Reliable and Standardized Animal Models to Study the Pathogenesis of Bluetongue and Schmallenberg Viruses in Ruminant Natural Host Species with Special Emphasis on Placental Crossing

 and
and 
Abstract
1. Introduction
2. Studying the Pathogenesis and Immune Response of BTV and SBV in Natural Ruminant Host Species
2.1. Selection of an Appropriate Inoculum is Crucial to Achieve Adequate Experimental Infection
- (1)
- Safe, meaning it should have been screened for contaminations, adventitial agents or other pathogens [9];
- (2)
- Easily available, practical and standardised; and
- (3)
- Made of a virus displaying similar replication and virulence properties than wild-type.
2.1.1. Infectious Blood versus Cell Passaged Inoculum
- (1)
- Original isolate or any strain of particular interest can be shipped almost anywhere in the world, leading to great improvement of standardization;
- (2)
- Viral amplification by cell-passages allows a high increase in viral titre, subsequently allowing to inoculate lower volumes;
- (3)
- Screening for contamination or other pathogens is easier in cell culture and eliminate some veterinary public health concerns about using ruminant blood to infect other ruminants; and
- (4)
- Virulence in cell culture can be easily standardized.
2.1.2. A Matter of Doses and Routes
- (1)
- The need for a route that best mimics the behaviour of the vector in field conditions. Usually haematophagous arthropods are either telmophagous or solenophagous; depending on the vector species the route might be intradermal (ID), subcutaneous (SC), or intravenous (IV). In experimental infections the inoculated viral load and volume are usually higher than the ones inoculated through naturally occurring feeding given the size of the arthropods and the size of their mouthparts [51]. Another drawback already mentioned is the lack of vector saliva components, which can modify the structure and infectivity of Reoviridae and Peribunyaviridae viral particles [43,52].
- (2)
- The need for a route that will ensure the virus to reach the blood stream. Quite obviously this is the intravenous route. Since vector saliva components can enhance the infectivity of arboviruses there is a risk that the inoculation of the virus alone or at a distal site from the vector feeding site could result in a failed infection [53]. Therefore, the option to bypass the skin for reaching the bloodstream may be relevant.
- (1)
- The Culicoides saliva contains proteases able to cleave VP2, leading to the formation of infectious subviral particles (ISVP) displaying higher infectivity in KC cells and Culicoides [43];
- (2)
- The ratio of infectious BTV particles versus defective virions produced within Culicoides might be higher when compared with cell culture grown BTV [66]; and
- (3)
2.1.3. Screening for Concomitant Pathogens
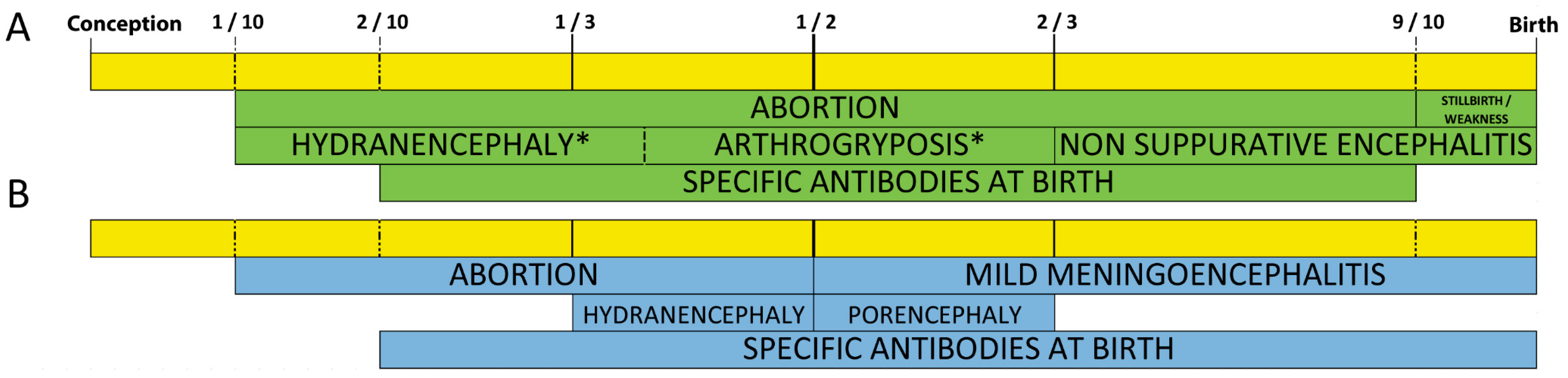
3. BTV and SBV Display Placental Crossing Abilities and Teratogenic Potential
4. Conclusion and Future Prospects
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Carrasco-Hernandez, R.; Jácome, R.; Vidal, Y.L.; De León, S.P. Are RNA Viruses Candidate Agents for the Next Global Pandemic? A Review. ILAR J. 2017, 58, 343–358. [Google Scholar] [CrossRef] [PubMed]
- Mertens, P.P.; Maan, S.; Samuel, A.; Attoui, H. Orbivirus, Reoviridae. In Virus Taxonomy, VIIIth Report of the ICTV; Fauquet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U., Ball, L.A., Eds.; Elsevier/Academic Press: Oxford, UK, 2005; pp. 466–483. [Google Scholar]
- Mertens, P.P.; Diprose, J.; Maan, S.; Singh, K.P.; Attoui, H.; Samuel, A.R. Bluetongue virus replication, molecular and structural biology. Vet. Ital. 2004, 40, 426–437. [Google Scholar] [PubMed]
- Attoui, H.; Maan, S.; Anthony, S.J.; Mertens, P.P.C. Bluetongue virus, other orbiviruses and other reoviruses: Their relationships and taxonomy. In Bluetongue; Elsevier/Academic Press: London, UK, 2009; Volume 1, pp. 23–52. [Google Scholar]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.Q.; Harrach, B.; Harrison, R.L.; Knowles, N.J.; Kropinski, A.M.; Krupovic, M.; Kuhn, J.H.; Mushegian, A.R.; et al. Changes to taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2017). Arch. Virol. 2017, 162, 2505–2538. [Google Scholar] [CrossRef] [PubMed]
- Kinney, R.M.; Calisher, C.H. Antigenic relationships among Simbu serogroup (Bunyaviridae) viruses. Am. J. Trop. Med. Hyg. 1981, 30, 1307–1318. [Google Scholar] [CrossRef] [PubMed]
- De Regge, N. Akabane, Aino and Schmallenberg virus-where do we stand and what do we know about the role of domestic ruminant hosts and Culicoides vectors in virus transmission and overwintering? Curr. Opin. Virol. 2017, 27, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, P.; van Vuuren, M.; Venter, E.H.; Stokstad, M. A review of experimental infections with bluetongue virus in the mammalian host. Virus Res. 2014, 182, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Speder, B. Regulatory Requirements for Viral-Challenge Studies: Influenza Case Study; SGS Life Science Services, Ed.; SGS Life Science Services: London, UK, 2014; p. 5. [Google Scholar]
- Bréard, E.; Schulz, C.; Sailleau, C.; Bernelin-Cottet, C.; Viarouge, C.; Vitour, D.; Guillaume, B.; Caignard, G.; Gorlier, A.; Attoui, H.; et al. Bluetongue virus serotype 27: Experimental infection of goats, sheep and cattle with three BTV-27 variants reveal atypical characteristics and likely direct contact transmission BTV-27 between goats. Transbound. Emerg. Dis. 2018, 65, e251–e263. [Google Scholar] [CrossRef] [PubMed]
- MacLachlan, N.J.; Nunamaker, R.A.; Katz, J.B.; Sawyer, M.M.; Akita, G.Y.; Osburn, B.I.; Tabachnick, W.J. Detection of bluetongue virus in the blood of inoculated calves: Comparison of virus isolation, PCR assay, and in vitro feeding of Culicoides variipennis. Arch. Virol. 1994, 136, 1–8. [Google Scholar] [CrossRef]
- Flannery, J.; Sanz-Bernardo, B.; Ashby, M.; Brown, H.; Carpenter, S.; Cooke, L.; Corla, A.; Frost, L.; Gubbins, S.; Hicks, H.; et al. Evidence of reduced viremia, pathogenicity and vector competence in a re-emerging European strain of bluetongue virus serotype 8 in sheep. Transbound. Emerg. Dis. 2019, 66, 1177–1185. [Google Scholar] [CrossRef]
- Putty, K.; Shaik, A.M.; Peera, S.J.; Reddy, Y.N.; Rao, P.P.; Patil, S.R.; Reddy, M.S.; Susmitha, B.; Jyothi, J.S. Infection kinetics and antibody responses in Deccani sheep during experimental infection and superinfection with bluetongue virus serotypes 4 and 16. Vet. World 2019, 12, 41–47. [Google Scholar] [CrossRef]
- Martinelle, L.; Dal Pozzo, F.; Thys, C.; De Leeuw, I.; Van Campe, W.; De Clercq, K.; Thiry, E.; Saegerman, C. Assessment of cross-protection induced by a bluetongue virus (BTV) serotype 8 vaccine towards other BTV serotypes in experimental conditions. Vet. Res. 2018, 49, 63. [Google Scholar] [CrossRef] [PubMed]
- Schulz, C.; Sailleau, C.; Bréard, E.; Flannery, J.; Viarouge, C.; Zientara, S.; Beer, M.; Batten, C.; Hoffmann, B. Experimental infection of sheep, goats and cattle with a bluetongue virus serotype 4 field strain from Bulgaria, 2014. Transbound Emerg Dis. 2018, 65, e243–e250. [Google Scholar] [CrossRef] [PubMed]
- van Rijn, P.A.; van de Water, S.G.P.; Maris-Veldhuis, M.A.; van Gennip, R.G.P. Experimental infection of small ruminants with bluetongue virus expressing Toggenburg Orbivirus proteins. Vet Microbiol. 2016, 192, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Martinelle, L.; Dal Pozzo, F.; Sarradin, P.; Van Campe, W.; De Leeuw, I.; De Clercq, K.; Thys, C.; Thiry, E.; Saegerman, C. Experimental bluetongue virus superinfection in calves previously immunized with bluetongue virus serotype 8. Vet. Res. 2016, 47, 73. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Darpel, K.E.; Barber, J.; Hope, A.; Wilson, A.J.; Gubbins, S.; Henstock, M.; Frost, L.; Batten, C.; Veronesi, E.; Moffat, K.; et al. Using shared needles for subcutaneous inoculation can transmit bluetongue virus mechanically between ruminant hosts. Sci Rep. 2016, 6, 20627. [Google Scholar] [CrossRef] [PubMed]
- Drolet, B.S.; Reister, L.M.; Lehiy, C.J.; Van Rijn, P.A.; Bowen, R.A. Effect of Culicoides sonorensis salivary proteins on clinical disease outcome in experimental bluetongue virus serotype 8 infection of Dorset sheep. Vet. Ital. 2015, 51, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Bréard, E.; Belbis, G.; Viarouge, C.; Nomikou, K.; Haegeman, A.; De Clercq, K.; Hudelet, P.; Hamers, C.; Moreau, F.; Lilin, T.; et al. Evaluation of adaptive immune responses and heterologous protection induced by inactivated bluetongue virus vaccines. Vaccine 2015, 33, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Kesik-Maliszewska, J.; Pomorska-Mol, M.; Collins, A.B.; Rola, J.; Larska, M. Potential use of hematological and acute phase protein parameters in the diagnosis of acute Schmallenberg virus infection in experimentally infected calves. Comp. Immunol. Microbiol. Infect. Dis. 2019, 64, 146–152. [Google Scholar] [CrossRef]
- Konig, P.; Wernike, K.; Hechinger, S.; Tauscher, K.; Breithaupt, A.; Beer, M. Fetal infection with Schmallenberg virus—An experimental pathogenesis study in pregnant cows. Transbound. Emerg Dis. 2019, 66, 454–462. [Google Scholar] [CrossRef]
- Endalew, A.D.; Morozov, I.; Davis, A.S.; Gaudreault, N.N.; Wernike, K.; Bawa, B.; Ruder, M.G.; Drolet, B.S.; McVey, D.S.; Shivanna, V.; et al. Virological and Serological Responses of Sheep and Cattle to Experimental Schmallenberg Virus Infection. Vector-Borne Zoonotic Dis. 2018. [Google Scholar] [CrossRef]
- Laloy, E.; Bréard, E.; Trapp, S.; Pozzi, N.; Riou, M.; Barc, C.; Breton, S.; Delaunay, R.; Cordonnier, N.; Chateau-Joubert, S.; et al. Fetopathic effects of experimental Schmallenberg virus infection in pregnant goats. Vet Microbiol. 2017, 211, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Prieto, V.; Kukielka, D.; Mourino, M.; Paradell, H.; Plaja, L.; Urniza, A.; Sanchez-Vizcaino, J.M. Natural Immunity of Sheep and Lambs Against the Schmallenberg Virus Infection. Transbound. Emerg. Dis. 2016, 63, e220–e228. [Google Scholar] [CrossRef] [PubMed]
- Poskin, A.; Verite, S.; Comtet, L.; Van der Stede, Y.; Cay, B.; De Regge, N. Persistence of the protective immunity and kinetics of the isotype specific antibody response against the viral nucleocapsid protein after experimental Schmallenberg virus infection of sheep. Vet Res. 2015, 46, 119. [Google Scholar] [CrossRef] [PubMed]
- Martinelle, L.; Poskin, A.; Dal Pozzo, F.; De Regge, N.; Cay, B.; Saegerman, C. Experimental Infection of Sheep at 45 and 60 Days of Gestation with Schmallenberg Virus Readily Led to Placental Colonization without Causing Congenital Malformations. PLoS ONE 2015, 10, e0139375. [Google Scholar] [CrossRef] [PubMed]
- Martinelle, L.; Poskin, A.; Dal Pozzo, F.; Mostin, L.; Van Campe, W.; Cay, A.B.; De Regge, N.; Saegerman, C. Three Different Routes of Inoculation for Experimental Infection with Schmallenberg Virus in Sheep. Transbound. Emerg. Dis. 2017, 64, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Laloy, E.; Riou, M.; Barc, C.; Belbis, G.; Bréard, E.; Breton, S.; Cordonnier, N.; Crochet, D.; Delaunay, R.; Moreau, J.; et al. Schmallenberg virus: Experimental infection in goats and bucks. BMC Vet. Res. 2015, 11, 221. [Google Scholar] [CrossRef]
- Poskin, A.; Martinelle, L.; Mostin, L.; Van Campe, W.; Dal Pozzo, F.; Saegerman, C.; Cay, A.B.; De Regge, N. Dose-dependent effect of experimental Schmallenberg virus infection in sheep. Vet. J. 2014, 201, 419–422. [Google Scholar] [CrossRef]
- Pourianfar, H.R.; Javadi, A.; Grollo, L. A colorimetric-based accurate method for the determination of enterovirus 71 titer. Indian, J. Virol. 2012, 23, 303–310. [Google Scholar] [CrossRef]
- Martinelle, L.; Dal Pozzo, F.; Sarradin, P.; De Leeuw, I.; De Clercq, K.; Thys, C.; Thiry, E.; Saegerman, C. Pulmonary artery haemorrhage in newborn calves following bluetongue virus serotype 8 experimental infections of pregnant heifers. Vet. Microbiol. 2013, 167, 250–259. [Google Scholar] [CrossRef]
- Martinelle, L.; Dal Pozzo, F.; Sarradin, P.; De Leeuw, I.; De Clercq, K.; Thys, C.; Ziant, D.; Thiry, E.; Saegerman, C. Two alternative inocula to reproduce bluetongue virus serotype 8 disease in calves. Vaccine 2011, 29, 3600–3609. [Google Scholar] [CrossRef]
- Dal Pozzo, F.; Martinelle, L.; Thys, C.; Sarradin, P.; De Leeuw, I.; Van Campe, W.; De Clercq, K.; Thiry, E.; Saegerman, C. Experimental co-infections of calves with bluetongue virus serotypes 1 and 8. Vet. Microbiol. 2013, 165, 167–172. [Google Scholar] [CrossRef][Green Version]
- Moulin, V.; Noordegraaf, C.V.; Makoschey, B.; van der Sluijs, M.; Veronesi, E.; Darpel, K.; Mertens, P.P.; de Smit, H. Clinical disease in sheep caused by bluetongue virus serotype 8, and prevention by an inactivated vaccine. Vaccine 2012, 30, 2228–2235. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.; Hagglund, S.; Bréard, E.; Riou, M.; Zohari, S.; Comtet, L.; Olofson, A.S.; Gelineau, R.; Martin, G.; Elvander, M.; et al. Strong protection induced by an experimental DIVA subunit vaccine against bluetongue virus serotype 8 in cattle. Vaccine 2014, 32, 6614–6621. [Google Scholar] [CrossRef] [PubMed]
- Domingo, E.; Holland, J.J. RNA virus mutations and fitness for survival. Annu. Rev. Microbiol. 1997, 51, 151–178. [Google Scholar] [CrossRef]
- Bonneau, K.R.; Mullens, B.A.; MacLachlan, N.J. Occurrence of genetic drift and founder effect during quasispecies evolution of the VP2 and NS3/NS3A genes of bluetongue virus upon passage between sheep, cattle, and Culicoides sonorensis. J. Virol. 2001, 75, 8298–8305. [Google Scholar] [CrossRef] [PubMed]
- Biebricher, C.K.; Eigen, M. What is a quasispecies? In Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2006; Volume 299, pp. 1–31. [Google Scholar]
- Domingo, E.; Escarmis, C.; Sevilla, N.; Moya, A.; Elena, S.F.; Quer, J.; Novella, I.S.; Holland, J.J. Basic concepts in RNA virus evolution. FASEB J. 1996, 10, 859–864. [Google Scholar] [CrossRef]
- Fischer, M.; Hoffmann, B.; Goller, K.V.; Hoper, D.; Wernike, K.; Beer, M. A mutation ‘hot spot’ in the Schmallenberg virus M segment. J. Gen. Virol. 2013, 94, 1161–1167. [Google Scholar] [CrossRef]
- Caporale, M.; Di Gialleonorado, L.; Janowicz, A.; Wilkie, G.; Shaw, A.; Savini, G.; Van Rijn, P.A.; Mertens, P.; Di Ventura, M.; Palmarini, M. Virus and host factors affecting the clinical outcome of bluetongue virus infection. J. Virol. 2014, 88, 10399–10411. [Google Scholar] [CrossRef]
- Darpel, K.E.; Langner, K.F.; Nimtz, M.; Anthony, S.J.; Brownlie, J.; Takamatsu, H.H.; Mellor, P.S.; Mertens, P.P. Saliva proteins of vector Culicoides modify structure and infectivity of bluetongue virus particles. PLoS ONE 2011, 6, e17545. [Google Scholar] [CrossRef][Green Version]
- Eschbaumer, M.; Wackerlin, R.; Rudolf, M.; Keller, M.; Konig, P.; Zemke, J.; Hoffmann, B.; Beer, M. Infectious blood or culture-grown virus: A comparison of bluetongue virus challenge models. Vet. Microbiol. 2010, 146, 150–154. [Google Scholar] [CrossRef]
- OIE. Bluetongue. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; Health WOfA, Ed.; Manual of Diagnostic Tests and Vaccines for Terrestrial Animals: Paris, France, 2014; pp. 1–18. [Google Scholar]
- Wernike, K.; Eschbaumer, M.; Breithaupt, A.; Hoffmann, B.; Beer, M. Schmallenberg virus challenge models in cattle: Infectious serum or culture-grown virus? Vet. Res. 2012, 43, 84. [Google Scholar] [CrossRef] [PubMed]
- Wernike, K.; Hoffmann, B.; Bréard, E.; Botner, A.; Ponsart, C.; Zientara, S.; Lohse, L.; Pozzi, N.; Viarouge, C.; Sarradin, P.; et al. Schmallenberg virus experimental infection of sheep. Vet. Microbiol. 2013, 166, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Varela, M.; Schnettler, E.; Caporale, M.; Murgia, C.; Barry, G.; McFarlane, M.; McGregor, E.; Piras, I.M.; Shaw, A.; Lamm, C.; et al. Schmallenberg virus pathogenesis, tropism and interaction with the innate immune system of the host. PLOS Pathog. 2013, 9, e1003133. [Google Scholar] [CrossRef] [PubMed]
- Kraatz, F.; Wernike, K.; Hechinger, S.; Konig, P.; Granzow, H.; Reimann, I.; Beer, M. Deletion mutants of Schmallenberg virus are avirulent and protect from virus challenge. J. Virol. 2015, 89, 1825–1837. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, M.A.; Mader, M.; Fluckiger, F.; Renzullo, S. Genetic stability of Schmallenberg virus in vivo during an epidemic, and in vitro, when passaged in the highly susceptible porcine SK-6 cell line. Vet. Microbiol. 2015, 176, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Venter, G.J.; Mellor, P.S.; Wright, I.; Paweska, J.T. Replication of live-attenuated vaccine strains of bluetongue virus in orally infected South African Culicoides species. Med. Vet. Entomol. 2007, 21, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Horne, K.M.; Vanlandingham, D.L. Bunyavirus-vector interactions. Viruses 2014, 6, 4373–4397. [Google Scholar] [CrossRef] [PubMed]
- Le Coupanec, A.; Babin, D.; Fiette, L.; Jouvion, G.; Ave, P.; Misse, D.; Bouloy, M.; Choumet, V. Aedes mosquito saliva modulates Rift Valley fever virus pathogenicity. PLOS Neglected Trop. Dis. 2013, 7, e2237. [Google Scholar] [CrossRef] [PubMed]
- Dal Pozzo, F.; De Clercq, K.; Guyot, H.; Vandemeulebroucke, E.; Sarradin, P.; Vandenbussche, F.; Thiry, E.; Saegerman, C. Experimental reproduction of bluetongue virus serotype 8 clinical disease in calves. Vet. Microbiol. 2009, 136, 352–358. [Google Scholar] [CrossRef]
- Nicolas, J.F.; Guy, B. Intradermal, epidermal and transcutaneous vaccination: From immunology to clinical practice. Expert Rev. Vaccines 2008, 7, 1201–1214. [Google Scholar] [CrossRef]
- Umeshappa, C.S.; Singh, K.P.; Channappanavar, R.; Sharma, K.; Nanjundappa, R.H.; Saxena, M.; Singh, R.; Sharma, A.K. A comparison of intradermal and intravenous inoculation of bluetongue virus serotype 23 in sheep for clinico-pathology, and viral and immune responses. Vet. Immunol. Immunopathol. 2011, 141, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Melzi, E.; Caporale, M.; Rocchi, M.; Martin, V.; Gamino, V.; di Provvido, A.; Marruchella, G.; Entrican, G.; Sevilla, N.; Palmarini, M. Follicular dendritic cell disruption as a novel mechanism of virus-induced immunosuppression. Proc. Natl. Acad. Sci. USA 2016, 113, E6238–E6247. [Google Scholar] [CrossRef] [PubMed]
- van der Sluijs, M.; Timmermans, M.; Moulin, V.; Noordegraaf, C.V.; Vrijenhoek, M.; Debyser, I.; de Smit, A.J.; Moormann, R. Transplacental transmission of Bluetongue virus serotype 8 in ewes in early and mid gestation. Vet. Microbiol. 2011, 149, 113–125. [Google Scholar] [CrossRef] [PubMed]
- van der Sluijs, M.T.; Schroer-Joosten, D.P.; Fid-Fourkour, A.; Vrijenhoek, M.P.; Debyser, I.; Moulin, V.; Moormann, R.J.; de Smit, A.J. Transplacental transmission of Bluetongue virus serotype 1 and serotype 8 in sheep: Virological and pathological findings. PLoS ONE 2013, 8, e81429. [Google Scholar] [CrossRef] [PubMed]
- Batten, C.A.; Henstock, M.R.; Steedman, H.M.; Waddington, S.; Edwards, L.; Oura, C.A. Bluetongue virus serotype 26: Infection kinetics, pathogenesis and possible contact transmission in goats. Vet. Microbiol. 2013, 162, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Batten, C.; Darpel, K.; Henstock, M.; Fay, P.; Veronesi, E.; Gubbins, S.; Graves, S.; Frost, L.; Oura, C. Evidence for transmission of bluetongue virus serotype 26 through direct contact. PLoS ONE 2014, 9, e96049. [Google Scholar] [CrossRef] [PubMed]
- Alexandersen, S.; Quan, M.; Murphy, C.; Knight, J.; Zhang, Z. Studies of quantitative parameters of virus excretion and transmission in pigs and cattle experimentally infected with foot-and-mouth disease virus. J. Comp. Pathol. 2003, 129, 268–282. [Google Scholar] [CrossRef]
- Howey, R.; Quan, M.; Savill, N.J.; Matthews, L.; Alexandersen, S.; Woolhouse, M. Effect of the initial dose of foot-and-mouth disease virus on the early viral dynamics within pigs. J. R. Soc. Interface 2009, 6, 835–847. [Google Scholar] [CrossRef] [PubMed]
- Quan, M.; Murphy, C.M.; Zhang, Z.; Alexandersen, S. Determinants of early foot-and-mouth disease virus dynamics in pigs. J. Comp. Pathol. 2004, 131, 294–307. [Google Scholar] [CrossRef]
- Di Gialleonardo, L.; Migliaccio, P.; Teodori, L.; Savini, G. The length of BTV-8 viraemia in cattle according to infection doses and diagnostic techniques. Res. Vet. Sci. 2011, 91, 316–320. [Google Scholar] [CrossRef]
- Pages, N.; Bréard, E.; Urien, C.; Talavera, S.; Viarouge, C.; Lorca-Oro, C.; Jouneau, L.; Charley, B.; Zientara, S.; Bensaid, A.; et al. Culicoides midge bites modulate the host response and impact on bluetongue virus infection in sheep. PLoS ONE 2014, 9, e83683. [Google Scholar] [CrossRef] [PubMed]
- Bishop, J.V.; Mejia, J.S.; Perez de Leon, A.A.; Tabachnick, W.J.; Titus, R.G. Salivary gland extracts of Culicoides sonorensis inhibit murine lymphocyte proliferation and no production by macrophages. Am. J. Trop. Med. Hyg. 2006, 75, 532–536. [Google Scholar] [CrossRef] [PubMed]
- Limesand, K.H.; Higgs, S.; Pearson, L.D.; Beaty, B.J. Effect of mosquito salivary gland treatment on vesicular stomatitis New Jersey virus replication and interferon alpha/beta expression in vitro. J. Med. Entomol. 2003, 40, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Boorman, J. The maintenance of laboratory colonies of Culicoides variipennis (Coq.), C. nubeculosus (Mg.) and C. riethi Kieff. (Diptera, Ceratopogonidae). Bull. Entomol. Res. 1974, 64, 371–377. [Google Scholar] [CrossRef]
- Veronesi, E.; Antony, F.; Gubbins, S.; Golding, N.; Blackwell, A.; Mertens, P.P.; Brownlie, J.; Darpel, K.E.; Mellor, P.S.; Carpenter, S. Measurement of the infection and dissemination of bluetongue virus in Culicoides biting midges using a semi-quantitative rt-PCR assay and isolation of infectious virus. PLoS ONE 2013, 8, e70800. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Evermann, J.F.; McKeirnan, A.J.; Wilbur, L.A.; Levings, R.L.; Trueblood, E.S.; Baldwin, T.J.; Hughbanks, F.G. Canine fatalities associated with the use of a modified live vaccine administered during late stages of pregnancy. J. Vet. Diagn. Investig. 1994, 6, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Wilbur, L.A.; Evermann, J.F.; Levings, R.L.; Stoll, I.R.; Starling, D.E.; Spillers, C.A.; Gustafson, G.A.; McKeirnan, A.J. Abortion and death in pregnant bitches associated with a canine vaccine contaminated with bluetongue virus. J. Am. Vet. Med. Assoc. 1994, 204, 1762–1765. [Google Scholar]
- Bréard, E.; Belbis, G.; Hamers, C.; Moulin, V.; Lilin, T.; Moreau, F.; Millemann, Y.; Montange, C.; Sailleau, C.; Durand, B.; et al. Evaluation of humoral response and protective efficacy of two inactivated vaccines against bluetongue virus after vaccination of goats. Vaccine 2011, 29, 2495–2502. [Google Scholar] [CrossRef]
- Eschbaumer, M.; Wackerlin, R.; Savini, G.; Zientara, S.; Sailleau, C.; Bréard, E.; Beer, M.; Hoffmann, B. Contamination in bluetongue virus challenge experiments. Vaccine 2011, 29, 4299–4301. [Google Scholar] [CrossRef]
- Rasmussen, L.D.; Savini, G.; Lorusso, A.; Bellacicco, A.; Palmarini, M.; Caporale, M.; Rasmussen, T.B.; Belsham, G.J.; Botner, A. Transplacental transmission of field and rescued strains of BTV-2 and BTV-8 in experimentally infected sheep. Vet. Res. 2013, 44, 75. [Google Scholar] [CrossRef]
- Vandenbussche, F.; Sailleau, C.; Rosseel, T.; Desprat, A.; Viarouge, C.; Richardson, J.; Eschbaumer, M.; Hoffmann, B.; De Clercq, K.; Bréard, E.; et al. Full-Genome Sequencing of Four Bluetongue Virus Serotype 11 Viruses. Transbound. Emerg. Dis. 2015, 62, 565–571. [Google Scholar] [CrossRef] [PubMed]
- van Rijn, P.A.; Geurts, Y.; van der Spek, A.N.; Veldman, D.; van Gennip, R.G. Bluetongue virus serotype 6 in Europe in 2008-Emergence and disappearance of an unexpected non-virulent BTV. Vet. Microbiol. 2012, 158, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Orlowska, A.; Trebas, P.; Smreczak, M.; Marzec, A.; Zmudzinski, J.F. First detection of bluetongue virus serotype 14 in Poland. Arch. Virol. 2016, 161, 1969–1972. [Google Scholar] [CrossRef] [PubMed]
- Gilbert-Barness, E. Teratogenic causes of malformations. Ann. Clin. Lab. Sci. 2010, 40, 99–114. [Google Scholar] [PubMed]
- MacLachlan, N.J.; Conley, A.J.; Kennedy, P.C. Bluetongue and equine viral arteritis viruses as models of virus-induced fetal injury and abortion. Anim. Reprod. Sci. 2000, 60–61, 643–651. [Google Scholar] [CrossRef]
- Maclachlan, N.J.; Osburn, B.I. Teratogenic bluetongue and related orbivirus infections in pregnant ruminant livestock: Timing and pathogen genetics are critical. Curr. Opin. Virol. 2017, 27, 31–35. [Google Scholar] [CrossRef]
- Doceul, V.; Lara, E.; Sailleau, C.; Belbis, G.; Richardson, J.; Bréard, E.; Viarouge, C.; Dominguez, M.; Hendrikx, P.; Calavas, D.; et al. Epidemiology, molecular virology and diagnostics of Schmallenberg virus, an emerging orthobunyavirus in Europe. Vet. Res. 2013, 44, 31. [Google Scholar] [CrossRef]
- Garigliany, M.M.; Bayrou, C.; Kleijnen, D.; Cassart, D.; Desmecht, D. Schmallenberg virus in domestic cattle, Belgium, 2012. Emerg. Infect. Dis. 2012, 18, 1512–1514. [Google Scholar] [CrossRef]
- Parsonson, I.M.; McPhee, D.A.; Della-Porta, A.J.; McClure, S.; McCullagh, P. Transmission of Akabane virus from the ewe to the early fetus (32 to 53 days). J. Comp. Pathol. 1988, 99, 215–227. [Google Scholar] [CrossRef]
- Charles, J.A. Akabane virus. Vet. Clin. N. Am. 1994, 10, 525–546. [Google Scholar] [CrossRef]
- King, G.J.; Atkinson, B.A.; Robertson, H.A. Development of the bovine placentome during the second month of gestation. Reproduction 1979, 55, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Assis Neto, A.C.; Pereira, F.T.; Santos, T.C.; Ambrosio, C.E.; Leiser, R.; Miglino, M.A. Morpho-physical recording of bovine conceptus (Bos indicus) and placenta from days 20 to 70 of pregnancy. Reprod. Domest. Anim. 2010, 45, 760–772. [Google Scholar] [CrossRef] [PubMed]
- Spencer, T.E.; Johnson, G.A.; Bazer, F.W.; Burghardt, R.C. Implantation mechanisms: Insights from the sheep. Reproduction 2004, 128, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Bryden, M.M.; Evans, H.E.; Binns, W. Embryology of the sheep. I. Extraembryonic membranes and the development of body form. J. Morphol. 1972, 138, 169–185. [Google Scholar] [CrossRef] [PubMed]
- Evans, H.E.; Sack, W.O. Prenatal development of domestic and laboratory mammals: Growth curves, external features and selected references. Anat. Histol. Embryol. 1973, 2, 11–45. [Google Scholar] [CrossRef]
- Jordan, R.K. Development of sheep thymus in relation to in utero thymectomy experiments. Eur. J. Immunol. 1976, 6, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Khaksary-Mahabady, M.; Khazaeel, K.; Pourmahdi Borujeni, M.; Yazdanjoo, B. Morphometric development of sheep (Ovis Aries) lymph nodes in fetal period. Vet. Res. Forum 2018, 9, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Maddox, J.F.; Mackay, C.R.; Brandon, M.R. Ontogeny of ovine lymphocytes. II. An immunohistological study on the development of T lymphocytes in the sheep fetal spleen. Immunology 1987, 62, 107–112. [Google Scholar] [PubMed]
- Szabo, K.T. Congenital Malformations in Laboratory and Farm Animals; Academic Press: Cambridge, MA, USA, 1989. [Google Scholar]
- Coppock, R.W.; Dziwenka, M.M. Chapter 72—Teratogenesis in Livestock. In Reproductive and Developmental Toxicology, 2nd ed.; Elsevier: Atlanta, GA, USA, 2017; pp. 1391–1408. [Google Scholar] [CrossRef]
- Schultz, R.D.; Dunne, H.W.; Heist, C.E. Ontogeny of the bovine immune response. Infect. Immun. 1973, 7, 981–991. [Google Scholar]
- Silverstein, A.M.; Uhr, J.W.; Kraner, K.L.; Lukes, R.J. Fetal response to antigenic stimulus. II. Antibody production by the fetal lamb. J. Exp. Med. 1963, 117, 799–812. [Google Scholar] [CrossRef]
- Fahey, K.J.; Morris, B. Humoral immune responses in foetal sheep. Immunology 1978, 35, 651–661. [Google Scholar] [PubMed]
- De Clercq, K.; De Leeuw, I.; Verheyden, B.; Vandemeulebroucke, E.; Vanbinst, T.; Herr, C.; Meroc, E.; Bertels, G.; Steurbaut, N.; Miry, C.; et al. Transplacental infection and apparently immunotolerance induced by a wild-type bluetongue virus serotype 8 natural infection. Transbound. Emerg. Dis. 2008, 55, 352–359. [Google Scholar] [CrossRef] [PubMed]
- De Regge, N.; van den Berg, T.; Georges, L.; Cay, B. Diagnosis of Schmallenberg virus infection in malformed lambs and calves and first indications for virus clearance in the fetus. Vet. Microbiol. 2013, 162, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Maclachlan, N.J.; Drew, C.P.; Darpel, K.E.; Worwa, G. The pathology and pathogenesis of bluetongue. J. Comp. Pathol. 2009, 141, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, P.; Stokstad, M.; Myrmel, M.; Mutowembwa, P.; Loken, T.; Venter, E.H.; Van Vuuren, M. Transplacental infection in goats experimentally infected with a European strain of bluetongue virus serotype 8. Vet. J. 2013, 197, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Poskin, A.; Martinelle, L.; Van der Stede, Y.; Saegerman, C.; Cay, A.B.; De Regge, N. Genetically stable infectious Schmallenberg virus persists in foetal envelopes of pregnant ewes. J. Gen. Virol. 2017, 98, 1630–1635. [Google Scholar] [CrossRef]
- Consortium, E. Schmallenberg Virus Technical and Scientific Studies Final Report; Central Veterinary Institute: Wageningen, The Netherlands, 2014; p. 67. [Google Scholar]
- Santman-Berends, I.M.; van Wuijckhuise, L.; Vellema, P.; van Rijn, P.A. Vertical transmission of bluetongue virus serotype 8 virus in Dutch dairy herds in 2007. Vet. Microbiol. 2010, 141, 31–35. [Google Scholar] [CrossRef] [PubMed][Green Version]
- van Wuijckhuise, L.; Vellema, P.; Pelgrim, W.; Tolboom, R.; Hans, M.; van Rijn, P. Bluetongue virus serotype 8 in healthy young calves. Tijdschr. Diergeneeskd. 2008, 133, 992–994. [Google Scholar]
- Darpel, K.E.; Batten, C.A.; Veronesi, E.; Williamson, S.; Anderson, P.; Dennison, M.; Clifford, S.; Smith, C.; Philips, L.; Bidewell, C.; et al. Transplacental transmission of bluetongue virus 8 in cattle, UK. Emerg. Infect. Dis. 2009, 15, 2025–2028. [Google Scholar] [CrossRef]
- Desmecht, D.; Bergh, R.V.; Sartelet, A.; Leclerc, M.; Mignot, C.; Misse, F.; Sudraud, C.; Berthemin, S.; Jolly, S.; Mousset, B.; et al. Evidence for transplacental transmission of the current wild-type strain of bluetongue virus serotype 8 in cattle. Vet. Rec. 2008, 163, 50–52. [Google Scholar] [CrossRef]
- Kirkland, P.D.; Hawkes, R.A. A comparison of laboratory and ‘wild’ strains of bluetongue virus--is there any difference and does it matter? Vet. Ital. 2004, 40, 448–455. [Google Scholar] [PubMed]
- Osburn, B.I.; Johnson, R.T.; Silverstein, A.M.; Prendergast, R.A.; Jochim, M.M.; Levy, S.E. Experimental viral-induced congenital encephalopathies. II. The pathogenesis of bluetongue vaccine virus infection in fetal lambs. Lab. Investig. 1971, 25, 206–210. [Google Scholar] [PubMed]
- Anderson, C.K.; Jensen, R. Pathologic changes in placentas of ewes inoculated with bluetongue virus. Am. J. Vet. Res. 1969, 30, 987–989. [Google Scholar] [PubMed]
- Belbis, G.; Bréard, E.; Cordonnier, N.; Moulin, V.; Desprat, A.; Sailleau, C.; Viarouge, C.; Doceul, V.; Zientara, S.; Millemann, Y. Evidence of transplacental transmission of bluetongue virus serotype 8 in goats. Vet. Microbiol. 2013, 166, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, P.D.; Barry, R.D.; Harper, P.A.; Zelski, R.Z. The development of Akabane virus-induced congenital abnormalities in cattle. Vet. Rec. 1988, 122, 582–586. [Google Scholar] [CrossRef] [PubMed]
- Saegerman, C.; Bolkaerts, B.; Baricalla, C.; Raes, M.; Wiggers, L.; de Leeuw, I.; Vandenbussche, F.; Zimmer, J.Y.; Haubruge, E.; Cassart, D.; et al. The impact of naturally-occurring, trans-placental bluetongue virus serotype-8 infection on reproductive performance in sheep. Vet. J. 2011, 187, 72–80. [Google Scholar] [CrossRef] [PubMed]
- MacLachlan, N.J.; Osburn, B.I.; Stott, J.L.; Ghalib, H.W. Orbivirus infection of the bovine fetus. Prog. Clin. Boil. Res. 1985, 178, 79–84. [Google Scholar]
- Vercauteren, G.; Miry, C.; Vandenbussche, F.; Ducatelle, R.; Van der Heyden, S.; Vandemeulebroucke, E.; De Leeuw, I.; Deprez, P.; Chiers, K.; De Clercq, K. Bluetongue virus serotype 8-associated congenital hydranencephaly in calves. Transbound. Emerg. Dis. 2008, 55, 293–298. [Google Scholar] [CrossRef]
- Richardson, C.; Taylor, W.P.; Terlecki, S.; Gibbs, E.P. Observations on transplacental infection with bluetongue virus in sheep. Am. J. Vet. Res. 1985, 46, 1912–1922. [Google Scholar]
- Savini, G.; Lorusso, A.; Paladini, C.; Migliaccio, P.; Di Gennaro, A.; Di Provvido, A.; Scacchia, M.; Monaco, F. Bluetongue serotype 2 and 9 modified live vaccine viruses as causative agents of abortion in livestock: A retrospective analysis in Italy. Transbound. Emerg. Dis. 2014, 61, 69–74. [Google Scholar] [CrossRef]
- Waldvogel, A.S.; Anderson, G.A.; Phillips, D.L.; Osburn, B.I. Infection of bovine fetuses at 120 days’ gestation with virulent and avirulent strains of bluetongue virus serotype 11. Comp. Immunol. Microbiol. Infect. Dis. 1992, 15, 53–63. [Google Scholar] [CrossRef]
- McKercher, D.G.; Saito, J.K.; Singh, K.V. Serologic evidence of an etiologic role for bluetongue virus in hydranencephaly of calves. J. Am. Vet. Med. Assoc. 1970, 156, 1044–1047. [Google Scholar] [PubMed]
- Wernike, K.; Elbers, A.; Beer, M. Schmallenberg virus infection. Rev. Sci. Tech. 2015, 34, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Veldhuis, A.M.; Carp-van Dijken, S.; van Wuijckhuise, L.; Witteveen, G.; van Schaik, G. Schmallenberg virus in Dutch dairy herds: Potential risk factors for high within-herd seroprevalence and malformations in calves, and its impact on productivity. Vet. Microbiol. 2014, 168, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, M.; Gache, K.; Touratier, A.; Perrin, J.B.; Fediaevsky, A.; Collin, E.; Bréard, E.; Sailleau, C.; Viarouge, C.; Zanella, G.; et al. Spread and impact of the Schmallenberg virus epidemic in France in 2012–2013. BMC Vet. Res. 2014, 10, 248. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, M.; Hendrikx, P.; Zientara, S.; Calavas, D.; Jay, M.; Touratier, A.; Languille, J.; Fediaevsky, A. Preliminary estimate of Schmallenberg virus infection impact in sheep flocks—France. Vet. Rec. 2012, 171, 426. [Google Scholar] [CrossRef] [PubMed]
- Saegerman, C.; Martinelle, L.; Dal Pozzo, F.; Kirschvink, N. Preliminary survey on the impact of Schmallenberg virus on sheep flocks in South of Belgium. Transbound. Emerg. Dis. 2014, 61, 469–472. [Google Scholar] [CrossRef]
- Agerholm, J.S.; Hewicker-Trautwein, M.; Peperkamp, K.; Windsor, P.A. Virus-induced congenital malformations in cattle. Acta Vet. Scand. 2015, 57, 54. [Google Scholar] [CrossRef]
- Martinelle, L.; Dal Pozzo, F.; Kirschvink, N.; De la Grandiere, M.A.; Thiry, E.; Saegerman, C. Le virus Schmallenberg ou l’émergence du premier Orthobunyavirus du sérogroupe Simbu en Europe. Ann. Med. Vet. 2012, 156, 7–24. [Google Scholar]
- Afonso, A.; Abrahantes, J.C.; Conraths, F.; Veldhuis, A.; Elbers, A.; Roberts, H.; Van der Stede, Y.; Meroc, E.; Gache, K.; Richardson, J. The Schmallenberg virus epidemic in Europe-2011-2013. Prev. Vet. Med. 2014, 116, 391–403. [Google Scholar] [CrossRef]
- Dixon, A.B.; Knights, M.; Winkler, J.L.; Marsh, D.J.; Pate, J.L.; Wilson, M.E.; Dailey, R.A.; Seidel, G.; Inskeep, E.K. Patterns of late embryonic and fetal mortality and association with several factors in sheep. J. Anim. Sci. 2007, 85, 1274–1284. [Google Scholar] [CrossRef] [PubMed]
- Diskin, M.G.; Morris, D.G. Embryonic and early foetal losses in cattle and other ruminants. Reprod. Domest. Anim. 2008, 43 (Suppl. 2), 260–267. [Google Scholar] [CrossRef] [PubMed]
- Geary, T. Management Strategies to Reduce Embryonic Loss. In Proceedings of the Range Beef Cow Symposium, Miles City, MT, USA, 6–8 December 2005. [Google Scholar]
- Nusinovici, S.; Seegers, H.; Joly, A.; Beaudeau, F.; Fourichon, C. Quantification and at-risk period of decreased fertility associated with exposure to bluetongue virus serotype 8 in naive dairy herds. J. Dairy Sci. 2012, 95, 3008–3020. [Google Scholar] [CrossRef] [PubMed]
- Santman-Berends, I.M.; Hage, J.J.; van Rijn, P.A.; Stegeman, J.A.; van Schaik, G. Bluetongue virus serotype 8 (BTV-8) infection reduces fertility of Dutch dairy cattle and is vertically transmitted to offspring. Theriogenology 2010, 74, 1377–1384. [Google Scholar] [CrossRef] [PubMed]
- Lechner, I.; Wuthrich, M.; Meylan, M.; van den Borne, B.H.P.; Schupbach-Regula, G. Association of clinical signs after acute Schmallenberg virus infection with milk production and fertility in Swiss dairy cows. Prev. Vet. Med. 2017, 146, 121–129. [Google Scholar] [CrossRef]
- Belgian Biosafety Server. Available online: https://www.biosafety.be/content/tools-belgian-classification-micro-organisms-based-their-biological-risks (accessed on 15 August 2018).
- Spencer, T.E.; Forde, N.; Lonergan, P. Insights into conceptus elongation and establishment of pregnancy in ruminants. Reprod. Fertil. Dev. 2016, 29, 84–100. [Google Scholar] [CrossRef]


{kind=link}
| Virus | Type of Inoculum | No. and Species of Infected Animals | Cell Type | Number of Passages | Inoculation Route | Volume (mL) | Doses (TCID50/mL) | Reference |
|---|---|---|---|---|---|---|---|---|
| BTV8 | Cell-passaged | 16 sheep | KC | 2 | SC | 1 | 105,75 | Flannery et al., 2019 [12] |
| BTV4 TV16 | Cell-passaged | 6 sheep | BHK21+KC | not specified | ID | 1 | 106 | Putty et al., 2019 [13] |
| BTV1 BTV2 BTV4 BTV9 BTV16 | Cell-passaged | 30 cattle | BHK-21 | 2-4 | SC | 2,5-4 | 106 | Martinelle et al., 2018 [14] |
| BTV27 | Cell-passaged blood (goat) | 7 sheep, 13 goats and 4 cattle | BSR; ECE+BSR | 3; 1+3 or 1+2 | SC; IV (blood) | 2, 3 or 4; 1 (blood) | 103–104.67 | Bréard et al., 2018 [10] |
| BTV4 | Cell-passaged | 4 sheep, 3 goats and 3 calves | KC+BHK-21 | 1+1 | SC | 2 – 4 | 106 | Schulz et al., 2018 [15] |
| BTV25 * | Reverse genetic | 10 sheep and 2 goats | / | / | SC+IV | 1 | 105 | van Rijn et al., 2016 [16] |
| BTV8 | Cell-passaged | 8 calves | BHK-21 | 2 | SC+IV | 1–4 | 104–106.15 | Martinelle et al., 2016 [17] |
| BTV8 | Cell-passaged | 10 sheep and 4 cattle | KC | 2 | SC, ID | 1 | 107 | Darpel et al., 2016 [18] |
| BTV8 | Blood | 8 sheep | / | / | ID | 2 | 106.08 | Drolet et al., 2015 [19] |
| BTV8 BTV16 | Cell-passaged | 37 sheep | KC | 3 and 2 | SC | 3 | not possible | Bréard et al., 2015 [20] |
| SBV | Cell-passaged | 13 cattle | BHK-21 | 4 | SC | 10 | 105 | Kęsik-Maliszewska et al., 2019 [21] |
| SBV | Serum (cattle) | 35 cattle | / | / | SC | 2 X 0.5 | not specified | König P et al., 2019 [22] |
| SBV | Cell-passaged/sheep brain homogenate/serum (sheep) | 10 sheep, 9 cattle | C6/36 | 1 | SC | 1 – 3 | 105.15 and 103.15** | Endalew et al., 2019 [23] |
| SBV | Serum (cattle) | 25 goats | / | / | SC | 1 | / | Laloy et al., 2017 [24] |
| SBV | Plasma | 9 sheep | / | / | IV | 20 | not specified | Rodríguez-Prieto et al., 2016 [25] |
| SBV | Serum (cattle) | 5 sheep | / | / | SC | 1 | 103.3 | Poskin et al., 2015 [26] |
| SBV | Serum (cattle) | 17 sheep | / | / | SC | 1 | 103.3 | Martinelle et al., 2015 [27] |
| SBV | Serum (cattle) | 9 sheep | / | / | SC, ID, IN | 1 | 103.3 | Martinelle et al., 2015 [28] |
| SBV | Serum (cattle), blood (sheep) | 6 goats | / | / | SC | 1 | not specified | Laloy et al., 2015 [29] |
| SBV | Serum (cattle) | 12 sheep | / | / | SC | 1 | 103.3, 102.3, 101.3 and 100.3 | Poskin et al., 2014 [30] |
| Event | Timing in Cow (dpc) | Timing in Sheep (dpc) |
|---|---|---|
| Blastocyst hatching from zona pellucida | 9 | 9 |
| Elongation of the blastocyst, establishment of the primitive streak, emergence of the notochord | 17–18 | 13–14 |
| Appearance of neural folds, closure of the neural groove | 17–19 | 15–16 |
| Implantation begins | 16–19 | 15–18 |
| Neurula | 20–21 | 17 |
| Neural tube complete; optic and otic vesicles present | 21–23 | 19–20 |
| Placentation begins | 22–23 | 17–22 |
| Three brain vesicles visible | 24–25 | 17 |
| Placentoma are detectable | 32–36 | 21 |
| Lymphoid development of the thymus | 42 | 36 |
| Spleen development | 55 | 43–44 |
| Peripheral lymph nodes | 60 | 45 |
| IgM containing cells | 59 | 65 |
| Myelin sheath acquisition (starting) | 60 | 54–63 |
| IgG containing cells | 145 | 87 |
| Lesion | Definition | BVDV | SBV | BTV | AKAV/AV |
|---|---|---|---|---|---|
| Hydranencephaly | Extensive loss of cerebral tissue with replacement by clear fluid | + | + | + | + |
| Porencephaly | Cystic fluid filled cavities in the brain tissue | + | + | + | + |
| Hydrocephalus | Dilation of the lateral ventricles by cerebrospinal fluid | + | + | + | - |
| Microencephaly | Reduced size of the cerebrum | + | + | + | + |
| Cerebellar hypoplasia | Reduced size of the cerebellum | + | + | + | |
| Kyphosis | Dorsal vertebral column curvature | - | + | - | - |
| Lordosis | Ventral vertebral column curvature | - | + | - | - |
| Scoliosis | Lateral vertebral column curvature | - | + | - | - |
| Torticollis | Twisted cervical vertebral column curvature | - | + | - | - |
| Arthrogryposis | Joint contraction of the limbs | - | + | +/- | + |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinelle, L.; Dal Pozzo, F.; Thiry, E.; De Clercq, K.; Saegerman, C. Reliable and Standardized Animal Models to Study the Pathogenesis of Bluetongue and Schmallenberg Viruses in Ruminant Natural Host Species with Special Emphasis on Placental Crossing. Viruses 2019, 11, 753. https://doi.org/10.3390/v11080753
Martinelle L, Dal Pozzo F, Thiry E, De Clercq K, Saegerman C. Reliable and Standardized Animal Models to Study the Pathogenesis of Bluetongue and Schmallenberg Viruses in Ruminant Natural Host Species with Special Emphasis on Placental Crossing. Viruses. 2019; 11(8):753. https://doi.org/10.3390/v11080753
Chicago/Turabian StyleMartinelle, Ludovic, Fabiana Dal Pozzo, Etienne Thiry, Kris De Clercq, and Claude Saegerman. 2019. "Reliable and Standardized Animal Models to Study the Pathogenesis of Bluetongue and Schmallenberg Viruses in Ruminant Natural Host Species with Special Emphasis on Placental Crossing" Viruses 11, no. 8: 753. https://doi.org/10.3390/v11080753
APA StyleMartinelle, L., Dal Pozzo, F., Thiry, E., De Clercq, K., & Saegerman, C. (2019). Reliable and Standardized Animal Models to Study the Pathogenesis of Bluetongue and Schmallenberg Viruses in Ruminant Natural Host Species with Special Emphasis on Placental Crossing. Viruses, 11(8), 753. https://doi.org/10.3390/v11080753