; Farooqi, J.Q.; Imtiaz, K.; Prakoso, D.; Malik, F.; Lednicky, J.A.; Long, M.T. Evidence of Chikungunya Virus Disease in Pakistan Since 2015 With Patients Demonstrating Involvement of the Central Nervous System. Front. Public Health 2018, 6, 186. [Google Scholar] [CrossRef] [PubMed]
- Khan, E.; Barr, K.L.; Farooqi, J.Q.; Prakoso, D.; Abbas, A.; Khan, Z.Y.; Ashi, S.; Imtiaz, K.; Aziz, Z.; Malik, F.; et al. Human West Nile Virus Disease Outbreak in Pakistan, 2015–2016. Front. Public Health 2018, 6, 20. [Google Scholar] [CrossRef] [PubMed]
- Abella, N.; Schelcher, F.; Delverdier, M.; Concordet, D.; Valarcher, J.F.; Espinasse, J.; Cabanie, P. Flow cytometric analysis of bovine CD4 and CD8 lymphocytes: Influence of blood sampling and processing methods. Res. Vet. Sci. 1994, 57, 163–171. [Google Scholar] [CrossRef]
- Shapiro-Mendoza, C.K.; Rice, M.E.; Galang, R.R.; Fulton, A.C.; VanMaldeghem, K.; Prado, M.V.; Ellis, E.; Anesi, M.S.; Simeone, R.M.; Petersen, E.E.; et al. Pregnancy Outcomes After Maternal Zika Virus Infection During Pregnancy - U.S. Territories, January 1, 2016-April 25, 2017. MMWR Morb. Mortal. Wkly. Rep. 2017, 66, 615–621. [Google Scholar] [CrossRef]
- Eppes, C.; Rac, M.; Dunn, J.; Versalovic, J.; Murray, K.O.; Suter, M.A.; Sanz Cortes, M.; Espinoza, J.; Seferovic, M.D.; Lee, W.; et al. Testing for Zika virus infection in pregnancy: Key concepts to deal with an emerging epidemic. Am. J. Obstet. Gynecol. 2017, 216, 209–225. [Google Scholar] [CrossRef]
- Dudley, D.M.; Van Rompay, K.K.; Coffey, L.L.; Ardeshir, A.; Keesler, R.I.; Bliss-Moreau, E.; Grigsby, P.L.; Steinbach, R.J.; Hirsch, A.J.; MacAllister, R.P.; et al. Miscarriage and stillbirth following maternal Zika virus infection in nonhuman primates. Nat. Med. 2018, 24, 1104–1107. [Google Scholar] [CrossRef]
- Hirsch, A.J.; Roberts, V.H.J.; Grigsby, P.L.; Haese, N.; Schabel, M.C.; Wang, X.; Lo, J.O.; Liu, Z.; Kroenke, C.D.; Smith, J.L.; et al. Zika virus infection in pregnant rhesus macaques causes placental dysfunction and immunopathology. Nat. Commun. 2018, 9, 263. [Google Scholar] [CrossRef] [PubMed]
- Szaba, F.M.; Tighe, M.; Kummer, L.W.; Lanzer, K.G.; Ward, J.M.; Lanthier, P.; Kim, I.J.; Kuki, A.; Blackman, M.A.; Thomas, S.J.; et al. Zika virus infection in immunocompetent pregnant mice causes fetal damage and placental pathology in the absence of fetal infection. PLoS Pathog. 2018, 14, e1006994. [Google Scholar] [CrossRef] [PubMed]
- Zacharias, N.; Whitty, J.; Noblin, S.; Tsakiri, S.; Garcia, J.; Covinsky, M.; Bhattacharjee, M.; Saulino, D.; Tatevian, N.; Blackwell, S. First Neonatal Demise with Travel-Associated Zika Virus Infection in the United States of America. AJP Rep. 2017, 7, e68–e73. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Coetzer, J.A.; Barnard, B.J. Hydrops amnii in sheep associated with hydranencephaly and arthrogryposis with wesselsbron disease and rift valley fever viruses as aetiological agents. Onderstepoort J. Vet. Res. 1977, 44, 119–126. [Google Scholar] [PubMed]
- Haby, M.M.; Pinart, M.; Elias, V.; Reveiz, L. Prevalence of asymptomatic Zika virus infection: A systematic review. Bull. World Health Organ. 2018, 96, 402–413D. [Google Scholar] [CrossRef] [PubMed]
- Mohr, E.L.; Block, L.N.; Newman, C.M.; Stewart, L.M.; Koenig, M.; Semler, M.; Breitbach, M.E.; Teixeira, L.B.C.; Zeng, X.; Weiler, A.M.; et al. Ocular and uteroplacental pathology in a macaque pregnancy with congenital Zika virus infection. PLoS ONE 2018, 13, e0190617. [Google Scholar] [CrossRef] [PubMed]
- Wilder-Smith, A.; Chang, C.R.; Leong, W.Y. Zika in travellers 1947–2017: A systematic review. J. Travel Med. 2018, 25. [Google Scholar] [CrossRef] [PubMed]
- Judice, C.C.; Tan, J.J.L.; Parise, P.L.; Kam, Y.W.; Milanez, G.P.; Leite, J.A.; Caserta, L.C.; Arns, C.W.; Resende, M.R.; Angerami, R.; et al. Efficient detection of Zika virus RNA in patients’ blood from the 2016 outbreak in Campinas, Brazil. Sci. Rep. 2018, 8, 4012. [Google Scholar] [CrossRef] [PubMed]
- Michlmayr, D.; Andrade, P.; Gonzalez, K.; Balmaseda, A.; Harris, E. CD14(+)CD16(+) monocytes are the main target of Zika virus infection in peripheral blood mononuclear cells in a paediatric study in Nicaragua. Nat. Microbiol. 2017, 2, 1462–1470. [Google Scholar] [CrossRef]
- Musso, D.; Rouault, E.; Teissier, A.; Lanteri, M.C.; Zisou, K.; Broult, J.; Grange, E.; Nhan, T.X.; Aubry, M. Molecular detection of Zika virus in blood and RNA load determination during the French Polynesian outbreak. J. Med. Virol. 2017, 89, 1505–1510. [Google Scholar] [CrossRef]
- Aliota, M.T.; Dudley, D.M.; Newman, C.M.; Mohr, E.L.; Gellerup, D.D.; Breitbach, M.E.; Buechler, C.R.; Rasheed, M.N.; Mohns, M.S.; Weiler, A.M.; et al. Heterologous Protection against Asian Zika Virus Challenge in Rhesus Macaques. PLoS Negl. Trop. Dis. 2016, 10, e0005168. [Google Scholar] [CrossRef] [PubMed]
- Martinot, A.J.; Abbink, P.; Afacan, O.; Prohl, A.K.; Bronson, R.; Hecht, J.L.; Borducchi, E.N.; Larocca, R.A.; Peterson, R.L.; Rinaldi, W.; et al. Fetal Neuropathology in Zika Virus-Infected Pregnant Female Rhesus Monkeys. Cell 2018, 173, 1111–1122.e10. [Google Scholar] [CrossRef] [PubMed]
- Coffey, L.L.; Pesavento, P.A.; Keesler, R.I.; Singapuri, A.; Watanabe, J.; Watanabe, R.; Yee, J.; Bliss-Moreau, E.; Cruzen, C.; Christe, K.L.; et al. Zika Virus Tissue and Blood Compartmentalization in Acute Infection of Rhesus Macaques. PLoS ONE 2017, 12, e0171148. [Google Scholar] [CrossRef] [PubMed]
- Foo, S.S.; Chen, W.; Chan, Y.; Bowman, J.W.; Chang, L.C.; Choi, Y.; Yoo, J.S.; Ge, J.; Cheng, G.; Bonnin, A.; et al. Asian Zika virus strains target CD14. Nat. Microbiol. 2017, 2, 1558–1570. [Google Scholar] [CrossRef] [PubMed]
- Roland, L.; Drillich, M.; Iwersen, M. Hematology as a diagnostic tool in bovine medicine. J. Vet. Diagn. Investig. 2014, 26, 592–598. [Google Scholar] [CrossRef] [PubMed]
- El-Mandrawy, S.A.M.; Alam, R.T.M. Hematological, biochemical and oxidative stress studies of lumpy skin disease virus infection in cattle. J. Appl. Anim. Res. 2018, 46, 1073–1077. [Google Scholar] [CrossRef]
- Larruskain, A.; Jugo, B. Retroviral Infections in Sheep and Goats: Small Ruminant Lentiviruses and Host Interaction. Viruses 2013, 5, 2043–2061. [Google Scholar] [CrossRef]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schwarz, E.R.; Pozor, M.A.; Pu, R.; Barr, K.L.; Beachboard, S.E.; MacLachlan, N.J.; Prakoso, D.; Long, M.T. Experimental Infection of Pregnant Female Sheep with Zika Virus During Early Gestation. Viruses 2019, 11, 795. https://doi.org/10.3390/v11090795
Schwarz ER, Pozor MA, Pu R, Barr KL, Beachboard SE, MacLachlan NJ, Prakoso D, Long MT. Experimental Infection of Pregnant Female Sheep with Zika Virus During Early Gestation. Viruses. 2019; 11(9):795. https://doi.org/10.3390/v11090795
Chicago/Turabian StyleSchwarz, Erika R., Malgorzata A. Pozor, Ruiyu Pu, Kelli L. Barr, Sarah E. Beachboard, N. James MacLachlan, Dhani Prakoso, and Maureen T. Long. 2019. "Experimental Infection of Pregnant Female Sheep with Zika Virus During Early Gestation" Viruses 11, no. 9: 795. https://doi.org/10.3390/v11090795
APA StyleSchwarz, E. R., Pozor, M. A., Pu, R., Barr, K. L., Beachboard, S. E., MacLachlan, N. J., Prakoso, D., & Long, M. T. (2019). Experimental Infection of Pregnant Female Sheep with Zika Virus During Early Gestation. Viruses, 11(9), 795. https://doi.org/10.3390/v11090795