Current Findings on Gut Microbiota Mediated Immune Modulation against Viral Diseases in Chicken


Abstract
:1. Introduction
2. Avian Influenza Virus
3. Commensal Bacteria Elicit Immunity
4. AIV Mediated Dysbiosis in Commensal Microbiota
5. Infectious Bursal Disease Virus (IBDV)
6. Marek’s Disease Virus
7. Newcastle Disease Virus (NDV)
8. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ley, R.E.; Peterson, D.A.; Gordon, J.I. Ecological and Evolutionary Forces Shaping Microbial Diversity in the Human Intestine. Cell 2006, 124, 837–848. [Google Scholar] [CrossRef] [Green Version]
- Dethlefsen, L.; McFall-Ngai, M.; Relman, D.A. An ecological and evolutionary perspective on human–microbe mutualism and disease. Nature 2007, 449, 811. [Google Scholar] [CrossRef]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313. [Google Scholar] [CrossRef]
- Flint, H.J.; Scott, K.P.; Louis, P.; Duncan, S.H. The role of the gut microbiota in nutrition and health. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 577. [Google Scholar] [CrossRef]
- Zhu, G.; Jiang, Y.; Yao, Y.; Wu, N.; Luo, J.; Hu, M.; Tu, Y.; Xu, M. Ovotransferrin ameliorates the dysbiosis of immunomodulatory function and intestinal microbiota induced by cyclophosphamide. Food Funct. 2019, 10, 1109–1122. [Google Scholar] [CrossRef]
- Lu, S.; Zuo, T.; Zhang, N.; Shi, H.; Liu, F.; Wu, J.; Wang, Y.; Xue, C.; Tang, Q.-J. High throughput sequencing analysis reveals amelioration of intestinal dysbiosis by squid ink polysaccharide. J. Funct. Foods 2016, 20, 506–515. [Google Scholar] [CrossRef]
- Xie, Q.; Pan, M.; Huang, R.; Tian, X.; Tao, X.; Shah, N.P.; Wei, H.; Wan, C. Modulation of the small intestinal microbial community composition over short-term or long-term administration with Lactobacillus plantarum ZDY2013. J. Dairy Sci. 2016, 99, 6913–6921. [Google Scholar] [CrossRef]
- Kamada, N.; Seo, S.-U.; Chen, G.Y.; Núñez, G. Role of the gut microbiota in immunity and inflammatory disease. Nat. Rev. Immunol. 2013, 13, 321. [Google Scholar] [CrossRef]
- Hooper, L.V.; Gordon, J.I. Commensal host-bacterial relationships in the gut. Science 2001, 292, 1115–1118. [Google Scholar] [CrossRef]
- Macpherson, A.J.; Harris, N.L. Interactions between commensal intestinal bacteria and the immune system. Nat. Rev. Immunol. 2004, 4, 478. [Google Scholar] [CrossRef]
- Hooper, L.V.; Littman, D.R.; Macpherson, A.J. Interactions between the microbiota and the immune system. Science 2012, 336, 1268–1273. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.; McCoy, K.D.; Macpherson, A.J. Use of axenic animals in studying the adaptation of mammals to their commensal intestinal microbiota. Semin. Immunol. 2007, 19, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Mazmanian, S.K.; Round, J.L.; Kasper, D.L. A microbial symbiosis factor prevents intestinal inflammatory disease. Nature 2008, 453, 620. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, I.I.; Atarashi, K.; Manel, N.; Brodie, E.L.; Shima, T.; Karaoz, U.; Wei, D.; Goldfarb, K.C.; Santee, C.A.; Lynch, S.V. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell 2009, 139, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Atarashi, K.; Tanoue, T.; Shima, T.; Imaoka, A.; Kuwahara, T.; Momose, Y.; Cheng, G.; Yamasaki, S.; Saito, T.; Ohba, Y. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 2011, 331, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Yegani, M.; Korver, D. Factors affecting intestinal health in poultry. Poult. Sci. 2008, 87, 2052–2063. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Kim, G.; Cha, C. Spatial heterogeneity and stability of bacterial community in the gastrointestinal tracts of broiler chickens. Poult. Sci. 2014, 93, 1942–1950. [Google Scholar] [CrossRef]
- Sharma, J. The avian immune system. Dis. Poult. Saif. YmEd Iowa State Univ. PressAmes. 2003, 5–16. [Google Scholar]
- Spor, A.; Koren, O.; Ley, R. Unravelling the effects of the environment and host genotype on the gut microbiome. Nat. Rev. Microbiol. 2011, 9, 279. [Google Scholar] [CrossRef]
- Chang, C.; Lin, H. Dysbiosis in gastrointestinal disorders. Best Pract. Res. Clin. Gastroenterol. 2016, 30, 3–15. [Google Scholar] [CrossRef]
- Deriu, E.; Boxx, G.M.; He, X.; Pan, C.; Benavidez, S.D.; Cen, L.; Rozengurt, N.; Shi, W.; Cheng, G. Influenza virus affects intestinal microbiota and secondary salmonella infection in the gut through type I interferons. PLoS Pathog. 2016, 12, e1005572. [Google Scholar] [CrossRef] [PubMed]
- Winter, S.E.; Winter, M.G.; Xavier, M.N.; Thiennimitr, P.; Poon, V.; Keestra, A.M.; Laughlin, R.C.; Gomez, G.; Wu, J.; Lawhon, S.D. Host-derived nitrate boosts growth of E. coli in the inflamed gut. Science 2013, 339, 708–711. [Google Scholar] [CrossRef] [PubMed]
- Barnes, E.M. The intestinal microflora of poultry and game birds during life and after storage. J. Appl. Bacteriol. 1979, 46, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Si, W.; Forster, R.J.; Huang, R.; Yu, H.; Yin, Y.; Yang, C.; Han, Y. 16S rRNA gene-based analysis of mucosa-associated bacterial community and phylogeny in the chicken gastrointestinal tracts: From crops to ceca. Fems Microbiol. Ecol. 2007, 59, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Wang, Q.; Li, H.; Xu, C.; Cui, N.; Zhao, X. 16S rRNA genes Illumina sequencing revealed differential cecal microbiome in specific pathogen free chickens infected with different subgroup of avian leukosis viruses. Vet. Microbiol. 2017, 207, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Stanley, D.; Wu, S.-B.; Rodgers, N.; Swick, R.A.; Moore, R.J. Differential responses of cecal microbiota to fishmeal, Eimeria and Clostridium perfringens in a necrotic enteritis challenge model in chickens. PLoS ONE 2014, 9, e104739. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-J.; Wu, G.-H.; Kuo, R.-L.; Shih, S.-R. Role of the intestinal microbiota in the immunomodulation of influenza virus infection. Microbes Infect. 2017, 19, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Budden, K.F.; Gellatly, S.L.; Wood, D.L.; Cooper, M.A.; Morrison, M.; Hugenholtz, P.; Hansbro, P.M. Emerging pathogenic links between microbiota and the gut–lung axis. Nat. Rev. Microbiol. 2017, 15, 55. [Google Scholar] [CrossRef] [PubMed]
- Thorburn, A.N.; McKenzie, C.I.; Shen, S.; Stanley, D.; Macia, L.; Mason, L.J.; Roberts, L.K.; Wong, C.H.; Shim, R.; Robert, R. Evidence that asthma is a developmental origin disease influenced by maternal diet and bacterial metabolites. Nat. Commun. 2015, 6, 7320. [Google Scholar] [CrossRef]
- Ichinohe, T.; Pang, I.K.; Kumamoto, Y.; Peaper, D.R.; Ho, J.H.; Murray, T.S.; Iwasaki, A. Microbiota regulates immune defense against respiratory tract influenza A virus infection. Proc. Natl. Acad. Sci. USA 2011, 108, 5354–5359. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.Z.; Ravindran, R.; Chassaing, B.; Carvalho, F.A.; Maddur, M.S.; Bower, M.; Hakimpour, P.; Gill, K.P.; Nakaya, H.I.; Yarovinsky, F. TLR5-mediated sensing of gut microbiota is necessary for antibody responses to seasonal influenza vaccination. Immunity 2014, 41, 478–492. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Jiang, Z.-Y.; Sun, Y.-F.; Yu, B.; Chen, J.; Dai, C.-Q.; Wu, X.-L.; Tang, X.-L.; Chen, X.-Y. Microbiota regulates the TLR7 signaling pathway against respiratory tract influenza A virus infection. Curr. Microbiol. 2013, 67, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Clarke, T.B. Early innate immunity to bacterial infection in the lung is regulated systemically by the commensal microbiota via nod-like receptor ligands. Infect. Immun. 2014, 82, 4596–4606. [Google Scholar] [CrossRef] [PubMed]
- Fagundes, C.T.; Amaral, F.A.; Vieira, A.T.; Soares, A.C.; Pinho, V.; Nicoli, J.R.; Vieira, L.Q.; Teixeira, M.M.; Souza, D.G. Transient TLR activation restores inflammatory response and ability to control pulmonary bacterial infection in germfree mice. J. Immunol. 2012, 188, 1411–1420. [Google Scholar] [CrossRef] [PubMed]
- Schuijt, T.J.; Lankelma, J.M.; Scicluna, B.P.; e Melo, F.d.S.; Roelofs, J.J.; de Boer, J.D.; Hoogendijk, A.J.; de Beer, R.; de Vos, A.; Belzer, C. The gut microbiota plays a protective role in the host defence against pneumococcal pneumonia. Gut 2016, 65, 575–583. [Google Scholar] [CrossRef] [PubMed]
- McAleer, J.P.; Nguyen, N.L.; Chen, K.; Kumar, P.; Ricks, D.M.; Binnie, M.; Armentrout, R.A.; Pociask, D.A.; Hein, A.; Yu, A. Pulmonary Th17 antifungal immunity is regulated by the gut microbiome. J. Immunol. 2016, 197, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, J.K.; Virgin, H.W. Transkingdom control of viral infection and immunity in the mammalian intestine. Science 2016, 351, aad5872. [Google Scholar] [CrossRef]
- Samuelson, D.R.; Welsh, D.A.; Shellito, J.E. Regulation of lung immunity and host defense by the intestinal microbiota. Front. Microbiol. 2015, 6, 1085. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Wei, T.; Tianv, H.; Cheng, J.; Xiao, J.; Wang, M.; Hu, Y. Small intestinal injury in mice infected with respiratory influenza A virus: Evidence for virus induced gastroenteritis. Biotechnol. Lett. 2015, 37, 1585–1592. [Google Scholar] [CrossRef]
- Maeda, N.; Nakamura, R.; Hirose, Y.; Murosaki, S.; Yamamoto, Y.; Kase, T.; Yoshikai, Y. Oral administration of heat-killed Lactobacillus plantarum L-137 enhances protection against influenza virus infection by stimulation of type I interferon production in mice. Int. Immunopharmacol. 2009, 9, 1122–1125. [Google Scholar] [CrossRef]
- Yitbarek, A.; Taha-Abdelaziz, K.; Hodgins, D.C.; Read, L.; Nagy, É.; Weese, J.S.; Caswell, J.L.; Parkinson, J.; Sharif, S. Gut microbiota-mediated protection against influenza virus subtype H9N2 in chickens is associated with modulation of the innate responses. Sci. Rep. 2018, 8, 13189. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, Y.; Moriya, T.; Sakai, F.; Ikeda, N.; Shiozaki, T.; Hosoya, T.; Nakagawa, H.; Miyazaki, T. Oral administration of Lactobacillus gasseri SBT2055 is effective for preventing influenza in mice. Sci. Rep. 2014, 4, 4638. [Google Scholar] [CrossRef] [Green Version]
- Sakai, F.; Hosoya, T.; Ono-Ohmachi, A.; Ukibe, K.; Ogawa, A.; Moriya, T.; Kadooka, Y.; Shiozaki, T.; Nakagawa, H.; Nakayama, Y. Lactobacillus gasseri SBT2055 induces TGF-β expression in dendritic cells and activates TLR2 signal to produce IgA in the small intestine. PLoS ONE 2014, 9, e105370. [Google Scholar] [CrossRef] [PubMed]
- Nishihira, J.; Nishimura, M.; Moriya, T.; Sakai, F.; Kabuki, T.; Kawasaki, Y. Lactobacillus Gasseri Potentiates Immune Response Against Influenza Virus Infection. In Immunity and Inflammation in Health and Disease; Elsevier: Amsterdam, The Netherland, 2018; pp. 249–255. [Google Scholar]
- Denny, J.E.; Powell, W.L.; Schmidt, N.W. Local and long-distance calling: Conversations between the gut microbiota and intra-and extra-gastrointestinal tract infections. Front. Cell. Infect. Microbiol. 2016, 6, 41. [Google Scholar] [CrossRef] [PubMed]
- Dunn, J.R.; Gimeno, I.M. Current status of Marek’s disease in the United States and worldwide based on a questionnaire survey. Avian Dis. 2013, 57, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Couteaudier, M.; Denesvre, C. Marek’s disease virus and skin interactions. Vet. Res. 2014, 45, 36. [Google Scholar] [CrossRef] [PubMed]
- Witter, R.; Schat, K. Diseases of Poultry, Chapter 15: Subchapter—Marek’s Disease. In Disease of Poltry; Agricultural Research Service U.S. Department of Agriculture: Urbana, IL, USA, 2003; pp. 407–465. [Google Scholar]
- Calnek, B.; Schat, K.; Ross, L.; Chen, C.L. Further characterization of marek’s disease virus-infected lymphocytes. II. In vitro infection. Int. J. Cancer 1984, 33, 399–406. [Google Scholar] [CrossRef]
- Read, A.F.; Baigent, S.J.; Powers, C.; Kgosana, L.B.; Blackwell, L.; Smith, L.P.; Kennedy, D.A.; Walkden-Brown, S.W.; Nair, V.K. Imperfect vaccination can enhance the transmission of highly virulent pathogens. PLoS Biol. 2015, 13, e1002198. [Google Scholar] [CrossRef]
- Witter, R. Increased virulence of Marek’s disease virus field isolates. Avian Dis. 1997, 41, 149–163. [Google Scholar] [CrossRef]
- Nair, V. Evolution of Marek’s disease–a paradigm for incessant race between the pathogen and the host. Vet. J. 2005, 170, 175–183. [Google Scholar] [CrossRef]
- Kamaldeep, P.; Sharma, C.; Narang, G. Occurrence of Marek’s disease in vaccinated poultry flocks of Haryana (India). Int. J. Poult. Sci. 2007, 6, 372–377. [Google Scholar]
- Arulmozhi, A.; Saravanan, S.; Mohan, B.; Balasubramaniam, G. Marek’s disease in vaccinated poultry flocks in and around Namakkal region of Tamil Nadu. Indian J. Vet. Pathol. 2011, 35, 45–47. [Google Scholar]
- Muniyellappa, H.; Satyanarayana, M.; Isloor, S.; Gowda, N.S. Marek’s disease outbreak among vaccinated commercial layer flocks in the mining area of Karnataka, India. Vet. Rec. 2013, 172, 452. [Google Scholar] [CrossRef] [PubMed]
- McPherson, M.; Delany, M. Virus and host genomic, molecular, and cellular interactions during Marek’s disease pathogenesis and oncogenesis. Poult. Sci. 2016, 95, 412–429. [Google Scholar] [CrossRef] [PubMed]
- Nair, V. Latency and tumorigenesis in Marek’s disease. Avian Dis. 2013, 57, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Lillehoj, H.S.; Trout, J.M. Avian gut-associated lymphoid tissues and intestinal immune responses to Eimeria parasites. Clin. Microbiol. Rev. 1996, 9, 349–360. [Google Scholar] [CrossRef]
- Arega, A.M. Review on infectious Bursal disease: Threat for Ethiopian poultry industry. IJALS 2018, 11, 52–65. [Google Scholar] [CrossRef]
- Khan, R.S.A.; Sajid, S.; Habib, M.; Ali, W.; Shah, M.S.-u.-D.; Sarfraz, M. History of Gumboro (infectious bursal disease) in Pakistan. Saudi Pharm. J. 2017, 25, 453–459. [Google Scholar] [CrossRef]
- Ingrao, F.; Rauw, F.; Lambrecht, B.; van den Berg, T. Infectious bursal disease: A complex host–pathogen interaction. Dev. Comp. Immunol. 2013, 41, 429–438. [Google Scholar] [CrossRef]
- Giambrone, J.; Donahoe, J.; Dawe, D.; Eidson, C. Specific suppression of the bursa-dependent immune system of chicks with infectious bursal disease virus. Am. J. Vet. Res. 1977, 38, 581–583. [Google Scholar]
- Delmas, B.; Kibenge, F.S.B.; Leon, J.C.; Mundt, E.; Vakaharia, V.N.; Wu, J.L. Birnaviridae. In Virus Virus Taxonomy. 8th Report ICTV; Fauquet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U., Ball, L.A., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2005; pp. 561–569. [Google Scholar]
- Brown, M.D.; Skinner, M.A. Coding sequences of both genome segments of a European ‘very virulent’infectious bursal disease virus. Virus Res. 1996, 40, 1–15. [Google Scholar] [CrossRef]
- Müller, H.; Islam, M.R.; Raue, R. Research on infectious bursal disease—the past, the present and the future. Vet. Microbiol. 2003, 97, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Wang, Y.; Zhang, E.; Han, X.; Yu, Z.; Liu, H. VP1 and VP3 are required and sufficient for translation initiation of uncapped infectious bursal disease virus genomic double-stranded RNA. J. Virol. 2018, 92, e01345-17. [Google Scholar] [CrossRef] [PubMed]
- Hoerr, F.J. Clinical aspects of immunosuppression in poultry. Avian Dis. 2010, 54, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Balamurugan, V.; Kataria, J. Economically important non-oncogenic immunosuppressive viral diseases of chicken—current status. Vet. Res. Commun. 2006, 30, 541–566. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, D.J. Advances in vaccine research against economically important viral diseases of food animals: Infectious bursal disease virus. Vet. Microbiol. 2017, 206, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Käufer, I.; Weiss, E. Significance of bursa of Fabricius as target organ in infectious bursal disease of chickens. Infect. Immun. 1980, 27, 364–367. [Google Scholar] [PubMed]
- Turmagambetova, A.S.; Alexyuk, M.S.; Bogoyavlenskiy, A.P.; Linster, M.; Alexyuk, P.G.; Zaitceva, I.A.; Smith, G.J.; Berezin, V.E. Monitoring of Newcastle disease virus in environmental samples. Arch. Virol. 2017, 162, 2843–2846. [Google Scholar] [CrossRef] [PubMed]
- Maes, P.; Amarasinghe, G.K.; Ayllón, M.A.; Basler, C.F.; Bavari, S.; Blasdell, K.R.; Briese, T.; Brown, P.A.; Bukreyev, A.; Balkema-Buschmann, A. Taxonomy of the order Mononegavirales: Second update 2018. Arch. Virol. 2019, 164, 1233–1244. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.P. ERIC-PCR Based Fingerprint to Analyze the Structural Features of Intestinal Microbial Communities of Chicks Infected with Salmonella gallinarum, Newcastle Disease Virus and the Healthy Chicks. Ph.D. Thesis, Sichuan Agricultural University, Sichuan, China, 2007. [Google Scholar]
- Post, J.; Burt, D.W.; Cornelissen, J.B.; Broks, V.; van Zoelen, D.; Peeters, B.; Rebel, J.M. Systemic virus distribution and host responses in brain and intestine of chickens infected with low pathogenic or high pathogenic avian influenza virus. Virol. J. 2012, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- Abt, M.C.; Osborne, L.C.; Monticelli, L.A.; Doering, T.A.; Alenghat, T.; Sonnenberg, G.F.; Paley, M.A.; Antenus, M.; Williams, K.L.; Erikson, J. Commensal bacteria calibrate the activation threshold of innate antiviral immunity. Immunity 2012, 37, 158–170. [Google Scholar] [CrossRef] [PubMed]
- Yitbarek, A.; Alkie, T.; Taha-Abdelaziz, K.; Astill, J.; Rodriguez-Lecompte, J.; Parkinson, J.; Nagy, É.; Sharif, S. Gut microbiota modulates type I interferon and antibody-mediated immune responses in chickens infected with influenza virus subtype H9N2. Benef. Microbes 2018, 9, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; Chen, Z.J. Intrinsic antiviral immunity. Nat. Immunol. 2012, 13, 214. [Google Scholar] [CrossRef] [PubMed]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O’garra, A. Type I interferons in infectious disease. Nat. Rev. Immunol. 2015, 15, 87. [Google Scholar] [CrossRef] [PubMed]
- Barjesteh, N.; Shojadoost, B.; Brisbin, J.T.; Emam, M.; Hodgins, D.C.; Nagy, É.; Sharif, S. Reduction of avian influenza virus shedding by administration of Toll-like receptor ligands to chickens. Vaccine 2015, 33, 4843–4849. [Google Scholar] [CrossRef]
- Hernández, P.P.; Mahlakõiv, T.; Yang, I.; Schwierzeck, V.; Nguyen, N.; Guendel, F.; Gronke, K.; Ryffel, B.; Hölscher, C.; Dumoutier, L. Interferon-λ and interleukin 22 act synergistically for the induction of interferon-stimulated genes and control of rotavirus infection. Nat. Immunol. 2015, 16, 698. [Google Scholar] [CrossRef] [PubMed]
- Gimeno Brias, S.; Stack, G.; Stacey, M.A.; Redwood, A.J.; Humphreys, I.R. The role of IL-22 in viral infections: Paradigms and paradoxes. Front. Immunol. 2016, 7, 211. [Google Scholar] [CrossRef]
- Yuk, S.-S.; Lee, D.-H.; Park, J.-K.; Tseren-Ochir, E.-O.; Kwon, J.-H.; Noh, J.-Y.; Lee, J.-B.; Park, S.-Y.; Choi, I.-S.; Song, C.-S. Pre-immune state induced by chicken interferon gamma inhibits the replication of H1N1 human and H9N2 avian influenza viruses in chicken embryo fibroblasts. Virol. J. 2016, 13, 71. [Google Scholar] [CrossRef]
- Xia, C.; Liu, J.; Wu, Z.; Lin, C.; Wang, M. The interferon-α genes from three chicken lines and its effects on H9N2 influenza viruses. Anim. Biotechnol. 2004, 15, 77–88. [Google Scholar] [CrossRef]
- Cornelissen, J.; Post, J.; Peeters, B.; Vervelde, L.; Rebel, J. Differential innate responses of chickens and ducks to low-pathogenic avian influenza. Avian Pathol. 2012, 41, 519–529. [Google Scholar] [CrossRef]
- Barbour, E.K.; Mastori, F.A.; Nour, A.A.; Shaib, H.A.; Jaber, L.S.; Yaghi, R.H.; Sabra, A.; Sleiman, F.T.; Sawaya, R.K.; Niedzwieck, A. Standardisation of a new model of H9N2/Escherichia coli challenge in broilers in the Lebanon. Vet. Ital. 2009, 45, 317–322. [Google Scholar] [PubMed]
- Li, H.; Liu, X.; Chen, F.; Zuo, K.; Wu, C.; Yan, Y.; Chen, W.; Lin, W.; Xie, Q. Avian influenza virus subtype H9N2 affects intestinal microbiota, barrier structure injury, and inflammatory intestinal disease in the chicken ileum. Viruses 2018, 10, 270. [Google Scholar] [CrossRef] [PubMed]
- Oakley, B.B.; Kogut, M.H. Spatial and temporal changes in the broiler chicken cecal and fecal microbiomes and correlations of bacterial taxa with cytokine gene expression. Front. Vet. Sci. 2016, 3, 11. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, F.; Wei, H.; Lian, Z.-X.; Sun, R.; Tian, Z. Respiratory influenza virus infection induces intestinal immune injury via microbiota-mediated Th17 cell–dependent inflammation. J. Exp. Med. 2014, 211, 2397–2410. [Google Scholar] [CrossRef] [PubMed]
- Keita, å.v.; Söderholm, J.D. The intestinal barrier and its regulation by neuroimmune factors. Neurogastroenterol. Motil. 2010, 22, 718–733. [Google Scholar] [CrossRef] [PubMed]
- Fagarasan, S. Intestinal IgA synthesis: A primitive form of adaptive immunity that regulates microbial communities in the gut. In Gut-Associated Lymphoid Tissues; Springer: Berlin, Germany, 2006; pp. 137–153. [Google Scholar]
- Burkholder, K.; Thompson, K.; Einstein, M.; Applegate, T.; Patterson, J. Influence of stressors on normal intestinal microbiota, intestinal morphology, and susceptibility to Salmonella enteritidis colonization in broilers. Poult. Sci. 2008, 87, 1734–1741. [Google Scholar] [CrossRef] [PubMed]
- Chappell, L.; Kaiser, P.; Barrow, P.; Jones, M.A.; Johnston, C.; Wigley, P. The immunobiology of avian systemic salmonellosis. Vet. Immunol. Immunopathol. 2009, 128, 53–59. [Google Scholar] [CrossRef]
- Hansson, G.C. Role of mucus layers in gut infection and inflammation. Curr. Opin. Microbiol. 2012, 15, 57–62. [Google Scholar] [CrossRef]
- Taupin, D.; Kinoshita, K.; Podolsky, D. Intestinal trefoil factor confers colonic epithelial resistance to apoptosis. Proc. Natl. Acad. Sci. USA 2000, 97, 799–804. [Google Scholar] [CrossRef] [Green Version]
- Sands, B.; Podolsky, D. The trefoil peptide family. Annu. Rev. Physiol. 1996, 58, 253–273. [Google Scholar] [CrossRef]
- Furuse, M.; Fujita, K.; Hiiragi, T.; Fujimoto, K.; Tsukita, S. Claudin-1 and-2: Novel integral membrane proteins localizing at tight junctions with no sequence similarity to occludin. J. Cell Biol. 1998, 141, 1539–1550. [Google Scholar] [CrossRef] [PubMed]
- Gumbiner, B.; Lowenkopf, T.; Apatira, D. Identification of a 160-kDa polypeptide that binds to the tight junction protein ZO-1. Proc. Natl. Acad. Sci. USA 1991, 88, 3460–3464. [Google Scholar] [CrossRef] [PubMed]
- Willott, E.; Balda, M.S.; Fanning, A.S.; Jameson, B.; Van Itallie, C.; Anderson, J.M. The tight junction protein ZO-1 is homologous to the Drosophila discs-large tumor suppressor protein of septate junctions. Proc. Natl. Acad. Sci. USA 1993, 90, 7834–7838. [Google Scholar] [CrossRef] [PubMed]
- Haskins, J.; Gu, L.; Wittchen, E.S.; Hibbard, J.; Stevenson, B.R. ZO-3, a novel member of the MAGUK protein family found at the tight junction, interacts with ZO-1 and occludin. J. Cell Biol. 1998, 141, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Befus, A.D.; Johnston, N.; Leslie, G.; Bienenstock, J. Gut-associated lymphoid tissue in the chicken. I. Morphology, ontogeny, and some functional characteristics of Peyer’s patches. J. Immunol. 1980, 125, 2626–2632. [Google Scholar]
- Liebler-Tenorio, E.M.; Pabst, R. MALT structure and function in farm animals. Vet. Res. 2006, 37, 257–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vindigni, S.M.; Zisman, T.L.; Suskind, D.L.; Damman, C.J. The intestinal microbiome, barrier function, and immune system in inflammatory bowel disease: A tripartite pathophysiological circuit with implications for new therapeutic directions. Ther. Adv. Gastroenterol. 2016, 9, 606–625. [Google Scholar] [CrossRef]
- Ruby, T.; Whittaker, C.; Withers, D.R.; Chelbi-Alix, M.K.; Morin, V.; Oudin, A.; Young, J.R.; Zoorob, R. Transcriptional profiling reveals a possible role for the timing of the inflammatory response in determining susceptibility to a viral infection. J. Virol. 2006, 80, 9207–9216. [Google Scholar] [CrossRef]
- Li, Z.; Wang, Y.; Xue, Y.; Li, X.; Cao, H.; Zheng, S.J. Critical role for voltage-dependent anion channel 2 in infectious bursal disease virus-induced apoptosis in host cells via interaction with VP5. J. Virol. 2012, 86, 1328–1338. [Google Scholar] [CrossRef]
- Li, L.; Kubasová, T.; Rychlik, I.; Hoerr, F.J.; Rautenschlein, S. Infectious bursal disease virus infection leads to changes in the gut associated-lymphoid tissue and the microbiota composition. PLoS ONE 2018, 13, e0192066. [Google Scholar] [CrossRef]
- Frank, D.N.; Amand, A.L.S.; Feldman, R.A.; Boedeker, E.C.; Harpaz, N.; Pace, N.R. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc. Natl. Acad. Sci. USA 2007, 104, 13780–13785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermúdez-Humarán, L.G.; Gratadoux, J.-J.; Blugeon, S.; Bridonneau, C.; Furet, J.-P.; Corthier, G. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef] [PubMed]
- Willing, B.; Halfvarson, J.; Dicksved, J.; Rosenquist, M.; Jänerot, G.; Engstrand, L.; Tysk, C.; Jansson, J.K. Twin studies reveal specific imbalances in the mucosa-associated microbiota of patients with ileal Crohns disease. Inflamm. Bowel Dis. 2009, 15, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Christophersen, C.; Morrison, M.; Conlon, M. Overestimation of the abundance of sulfate-reducing bacteria in human feces by quantitative PCR targeting the Desulfovibrio 16S rRNA gene. Appl. Env. Microbiol. 2011, 77, 3544–3546. [Google Scholar] [CrossRef]
- Blachier, F.; Davila, A.-M.; Mimoun, S.; Benetti, P.-H.; Atanasiu, C.; Andriamihaja, M.; Benamouzig, R.; Bouillaud, F.; Tomé, D. Luminal sulfide and large intestine mucosa: Friend or foe? Amino Acids 2010, 39, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.H.; Hold, G.L.; Harmsen, H.J.; Stewart, C.S.; Flint, H.J. Growth requirements and fermentation products of Fusobacterium prausnitzii, and a proposal to reclassify it as Faecalibacterium prausnitzii gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 2141–2146. [Google Scholar] [PubMed]
- Van Deun, K.; Pasmans, F.; van Immerseel, F.; Ducatelle, R.; Haesebrouck, F. Butyrate protects Caco-2 cells from Campylobacter jejuni invasion and translocation. Br. J. Nutr. 2008, 100, 480–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Pielsticker, C.; Han, Z.; Kubasová, T.; Rychlik, I.; Kaspers, B.; Rautenschlein, S. Infectious bursal disease virus inoculation infection modifies Campylobacter jejuni–host interaction in broilers. Gut Pathog. 2018, 10, 13. [Google Scholar] [CrossRef] [PubMed]
- Subler, K.A.; Mickael, C.S.; Jackwood, D.J. Infectious bursal disease virus-induced immunosuppression exacerbates Campylobacter jejuni colonization and shedding in chickens. Avian Dis. 2006, 50, 179–184. [Google Scholar] [CrossRef]
- MacDonald, T.T.; Monteleone, G. Immunity, inflammation, and allergy in the gut. Science 2005, 307, 1920–1925. [Google Scholar] [CrossRef]
- Lukert, P.D.; Saif, Y.M. Infectious Bursal Disease, 10th ed.; Calnek, B.W., Barnes, H.J., Beard, C.W., McDougald, L.R., Saif, Y.M., Eds.; Iowa State University Press: Ames, IA, USA, 1997; pp. 721–738. [Google Scholar]
- Bautista, D.; Elankumaran, S.; Heckert, R. Effect of a variant infectious bursal disease virus (E/Del) on Salmonella typhimurium infection in commercial broiler chickens. Avian Dis. 2004, 48, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Wyeth, P. Effect of infectious bursal disease on the response of chickens to Salmonella typhimurium and Escherichia coli infections. Vet. Rec. 1975, 96, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Arafat, N.; Eladl, A.H.; Mahgoub, H.; El-Shafei, R.A. Effect of infectious bursal disease (IBD) vaccine on Salmonella Enteritidis infected chickens. Vaccine 2017, 35, 3682–3689. [Google Scholar] [CrossRef] [PubMed]
- Naseem, S.; Rahman, S.; Shafee, M.; Sheikh, A.; Khan, A. Immunomodulatory and growth-promoting effect of a probiotic supplemented in the feed of broiler chicks vaccinated against infectious bursal disease. Braz. J. Poult. Sci. 2012, 14, 109–113. [Google Scholar] [CrossRef]
- Witter, R.; Gimeno, I.; Reed, W.; Bacon, L. An acute form of transient paralysis induced by highly virulent strains of Marek’s disease virus. Avian Dis. 1999, 43, 704–720. [Google Scholar] [CrossRef] [PubMed]
- Calnek, B.; Harris, R.; Buscaglia, C.; Schat, K.; Lucio, B. Relationship between the immunosuppressive potential and the pathotype of Marek’s disease virus isolates. Avian Dis. 1998, 42, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Perumbakkam, S.; Hunt, H.D.; Cheng, H.H. Marek’s disease virus influences the core gut microbiome of the chicken during the early and late phases of viral replication. Fems Microbiol. Ecol. 2014, 90, 300–312. [Google Scholar] [CrossRef] [PubMed]
- Giansanti, F.; Giardi, M.F.; Massucci, M.T.; Botti, D.; Antonini, G. Ovotransferrin expression and release by chicken cell lines infected with Marek’s disease virus. Biochem. Cell Biol. 2007, 85, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Giansanti, F.; Leboffe, L.; Angelucci, F.; Antonini, G. The nutraceutical properties of ovotransferrin and its potential utilization as a functional food. Nutrients 2015, 7, 9105–9115. [Google Scholar] [CrossRef]
- Giardi, M.F.; La Torre, C.; Giansanti, F.; Botti, D. Effects of transferrins and cytokines on nitric oxide production by an avian lymphoblastoid cell line infected with Marek’s disease virus. Antivir. Res. 2009, 81, 248–252. [Google Scholar] [CrossRef]
- Atarashi, K.; Tanoue, T.; Oshima, K.; Suda, W.; Nagano, Y.; Nishikawa, H.; Fukuda, S.; Saito, T.; Narushima, S.; Hase, K. T reg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 2013, 500, 232. [Google Scholar] [CrossRef] [PubMed]
- Perumbakkam, S.; Hunt, H.D.; Cheng, H.H. Differences in CD8αα and cecal microbiome community during proliferation and late cytolytic phases of Marek’s disease virus infection are associated with genetic resistance to Marek’s disease. Fems Microbiol. Ecol. 2016, 92, fiw188. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Finegold, S.M.; Song, Y.; Lawson, P.A. Reclassification of Clostridium coccoides, Ruminococcus hansenii, Ruminococcus hydrogenotrophicus, Ruminococcus luti, Ruminococcus productus and Ruminococcus schinkii as Blautia coccoides gen. nov., comb. nov., Blautia hansenii comb. nov., Blautia hydrogenotrophica comb. nov., Blautia luti comb. nov., Blautia producta comb. nov., Blautia schinkii comb. nov. and description of Blautia wexlerae sp. nov., isolated from human faeces. Int. J. Syst. Evol. Microbiol. 2008, 58, 1896–1902. [Google Scholar] [PubMed]
- Biddle, A.; Stewart, L.; Blanchard, J.; Leschine, S. Untangling the genetic basis of fibrolytic specialization by Lachnospiraceae and Ruminococcaceae in diverse gut communities. Diversity 2013, 5, 627–640. [Google Scholar] [CrossRef]
- Wages, D.P. Streptococcosis. In Diseases of Poultry, 11th ed.; Saif, Y.M., Barnes, H.J., Glisson, J.R., Fadly, A.M., McDougald, L.R., Swayne, D.E., Eds.; Iowa State Press: Ames, IA, USA, 2003; p. 805812. [Google Scholar]
- Wakamatsu, N.; King, D.; Kapczynski, D.; Seal, B.; Brown, C. Experimental pathogenesis for chickens, turkeys, and pigeons of exotic Newcastle disease virus from an outbreak in California during 2002–2003. Vet. Pathol. 2006, 43, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Brown, V.R.; Bevins, S.N. A review of virulent Newcastle disease viruses in the United States and the role of wild birds in viral persistence and spread. Vet. Res. 2017, 48, 68. [Google Scholar] [CrossRef] [PubMed]
- Baron, S.; Buckler, C.E. Circulating interferon in mice after intravenous injection of virus. Science 1963, 141, 1061–1063. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.-Y.; Landay, M.E. Enhancement of the lethal effects of endotoxins by interferon inducers. J. Bacteriol. 1969, 100, 1110. [Google Scholar]
- Zhang, L.; Wang, G.; Yu, H.; Wang, J.; Wang, S.; Jia, Y.; Yu, Y.; Xu, J. First report of human infection by Rhodoplanes sp., Alphaproteobacteria in China. Asian Pac. J. Trop. Med. 2011, 4, 248–250. [Google Scholar] [CrossRef] [Green Version]
- Cui, N.; Huang, X.; Kong, Z.; Huang, Y.; Huang, Q.; Yang, S.; Zhang, L.; Xu, C.; Zhang, X.; Cui, Y. Newcastle disease virus infection interferes with the formation of intestinal microflora in newly hatched specific-pathogen-free chicks. Front. Microbiol. 2018, 9, 900. [Google Scholar] [CrossRef]
- Girardin, H.; Albagnac, C.; Dargaignaratz, C.; Nguyen-The, C.; Carlin, F. Antimicrobial activity of foodborne Paenibacillus and Bacillus spp. against Clostridium botulinum. J. Food Prot. 2002, 65, 806–813. [Google Scholar] [CrossRef] [PubMed]
- Passera, A.; Venturini, G.; Battelli, G.; Casati, P.; Penaca, F.; Quaglino, F.; Bianco, P.A. Competition assays revealed Paenibacillus pasadenensis strain R16 as a novel antifungal agent. Microbiol. Res. 2017, 198, 16–26. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Virus | Control Group | Infected Group | ||
|---|---|---|---|---|
| Commensals | Effector Molecules and Outcomes | Pathogens | Effector Molecules and Outcomes | |
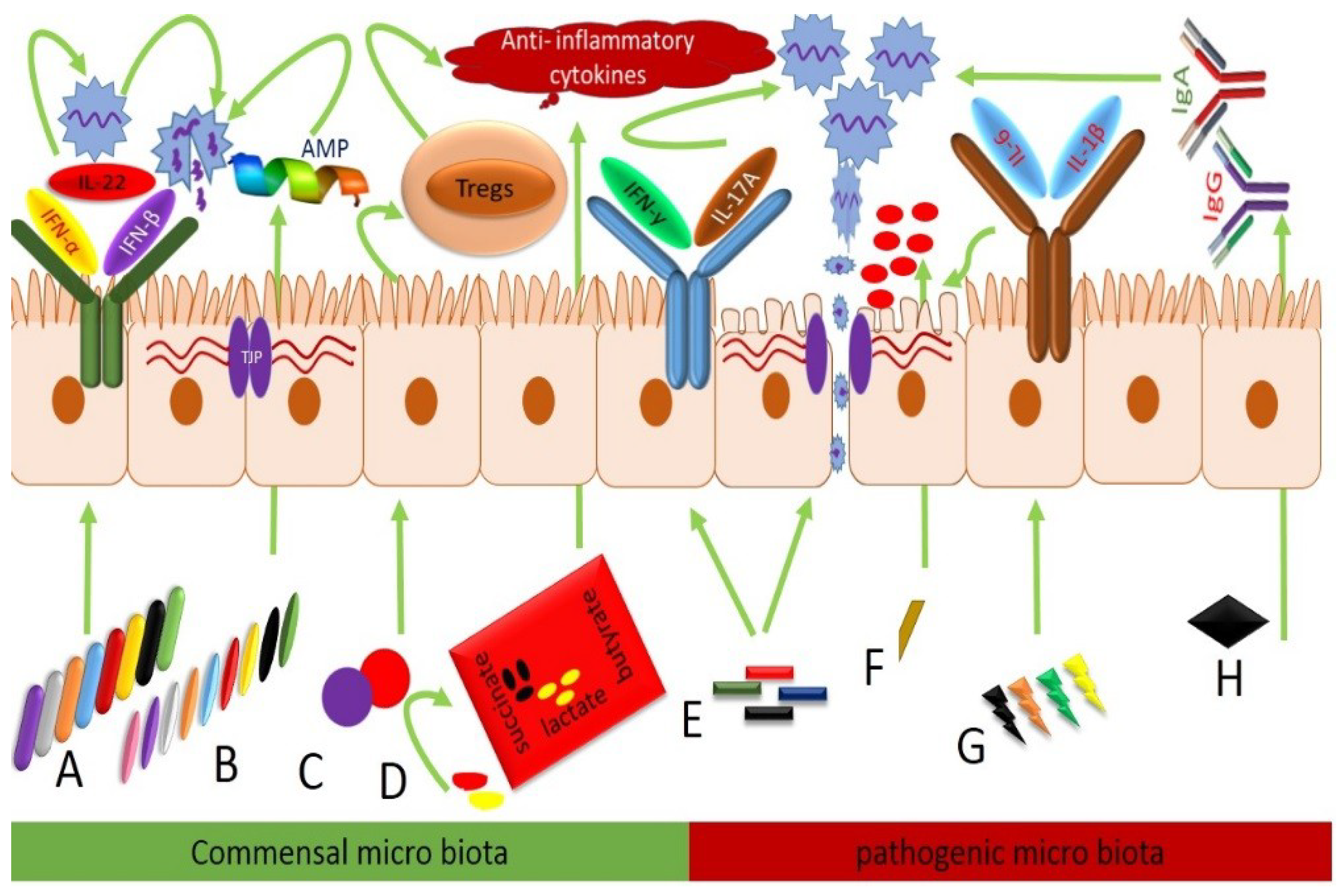
| AIV | Collinsella, Faecalibacterium, Oscillibacter, Holdemanella, Pseudoflavonifractor, Anaerotruncus, Butyricoccus, and Bifidobacterium | Increase IFN-α, IFN-β, and IL-22 and antimicrobial peptides such as MUC, TFF, ZO, and tight junction proteins comprised of claudins, occludin, and zona occludens mRNA expressions | Proteobacteria Clostridium cluster XI, Escherichia, Shigella, Salmonella, Vampirovibrio, Clostridium cluster XIVb, and genus Ruminococcus | Downregulate the IFN-α, IFN-β, and IL-22 secretion and antimicrobial peptides such as MUC, TFF, ZO, and tight junction proteins comprised of claudins, occludin, and zona occludens mRNA expressions also enhance the secretions of proinflammatory cytokines IFN-γ, IL-17A, IL-6, and IL-1B and produce inflammation |
| IBDV | Clostridium XlVa | Induce T regulatory cells to produce anti-inflammatory cytokines | Desulfovibrionaceae | Produce hydrogen sulfides and cause inflammation |
| Faecalibacterium | Enhance butyrate shortchain fatty acids (SCFA) and suppress the inflammation | Campylobacter jejuni | Inhibit butyrate SCFA production cause inflammation of GIT | |
| Probiotics | Increase immunoglobulins, FCR body weight gain | Salmonella typhimurium and Campylobacter jejuni | Decreased IgG and IgA production | |
| MDV | Firmicutes | Induce T regulatory cells to produce anti-inflammatory cytokines | Pathogenic Lactobacillus spp., Proteobacteria | Suppress the T regulatory cells stimulation produce inflammation |
| Blautia spp. and Faecalibacterium spp. | Produce succinate and lactate and provide energy and reduce inflammation | Streptococcus spp. | Septicemia, peritonitis, and endocarditis | |
| NDV | Paenibacillus and Enterococcus | Antimicrobial peptides | Rhodoplanes, Clostridium, and Epulopiscium | Cause local mucosal infection |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abaidullah, M.; Peng, S.; Kamran, M.; Song, X.; Yin, Z. Current Findings on Gut Microbiota Mediated Immune Modulation against Viral Diseases in Chicken. Viruses 2019, 11, 681. https://doi.org/10.3390/v11080681
Abaidullah M, Peng S, Kamran M, Song X, Yin Z. Current Findings on Gut Microbiota Mediated Immune Modulation against Viral Diseases in Chicken. Viruses. 2019; 11(8):681. https://doi.org/10.3390/v11080681
Chicago/Turabian StyleAbaidullah, Muhammad, Shuwei Peng, Muhammad Kamran, Xu Song, and Zhongqiong Yin. 2019. "Current Findings on Gut Microbiota Mediated Immune Modulation against Viral Diseases in Chicken" Viruses 11, no. 8: 681. https://doi.org/10.3390/v11080681
APA StyleAbaidullah, M., Peng, S., Kamran, M., Song, X., & Yin, Z. (2019). Current Findings on Gut Microbiota Mediated Immune Modulation against Viral Diseases in Chicken. Viruses, 11(8), 681. https://doi.org/10.3390/v11080681