Feline Foamy Virus Infection: Characterization of Experimental Infection and Prevalence of Natural Infection in Domestic Cats with and without Chronic Kidney Disease




 , ,
, ,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Virus Generation
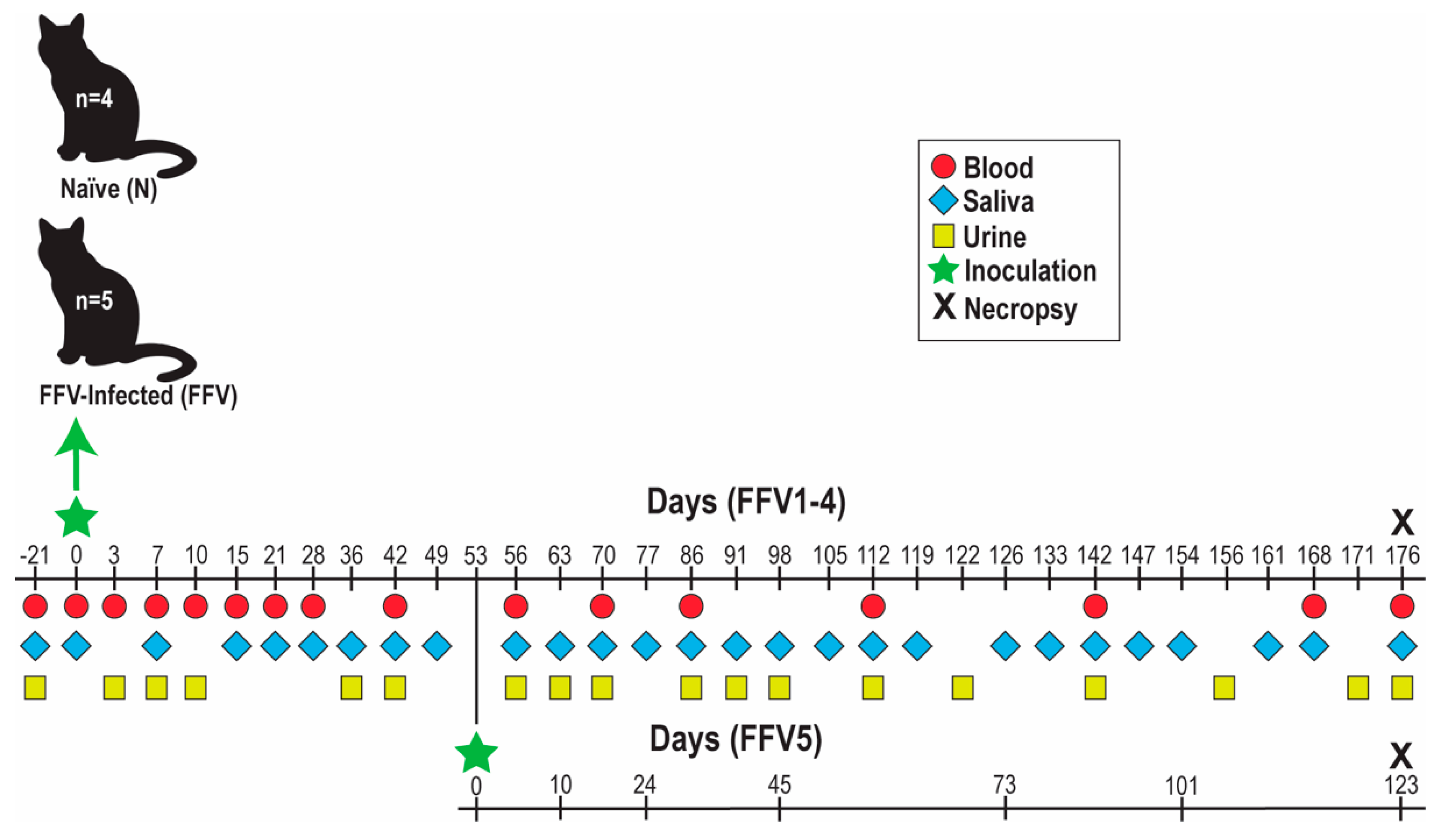
2.2. Animals and Study Design for Experimental FFV Inoculation
2.3. Nested and Real-Time Quantitative PCR for Virus Detection and Quantification
2.4. Hematological and Flow Cytometric Analyses for PBMC Phenotyping
2.5. Gross Necropsy and Histologic Characterization of Tissues
2.6. Renal Tissue Microscopic and Ultrastructural Examination
2.7. Samples from Australian Pet Cats with CKD and Non-Azometic, Age-Matched Controls
2.8. Statistical Analyses
3. Results
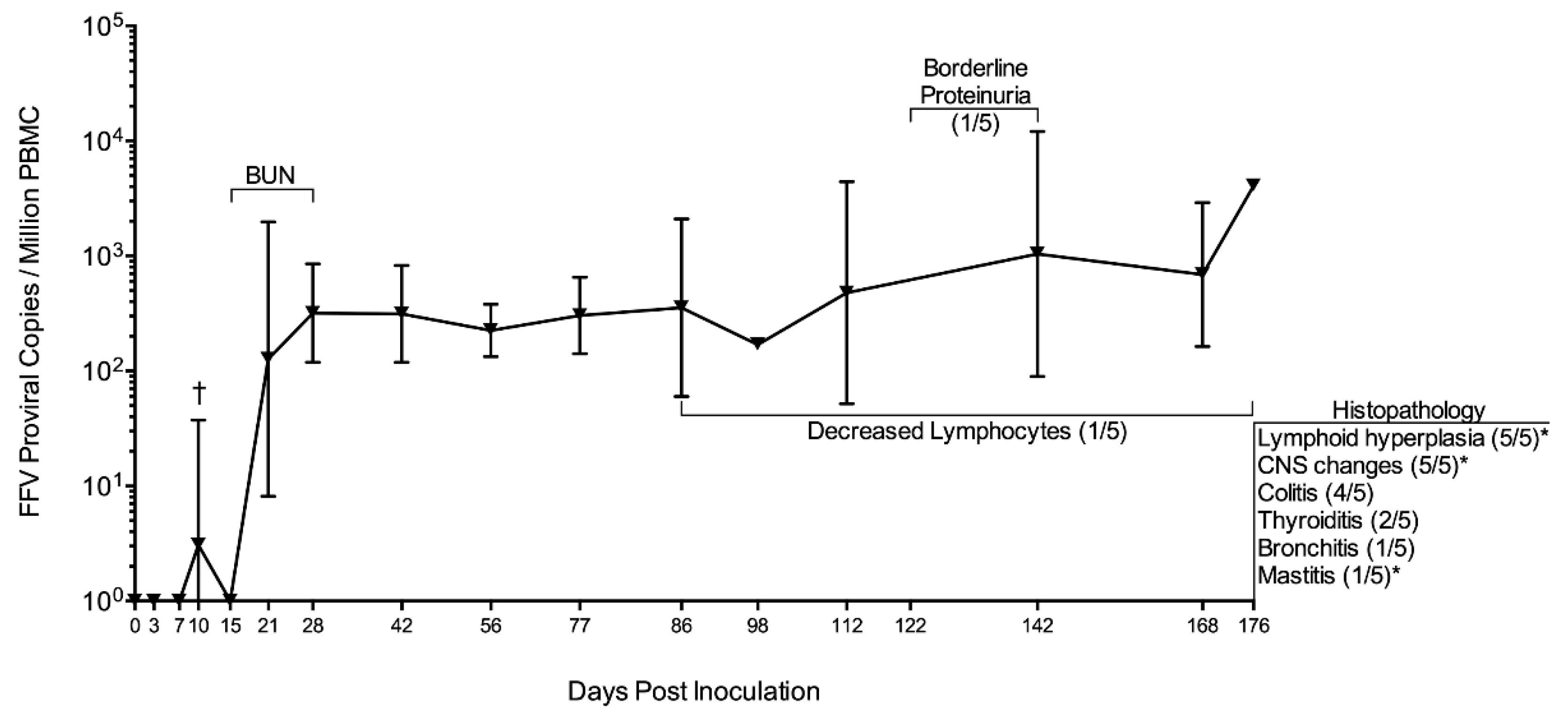
3.1. FFV-Infected Cats Did Not Show Clinical Signs of Infection Despite a Persistent FFV Proviral Load and Specific Humoral Response
3.2. FFV Provirus Tissue Tropism Is Primarily Lymphoid
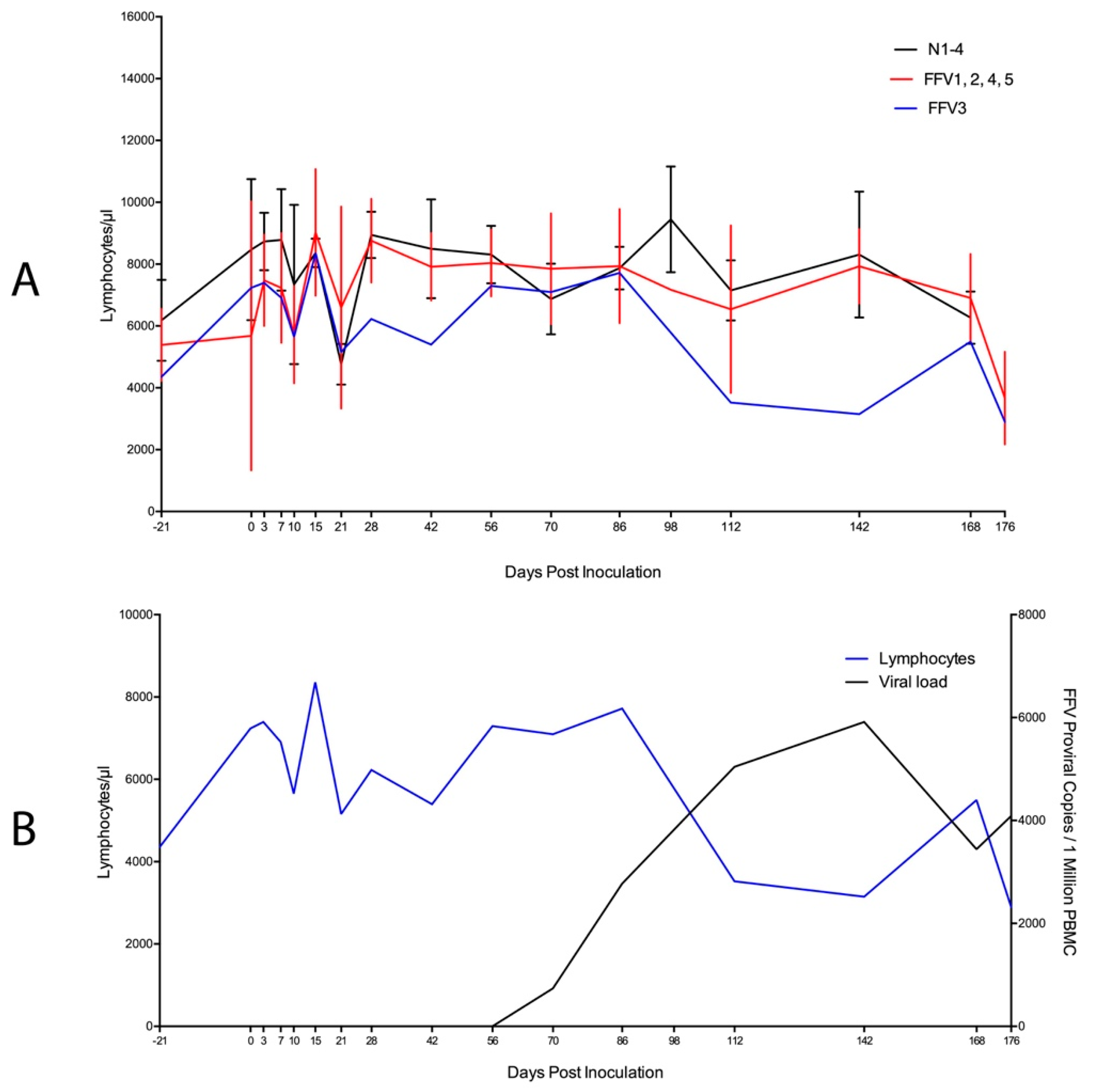
3.3. Significant PBMC Phenotypic Changes Were Rare though a Negative Correlation Was Found between Lymphocytes and FFV Proviral Load in Cat FFV3
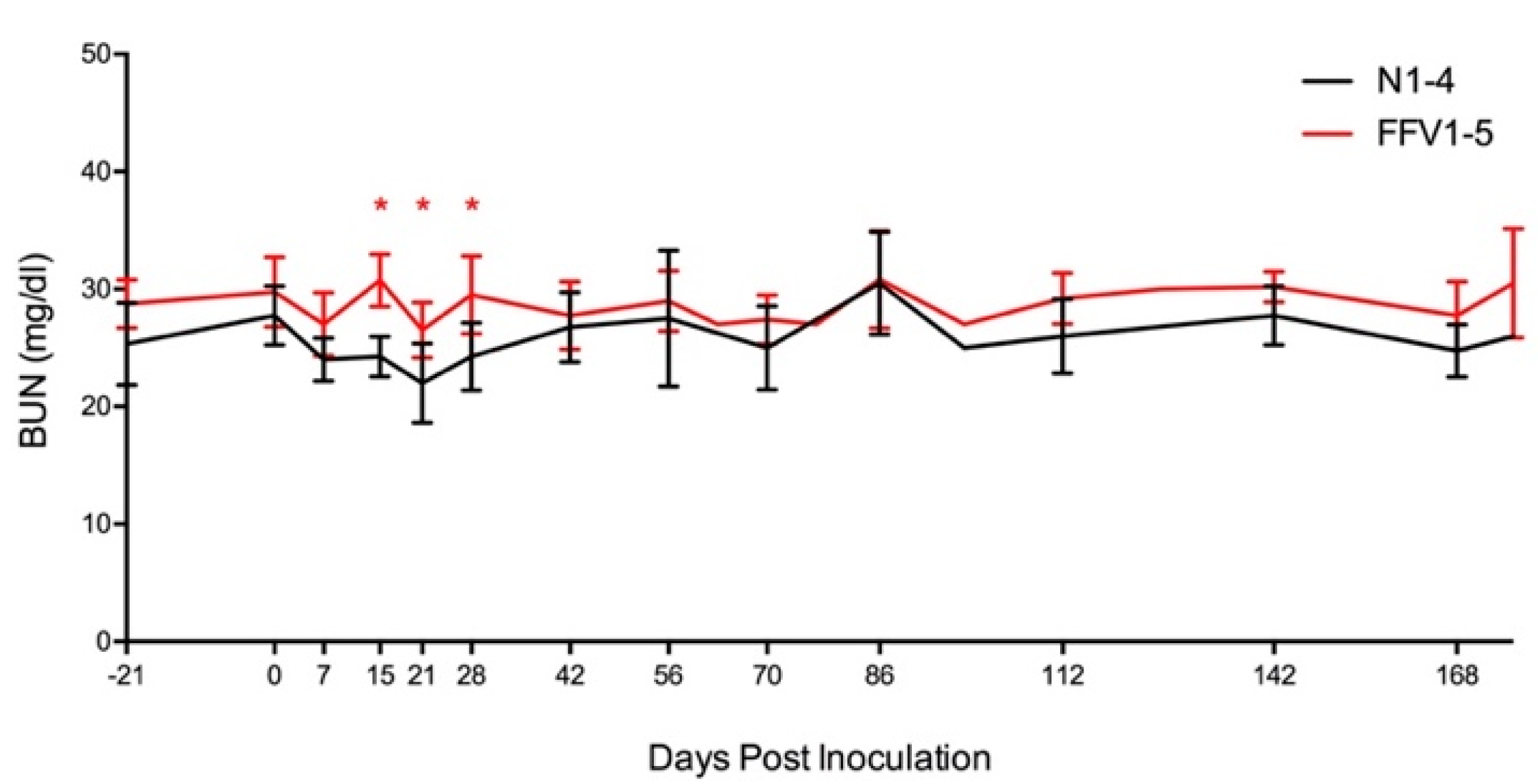
3.4. Subtle Differences in Urine and Hematological Parameters Were Detected between Experimentally Infected and Control Cats
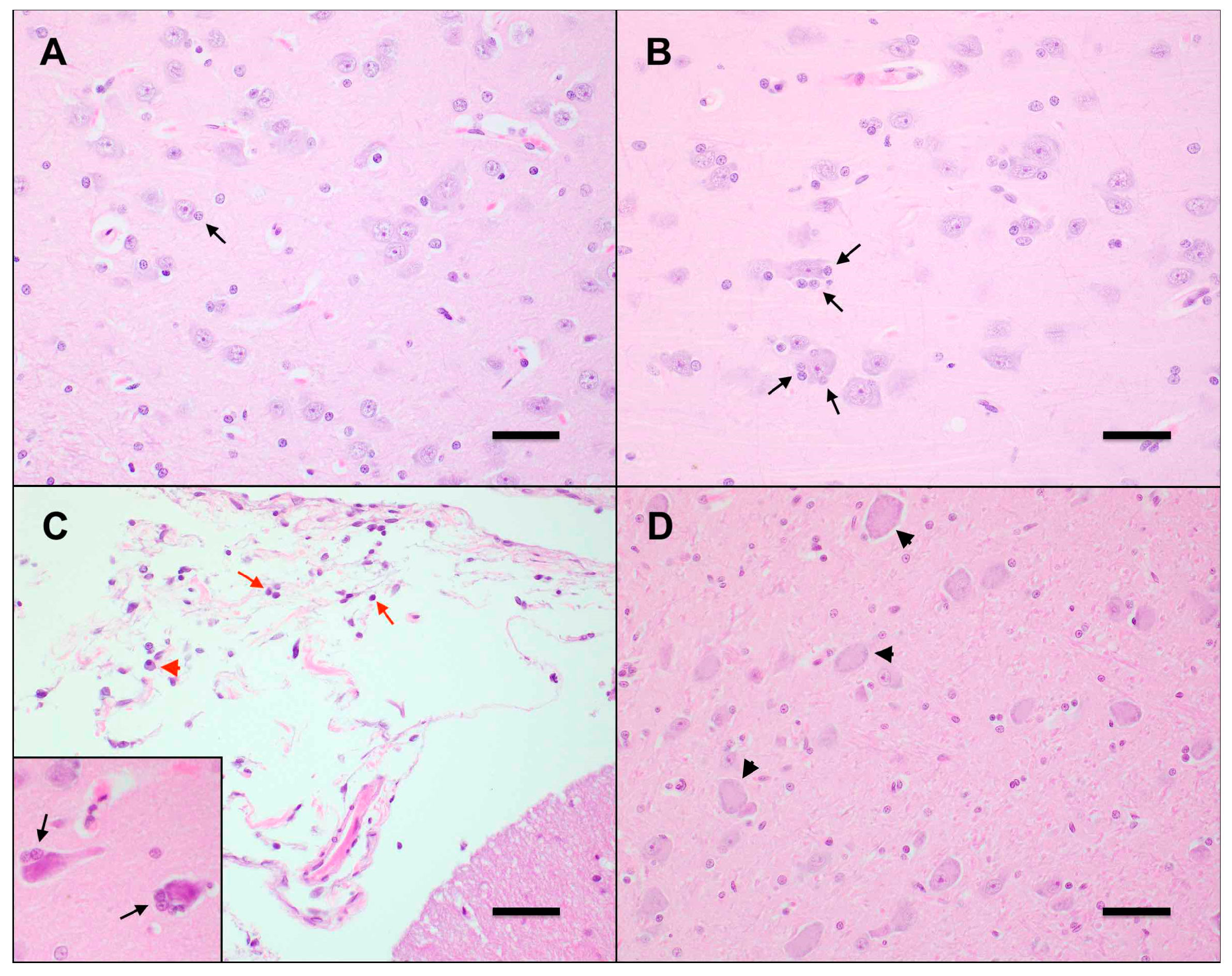
3.5. Mild Lymphoplasmacytic Infiltrates and Lymphoid Hyperplasia of Multiple Tissues Were Associated with FFV Exposure
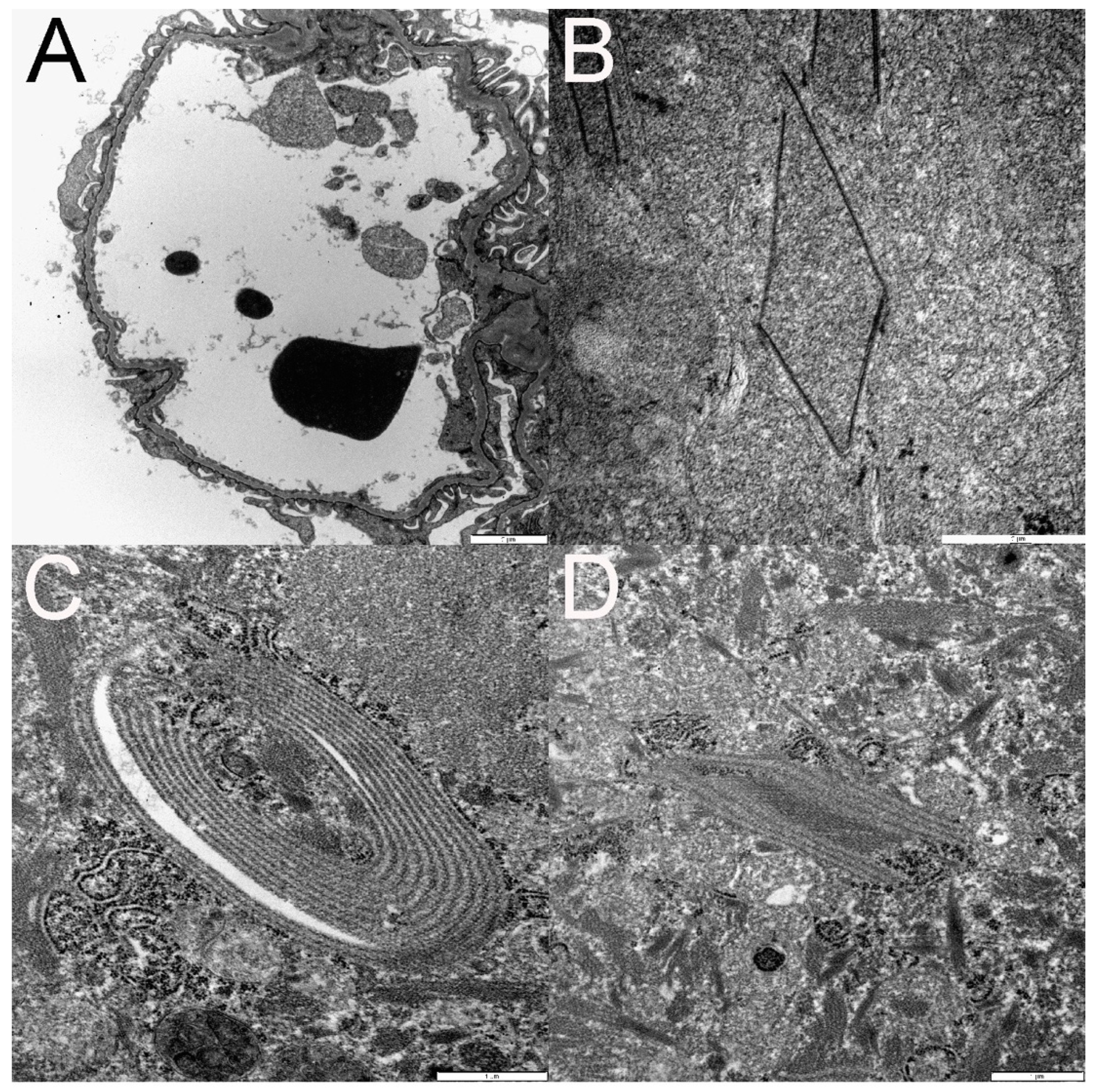
3.6. Ultrastructural Changes Were Noted in the Kidneys of FFV-Infected Cats
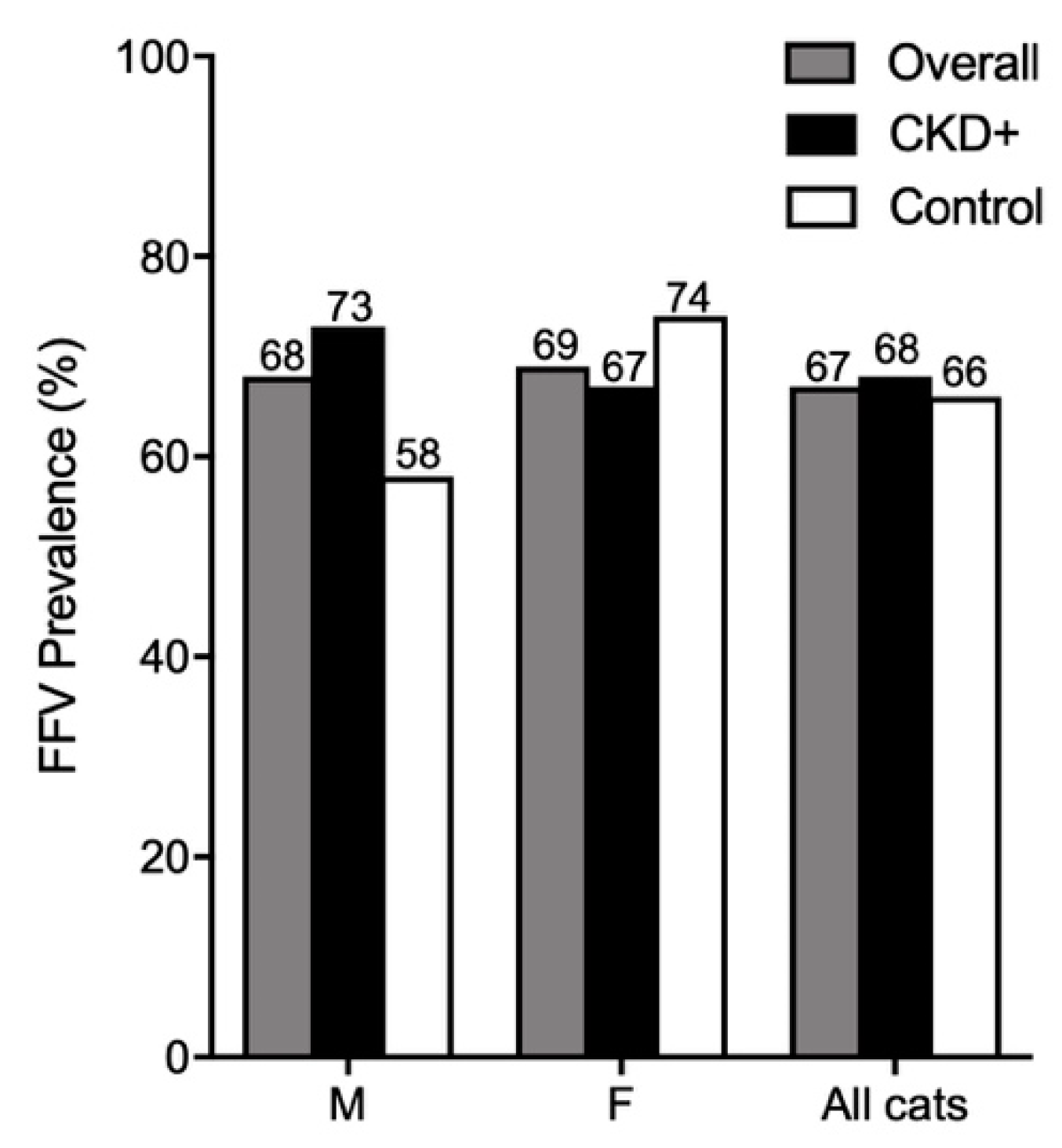
3.7. FFV Is Highly Prevalent in Australian Domestic Pet Cats
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Riggs, J.L.; Oshiro, L.S.; Taylor, D.O.; Lennette, E.H. Syncytium-forming agent isolated from domestic cats. Nature 1969, 222, 1190–1191. [Google Scholar] [CrossRef] [PubMed]
- Aiewsakun, P.; Katzourakis, A. Marine origin of retroviruses in the early palaeozoic era. Nat. Commun. 2017, 8, 13954. [Google Scholar] [CrossRef] [PubMed]
- Kasza, L.; Hayward, A.H.; Betts, A.O. Isolation of a virus from a cat sarcoma in an established canine melanoma cell line. Res. Vet. Sci. 1969, 10, 216–218. [Google Scholar] [CrossRef]
- Shroyer, E.L.; Shalaby, M.R. Isolation of feline syncytia-forming virus from oropharyngeal swab samples and buffy coat cells. Am. J. Vet. Res. 1978, 39, 555–560. [Google Scholar] [PubMed]
- Ellis, T.M.; MacKenzie, J.S.; Wilcox, G.E.; Cook, R.D. Isolation of feline syncytia-forming virus from oropharyngeal swabs of cats. Aust. Vet. J. 1979, 55, 202–203. [Google Scholar] [CrossRef]
- German, A.C.; Harbour, D.A.; Helps, C.R.; Gruffydd-Jones, T.J. Is feline foamy virus really apathogenic? Vet. Immunol. Immunopathol. 2008, 123, 114–118. [Google Scholar] [CrossRef]
- Pedersen, N.C. Feline syncytium-forming virus infection. In Diseases of the Cat; Holyworth, J., Ed.; The W. B. Saunder Co.: Philadelphia, PA, USA, 1986; pp. 268–272. [Google Scholar]
- Alke, A.; Schwantes, A.; Zemba, M.; Flugel, R.M.; Löchelt, M. Characterization of the humoral immune response and virus replication in cats experimentally infected with feline foamy virus. Virology 2000, 275, 170–176. [Google Scholar] [CrossRef]
- Von Laer, D.; Neumann-Haefelin, D.; Heeney, J.L.; Schweizer, M. Lymphocytes are the major reservoir for foamy viruses in peripheral blood. Virology 1996, 221, 240–244. [Google Scholar] [CrossRef]
- Ledesma-Feliciano, C.; Hagen, S.; Troyer, R.; Zheng, X.; Musselman, E.; Slavkovic Lukic, D.; Franke, A.M.; Maeda, D.; Zielonka, J.; Munk, C.; et al. Replacement of feline foamy virus bet by feline immunodeficiency virus vif yields replicative virus with novel vaccine candidate potential. Retrovirology 2018, 15, 38. [Google Scholar] [CrossRef]
- Weikel, J.; Löchelt, M.; Truyen, U. Demonstration of feline foamy virus in experimentally infected cats by immunohistochemistry. J. Vet. Med. Physiol. Pathol. Clin. Med. 2003, 50, 415–417. [Google Scholar] [CrossRef]
- Kehl, T.; Tan, J.; Materniak, M. Non-simian foamy viruses: Molecular virology, tropism and prevalence and zoonotic/interspecies transmission. Viruses 2013, 5, 2169–2209. [Google Scholar] [CrossRef]
- Liu, W.; Lei, J.; Liu, Y.; Lukic, D.S.; Rathe, A.M.; Bao, Q.; Kehl, T.; Bleiholder, A.; Hechler, T.; Löchelt, M. Feline foamy virus-based vectors: Advantages of an authentic animal model. Viruses 2013, 5, 1702–1718. [Google Scholar] [CrossRef]
- Romen, F.; Backes, P.; Materniak, M.; Sting, R.; Vahlenkamp, T.W.; Riebe, R.; Pawlita, M.; Kuzmak, J.; Löchelt, M. Serological detection systems for identification of cows shedding bovine foamy virus via milk. Virology 2007, 364, 123–131. [Google Scholar] [CrossRef]
- Yamamoto, J.K.; Sparger, E.; Ho, E.W.; Andersen, P.R.; O’connor, T.P.; Mandell, C.P.; Lowenstine, L.; Munn, R.; Pedersen, N.C. Pathogenesis of experimentally induced feline immunodeficiency virus infection in cats. Am. J. Vet. Res. 1988, 49, 1246–1258. [Google Scholar]
- Winkler, I.G.; Löchelt, M.; Flower, R.L.P. Epidemiology of feline foamy virus and feline immunodeficiency virus infections in domestic and feral cats: A seroepidemiological study. J. Clin. Microbiol. 1999, 37, 2848–2851. [Google Scholar]
- Bandecchi, P.; Matteucci, D.; Baldinotti, F.; Guidi, G.; Abramo, F.; Tozzini, F.; Bendinelli, M. Prevalence of feline immunodeficiency virus and other retroviral infections in sick cats in italy. Vet. Immunol. Immunopathol. 1992, 31, 337–345. [Google Scholar] [CrossRef]
- Glaus, T.; Hofmann-Lehmann, R.; Greene, C.; Glaus, B.; Wolfensberger, C.; Lutz, H. Seroprevalence of bartonella henselae infection and correlation with disease status in cats in switzerland. J. Clin. Microbiol. 1997, 35, 2883–2885. [Google Scholar]
- Winkler, I.G.; Lochelt, M.; Levesque, J.P.; Bodem, J.; Flugel, R.M.; Flower, R.L. A rapid streptavidin-capture elisa specific for the detection of antibodies to feline foamy virus. J. Immunol. Methods 1997, 207, 69–77. [Google Scholar] [CrossRef]
- Miyazawa, T.; Ikeda, Y.; Maeda, K.; Horimoto, T.; Tohya, Y.; Mochizuki, M.; Vu, D.; Vu, G.D.; Cu, D.X.; Ono, K.; et al. Seroepidemiological survey of feline retrovirus infections in domestic and leopard cats in northern vietnam in 1997. J. Vet. Med. Sci. 1998, 60, 1273–1275. [Google Scholar] [CrossRef]
- Winkler, I.G.; Flügel, R.M.; Löchelt, M.; Flower, R.L.P. Detection and molecular characterisation of feline foamy virus serotypes in naturally infected cats. Virology 1998, 247, 144–151. [Google Scholar] [CrossRef]
- Nakamura, K.; Miyazawa, T.; Ikeda, Y.; Sato, E.; Nishimura, Y.; Nguyen, N.T.; Takahashi, E.; Mochizuki, M.; Mikami, T. Contrastive prevalence of feline retrovirus infections between northern and southern vietnam. J. Vet. Med. Sci. 2000, 62, 921–923. [Google Scholar] [CrossRef]
- Bleiholder, A.; Mühle, M.; Hechler, T.; Bevins, S.; vandeWoude, S.; Denner, J.; Löchelt, M. Pattern of seroreactivity against feline foamy virus proteins in domestic cats from germany. Vet. Immunol. Immunopathol. 2011, 143, 292–300. [Google Scholar] [CrossRef]
- Mochizuki, M.; Konishi, S. Feline syncytial virus spontaneously detected in feline cell cultures. Nihon Juigaku Zasshi 1979, 41, 351–362. [Google Scholar] [CrossRef]
- Flower, R.L.; Wilcox, G.E.; Cook, R.D.; Ellis, T.M. Detection and prevalence of serotypes of feline syncytial spumaviruses. Arch. Virol. 1985, 83, 53–63. [Google Scholar] [CrossRef]
- Romen, F.; Pawlita, M.; Sehr, P.; Bachmann, S.; Schroder, J.; Lutz, H.; Lochelt, M. Antibodies against Gag are diagnostic markers for feline foamy virus infections while Env and Bet reactivity is undetectable in a substantial fraction of infected cats. Virology 2006, 345, 502–508. [Google Scholar] [CrossRef]
- Kechejian, S.; Dannemiller, N.; Kraberger, S.; Ledesma-Feliciano, C.; Löchelt, M.; Carver, S.; VandeWoude, S. Feline foamy virus sero-prevalence and demographic risk factors in Colorado, Southern California and Florida stray domestic cat populations. JFMS Open Rep. 2019. under review. [Google Scholar]
- Switzer, W.M.; Bhullar, V.; Shanmugam, V.; Cong, M.E.; Parekh, B.; Lerche, N.W.; Yee, J.L.; Ely, J.J.; Boneva, R.; Chapman, L.E.; et al. Frequent simian foamy virus infection in persons occupationally exposed to nonhuman primates. J. Virol. 2004, 78, 2780–2789. [Google Scholar] [CrossRef]
- Switzer, W.M.; Garcia, A.D.; Yang, C.; Wright, A.; Kalish, M.L.; Folks, T.M.; Heneine, W. Coinfection with hiv-1 and simian foamy virus in west central africans. J. Infect. Dis. 2008, 197, 1389–1393. [Google Scholar] [CrossRef]
- Buseyne, F.; Betsem, E.; Montange, T.; Njouom, R.; Bilounga Ndongo, C.; Hermine, O.; Gessain, A. Clinical signs and blood test results among humans infected with zoonotic simian foamy virus: A case-control study. J. Infect. Dis. 2018, 218, 144–151. [Google Scholar] [CrossRef]
- Rua, R.; Betsem, E.; Montange, T.; Buseyne, F.; Gessain, A. In vivo cellular tropism of gorilla simian foamy virus in blood of infected humans. J. Virol. 2014, 88, 13429–13435. [Google Scholar] [CrossRef]
- Betsem, E.; Rua, R.; Tortevoye, P.; Froment, A.; Gessain, A. Frequent and recent human acquisition of simian foamy viruses through apes’ bites in central africa. PLoS Pathog 2011, 7, e1002306. [Google Scholar] [CrossRef]
- Cummins, J.E., Jr.; Boneva, R.S.; Switzer, W.M.; Christensen, L.L.; Sandstrom, P.; Heneine, W.; Chapman, L.E.; Dezzutti, C.S. Mucosal and systemic antibody responses in humans infected with simian foamy virus. J. Virol. 2005, 79, 13186–13189. [Google Scholar] [CrossRef]
- Boneva, R.S.; Switzer, W.M.; Spira, T.J.; Bhullar, V.B.; Shanmugam, V.; Cong, M.E.; Lam, L.; Heneine, W.; Folks, T.M.; Chapman, L.E. Clinical and virological characterization of persistent human infection with simian foamy viruses. AIDS Res. Hum. Retrovir. 2007, 23, 1330–1337. [Google Scholar] [CrossRef]
- Butera, S.T.; Brown, J.; Callahan, M.E.; Owen, S.M.; Matthews, A.L.; Weigner, D.D.; Chapman, L.E.; Sandstrom, P.A. Survey of veterinary conference attendees for evidence of zoonotic infection by feline retroviruses. J. Am. Vet. Med. Assoc. 2000, 217, 1475–1479. [Google Scholar] [CrossRef]
- Kiem, H.P.; Wu, R.A.; Sun, G.; von Laer, D.; Rossi, J.J.; Trobridge, G.D. Foamy combinatorial anti-hiv vectors with mgmtp140k potently inhibit hiv-1 and shiv replication and mediate selection in vivo. Gene Ther. 2010, 17, 37–49. [Google Scholar] [CrossRef]
- Lei, J.; Osen, W.; Gardyan, A.; Hotz-Wagenblatt, A.; Wei, G.; Gissmann, L.; Eichmuller, S.; Löchelt, M. Replication-competent foamy virus vaccine vectors as novel epitope scaffolds for immunotherapy. PLoS ONE 2015, 10, e0138458. [Google Scholar] [CrossRef]
- Schwantes, A.; Ortlepp, I.; Löchelt, M. Construction and functional characterization of feline foamy virus-based retroviral vectors. Virology 2002, 301, 53–63. [Google Scholar] [CrossRef]
- Trobridge, G.D.; Miller, D.G.; Jacobs, M.A.; Allen, J.M.; Kiem, H.P.; Kaul, R.; Russell, D.W. Foamy virus vector integration sites in normal human cells. Proc. Natl. Acad. Sci. USA 2006, 103, 1498–1503. [Google Scholar] [CrossRef]
- Burtner, C.R.; Beard, B.C.; Kennedy, D.R.; Wohlfahrt, M.E.; Adair, J.E.; Trobridge, G.D.; Scharenberg, A.M.; Torgerson, T.R.; Rawlings, D.J.; Felsburg, P.J.; et al. Intravenous injection of a foamy virus vector to correct canine scid-x1. Blood 2014, 123, 3578–3584. [Google Scholar] [CrossRef]
- Bauer, T.R., Jr.; Allen, J.M.; Hai, M.; Tuschong, L.M.; Khan, I.F.; Olson, E.M.; Adler, R.L.; Burkholder, T.H.; Gu, Y.C.; Russell, D.W.; et al. Successful treatment of canine leukocyte adhesion deficiency by foamy virus vectors. Nat. Med. 2008, 14, 93–97. [Google Scholar] [CrossRef]
- Schwantes, A.; Truyen, U.; Weikel, J.; Weiss, C.; Lochelt, M. Application of chimeric feline foamy virus-based retroviral vectors for the induction of antiviral immunity in cats. J. Virol. 2003, 77, 7830–7842. [Google Scholar] [CrossRef]
- Lindemann, D.; Steffen, I.; Pohlmann, S. Cellular entry of retroviruses. Adv. Exp. Med. Biol. 2013, 790, 128–149. [Google Scholar]
- Plochmann, K.; Horn, A.; Gschmack, E.; Armbruster, N.; Krieg, J.; Wiktorowicz, T.; Weber, C.; Stirnnagel, K.; Lindemann, D.; Rethwilm, A.; et al. Heparan sulfate is an attachment factor for foamy virus entry. J. Virol. 2012, 86, 10028–10035. [Google Scholar] [CrossRef]
- Pedersen, N.C.; Pool, R.R.; O’Brien, T. Feline chronic progressive polyarthritis. Am. J. Vet. Res. 1980, 41, 522–535. [Google Scholar]
- Yamamoto, J.K.; Hansen, H.; Ho, E.W.; Morishita, T.Y.; Okuda, T.; Sawa, T.R.; Nakamura, R.M.; Pedersen, N.C. Epidemiologic and clinical aspects of feline immunodeficiency virus infection in cats from the continental united states and canada and possible mode of transmission. J. Am. Vet. Med. Assoc. 1989, 194, 213–220. [Google Scholar]
- Inkpen, H. Chronic progressive polyarthritis in a domestic shorthair cat. Can. Vet. J. 2015, 56, 621–623. [Google Scholar]
- Whitman, J.E., Jr.; Cockrell, K.O.; Hall, W.T.; Gilmore, C.E. An unusual case of feline leukemia and an associated syncytium-forming virus. Am. J. Vet. Res. 1975, 36, 873–880. [Google Scholar]
- Powers, J.A.; Chiu, E.S.; Kraberger, S.J.; Roelke-Parker, M.; Lowery, I.; Erbeck, K.; Troyer, R.; Carver, S.; VandeWoude, S. Feline Leukemia Virus (FeLV) Disease Outcomes in a Domestic Cat Breeding Colony: Relationship to Endogenous FeLV and Other Chronic Viral. J. Virol. 2018, 92, e00649-18. [Google Scholar] [CrossRef]
- Ward, B.C.; Pederson, N. Infectious peritonitis in cats. J. Am. Vet. Med. Assoc. 1969, 154, 26–35. [Google Scholar]
- Fabricant, C.G.; King, J.M.; Gaskin, J.M.; Gillespie, J.H. Isolation of a virus from a female cat with urolithiasis. J. Am. Vet. Med. Assoc. 1971, 158, 200–201. [Google Scholar]
- Fabricant, C.G.; Rich, L.J.; Gillespie, J.H. Feline viruses. Xi. Isolation of a virus similar to a myxovirus from cats in which urolithiasis was experimentally induced. Cornell. Vet. 1969, 59, 667–672. [Google Scholar]
- Martens, J.G.; McConnell, S.; Swanson, C.L. The role of infectious agents in naturally occurring feline urologic syndrome. Vet. Clin. North Am. Small Anim. Pract. 1984, 14, 503–511. [Google Scholar] [CrossRef]
- Gaskell, R.M.; Gaskell, C.J.; Page, W.; Dennis, P.; Voyle, C.A. Studies on a possible viral aetiology for the feline urological syndrome. Vet. Rec. 1979, 105, 243–247. [Google Scholar] [CrossRef]
- Kruger, J.M.; Osborne, C.A. The role of viruses in feline lower urinary tract disease. J. Vet. Intern. Med. 1990, 4, 71–78. [Google Scholar] [CrossRef]
- McKissick, G.E.; Lamont, P.H. Characteristics of a virus isolated from a feline fibrosarcoma. J. Virol. 1970, 5, 247–257. [Google Scholar]
- Marino, C.L.; Lascelles, B.D.; Vaden, S.L.; Gruen, M.E.; Marks, S.L. Prevalence and classification of chronic kidney disease in cats randomly selected from four age groups and in cats recruited for degenerative joint disease studies. J. Feline Med. Surg. 2014, 16, 465–472. [Google Scholar] [CrossRef]
- Hamilton, J.B.; Hamilton, R.S.; Mestler, G.E. Duration of life and causes of death in domestic cats: Influence of sex, gonadectomy, and inbreeding. J. Gerontol. 1969, 24, 427–437. [Google Scholar] [CrossRef]
- O’Neill, D.G.; Church, D.B.; McGreevy, P.D.; Thomson, P.C.; Brodbelt, D.C. Longevity and mortality of cats attending primary care veterinary practices in england. J. Feline Med. Surg. 2015, 17, 125–133. [Google Scholar] [CrossRef]
- Reynolds, B.S.; Lefebvre, H.P. Feline ckd: Pathophysiology and risk factors–what do we know? J. Feline Med. Surg. 2013, 15, 3–14. [Google Scholar] [CrossRef]
- Bartges, J.W. Chronic kidney disease in dogs and cats. Vet. Clin. N. Am-Small Anim. Pract. 2012, 42, 669–692. [Google Scholar] [CrossRef]
- Brown, C.A.; Elliott, J.; Schmiedt, C.W.; Brown, S.A. Chronic kidney disease in aged cats: Clinical features, morphology, and proposed pathogeneses. Vet. Pathol. 2016, 53, 309–326. [Google Scholar] [CrossRef]
- McLeland, S.M.; Cianciolo, R.E.; Duncan, C.G.; Quimby, J.M. A comparison of biochemical and histopathologic staging in cats with chronic kidney disease. Vet. Pathol. 2015, 52, 524–534. [Google Scholar] [CrossRef]
- Poli, A.; Tozon, N.; Guidi, G.; Pistello, M. Renal alterations in feline immunodeficiency virus (fiv)-infected cats: Anatural model of lentivirus-induced renal disease changes. Viruses 2012, 4, 1372–1389. [Google Scholar] [CrossRef]
- Baxter, K.J.; Levy, J.K.; Edinboro, C.H.; Vaden, S.L.; Tompkins, M.B. Renal disease in cats infected with feline immunodeficiency virus. J. Vet. Intern. Med. 2012, 26, 238–243. [Google Scholar] [CrossRef]
- Poli, A.; Abramo, F.; Taccini, E.; Guidi, G.; Barsotti, P.; Bendinelli, M.; Malvaldi, G. Renal involvement in feline immunodeficiency virus infection: A clinicopathological study. Nephron 1993, 64, 282–288. [Google Scholar] [CrossRef]
- Glick, A.D.; Horn, R.G.; Holscher, M. Characterization of feline glomerulonephritis associated with viral-induced hematopoietic neoplasms. Am. J. Pathol. 1978, 92, 321–332. [Google Scholar]
- Löchelt, M.; Romen, F.; Bastone, P.; Muckenfuss, H.; Kirchner, N.; Kim, Y.B.; Truyen, U.; Rosler, U.; Battenberg, M.; Saib, A.; et al. The antiretroviral activity of APOBEC3 is inhibited by the foamy virus accessory Bet protein. Proc. Natl. Acad. Sci. USA 2005, 102, 7982–7987. [Google Scholar] [CrossRef]
- Reed, L.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Arzi, B.; Kol, A.; Murphy, B.; Walker, N.J.; Wood, J.A.; Clark, K.; Verstraete, F.J.M.; Borjesson, D.L. Feline foamy virus adversely affects feline mesenchymal stem cell culture and expansion: Implications for animal model development. Stem Cells Dev. 2014, 24, 814–823. [Google Scholar] [CrossRef]
- Society, I.R.I. Iris Staging of Ckd. Available online: http://www.iris-kidney.com/guidelines/staging.html (accessed on 28 May 2016).
- Lee, J.S.; Mackie, R.S.; Harrison, T.; Shariat, B.; Kind, T.; Kehl, T.; Löchelt, M.; Boucher, C.; VandeWoude, S. Targeted enrichment for pathogen detection and characterization in three felid species. J. Clin. Microbiol. 2017, 55, 1658–1670. [Google Scholar] [CrossRef]
- Cianciolo, R.E.; Brown, C.A.; Mohr, F.C.; Spangler, W.L.; Aresu, L.; van der Lugt, J.J.; Jansen, J.H.; James, C.; Clubb, F.J.; Lees, G.E. Pathologic evaluation of canine renal biopsies: Methods for identifying features that differentiate immune-mediated glomerulonephritides from other categories of glomerular diseases. J. Vet. Intern. Med. 2013, 27 (Suppl. S1), S10–S18. [Google Scholar] [CrossRef]
- ScyMed. Medicalc: Blood Urea Nitrogen (Unit Conversion). Available online: http://www.scymed.com/en/smnxps/psxff047_c.htm (accessed on 24 June 2019).
- ScyMed. Medicalc: Creatinine (Unit Conversion). Available online: http://www.scymed.com/en/smnxps/psxdf212_c.htm (accessed on 24 June 2019).
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Delignette-Muller, M.L.; Dutang, C. fitdistrplus: An R package for fitting distributions. J. Stat. Softw. 2015, 64, 1–34. [Google Scholar] [CrossRef]
- Brooks, M.; Kristensen, K.; van Benthem, K.; Magnusson, A.; Berg, C.; Nielsen, A.; Skaug, H.; Mächler, M.; Bolker, B. Glmmtmb balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- Epitools: Epidemiology Tools. Available online: https://rdrr.io/cran/epitools/ (accessed on 15 July 2019).
- Ledesma-Feliciano, C.; Department of Microbiology, Immunology, and Pathology, College of Veterinary Medicine and Biomedical Sciences, Colorado State University, Fort Collins, CO, USA. FFV-inoculated and control cat complete blood count and serum chemistry data. 2014. [Google Scholar]
- Cowgill, L.D.; Polzin, D.J.; Elliott, J.; Nabity, M.B.; Segev, G.; Grauer, G.F.; Brown, S.; Langston, C.; van Dongen, A.M. Is progressive chronic kidney disease a slow acute kidney injury? Vet. Clin. Small Anim. Pract. 2016, 46, 995–1013. [Google Scholar] [CrossRef]
- Ledesma-Feliciano, C.; Department of Microbiology, Immunology, and Pathology, College of Veterinary Medicine and Biomedical Sciences, Colorado State University, Fort Collins, CO, USA. FFV-inoculated and control cat urinalysis and serum creatinine data. 2014. [Google Scholar]
- Cavalcante, L.T.F.; Muniz, C.P.; Jia, H.; Augusto, A.M.; Troccoli, F.; Medeiros, S.O.; Dias, C.G.A.; Switzer, W.M.; Soares, M.A.; Santos, A.F. Clinical and molecular features of feline foamy virus and feline leukemia virus co-infection in naturally-infected cats. Viruses 2018, 10, 702. [Google Scholar] [CrossRef]
- Brooks, J.I.; Merks, H.W.; Fournier, J.; Boneva, R.S.; Sandstrom, P.A. Characterization of blood-borne transmission of simian foamy virus. Transfusion 2007, 47, 162–170. [Google Scholar] [CrossRef]
- Murray, S.M.; Linial, M.L. Foamy virus infection in primates. J. Med. Primatol. 2006, 35, 225–235. [Google Scholar] [CrossRef]
- Soliven, K.; Wang, X.; Small, C.T.; Feeroz, M.M.; Lee, E.-G.; Craig, K.L.; Hasan, K.; Engel, G.a.; Jones-Engel, L.; Matsen, F.a.; et al. Simian foamy virus infection of rhesus macaques in bangladesh: Relationship of latent proviruses and transcriptionally active viruses. J. Virol. 2013, 87, 13628–13639. [Google Scholar] [CrossRef]
- Falcone, V.; Leupold, J.; Clotten, J.; Urbanyi, E.; Herchenroder, O.; Spatz, W.; Volk, B.; Bohm, N.; Toniolo, A.; Neumann-Haefelin, D.; et al. Sites of simian foamy virus persistence in naturally infected african green monkeys: Latent provirus is ubiquitous, whereas viral replication is restricted to the oral mucosa. Virology 1999, 257, 7–14. [Google Scholar] [CrossRef]
- Gray, F.; Lescs, M.C.; Keohane, C.; Paraire, F.; Marc, B.; Durigon, M.; Gherardi, R. Early brain changes in hiv infection: Neuropathological study of 11 hiv seropositive, non-aids cases. J. Neuropathol. Exp. Neurol. 1992, 51, 177–185. [Google Scholar] [CrossRef]
- Bell, J.E. The neuropathology of adult hiv infection. Rev. Neurol. 1998, 154, 816–829. [Google Scholar]
- Torres, L.; Noel, R.J., Jr. Astrocytic expression of hiv-1 viral protein r in the hippocampus causes chromatolysis, synaptic loss and memory impairment. J. Neuroinflamm. 2014, 11, 53. [Google Scholar] [CrossRef]
- Wilcox, G.E.; Flower, R.L.; Cook, R.D. Recovery of viral agents from the central nervous system of cats. Vet. Microbiol. 1984, 9, 355–366. [Google Scholar] [CrossRef]
- Wharram, B.L.; Goyal, M.; Wiggins, J.E.; Sanden, S.K.; Hussain, S.; Filipiak, W.E.; Saunders, T.L.; Dysko, R.C.; Kohno, K.; Holzman, L.B.; et al. Podocyte depletion causes glomerulosclerosis: Diphtheria toxin-induced podocyte depletion in rats expressing human diphtheria toxin receptor transgene. J. Am. Soc. Nephrol. 2005, 16, 2941–2952. [Google Scholar] [CrossRef]
- Chiswell, D.J.; Pringle, C.R. Feline syncytium-forming virus: Identification of a virion associated reverse transcriptase and electron microscopical observations of infected cells. J. Gen. Virol. 1979, 43, 429–434. [Google Scholar] [CrossRef]
- Alke, A.; Schwantes, A.; Kido, K.; Flotenmeyer, M.; Flugel, R.M.; Löchelt, M. The bet gene of feline foamy virus is required for virus replication. Virology 2001, 287, 310–320. [Google Scholar] [CrossRef]
- Cook, R.D. Observations on focal demyelinating lesions in cat optic nerves. Neuropathol. Appl. Neurobiol. 1979, 5, 395–404. [Google Scholar] [CrossRef]
- Toribio, J.A.; Norris, J.M.; White, J.D.; Dhand, N.K.; Hamilton, S.A.; Malik, R. Demographics and husbandry of pet cats living in sydney, australia: Results of cross-sectional survey of pet ownership. J. Feline Med. Surg. 2009, 11, 449–461. [Google Scholar] [CrossRef]
- Johnston, L.; Szczepanski, J.; McDonagh, P. Demographics, lifestyle and veterinary care of cats in australia and new zealand. J. Feline Med. Surg. 2017, 19, 1199–1205. [Google Scholar] [CrossRef]
- White, J.D.; Norris, J.M.; Baral, R.M.; Malik, R. Naturally-occurring chronic renal disease in australian cats: A prospective study of 184 cases. Aust. Vet. J. 2006, 84, 188–194. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | Population | Cluster of Differentiation |
|---|---|---|
| CBC | WBC | - |
| Monocytes | - | |
| Lymphocytes | - | |
| Neutrophils | - | |
| Flow Cytometry (Panel A) | Th 1 lymphocytes | CD4+ |
| Activation | CD4+CD25+ | |
| Activation | CD4+CD134+ | |
| Apoptosis | CD4+Fas+ | |
| Tc 2 lymphocytes | CD8+ | |
| Activation | CD8+CD25+ | |
| Activation | CD8+CD134+ | |
| Apoptosis | CD8+Fas+ | |
| Double-positive T cells | CD4+CD8+ | |
| Flow Cytometry (Panel B) | B cells | CD21+ |
| Activation | CD21+MHCII+ | |
| Apoptosis | CD21+Fas+ | |
| NK cells | CD56+ | |
| Activation | CD56+MHCII+ | |
| Apoptosis | CD56+Fas+ | |
| Monocytes | CD14+ | |
| Activation | CD14+MHCII+ | |
| Apoptosis | CD14+Fas+ | |
| WBC activation | MHCII+ | |
| WBC apoptosis | Fas+ |
| Assay | Days Tested | Summary of Findings |
|---|---|---|
| Saliva qPCR (RNA) | 36, 42, 49, 56, 63, 86, 112, 119, 126, 133, 142, 147, 154, 161, 168, 176 | D133 saliva sample was FFV DNA-negative. |
| Plasma qPCR (DNA) | 42, 86, 142, 176 | FFV not detected. |
| Plasma qPCR (RNA) | 15, 56, 112 | FFV not detected. |
| Tissue qPCR (DNA) | 176 | Virus shows primarily lymphoid tropism. Cat FFV3 had expanded tropism to other lymphoid and non-lymphoid tissues (Table 3). FFV not detected in cat FFV4’s tissues. |
| CBC, Chemistry | −21, 0, 3, 7, 10, 15, 21, 28, 42, 56, 63, 70, 77, 86, 98, 112, 126, 142, 154, 168, 176 | Not indicative of disease. |
| PBMC Phenotype | −21, 0, 3, 7, 10, 15, 21, 28, 42, 56, 70, 86, 112, 142, 168 | Significantly increased populations in FFV cats included monocytes and CD21+MHCII+, while CD8+CD25+, CD8+CD134+, CD8+FAS+, CD56+, and CD56+MHCII+ cells were decreased. |
| BUN, Creatinine | −21, 0, 7, 15, 21, 28, 42, 56, 63, 70, 77, 86, 98, 112, 126, 142, 154, 168, 176 | While BUN remained within normal limits for all cats, values were significantly increased in FFV group cats compared to naïve on bolded days. Creatinine values were within normal ranges and did not rise above 1.8 mg/dl. |
| Urinalysis | −21, 3, 7, 10, 42, 56, 63, 70, 86, 91, 98, 112, 122, 142, 156, 171, 176 | USG > 1.035 for all cats. Urinalysis and urine sediment were unremarkable. |
| UPC Ratio | 36, 70, 86, 91, 98, 122, 142, 156, 171, 176 | UPC ratio was 0.1 (normal) for all cats, except cat FFV3 where it increased to 0.2 (borderline proteinuric) on 122 and 142 days pi, during highest PBMC proviral load for this cat [10]. |
| Tissue | N4 | FFV1 | FFV2 | FFV3 | FFV4 | FFV5 | Total Cats |
|---|---|---|---|---|---|---|---|
| Salivary gland | - | - | - | - | - | - | 0 |
| Tongue | - | - | - | - | - | - | 0 |
| Oral Mucosa | - | - | - | 2.10 × 102 | - | - | 1 |
| Tonsil | - | 2.41 × 102 | 4.03 × 102 | - | - | 5.10 × 102 | 3 |
| Prescapular LN | - | 5.89 × 103 | - | - | - | 4.96 × 102 | 2 |
| Submandibular LN | - | 3.35 × 102 | 2.26 × 102 | - | - | 5.78 × 102 | 3 |
| Retropharyngeal LN | - | 1.86 × 102 | 1.19 × 102 | 1.49 × 102 | - | 2.35 × 102 | 4 |
| Mesenteric LN | - | - | - | - | - | - | 0 |
| Thymus | - | - | - | 3.48 × 102 | - | - | 1 |
| Spleen | - | 5.93 × 102 | 3.11 × 102 | 2.10 × 102 | - | 3.35 × 102 | 4 |
| Ileum | - | - | - | - | - | - | 0 |
| Bone marrow | - | - | - | 6.10 × 102 | - | - | 1 |
| Kidney | - | - | - | - | - | - | 0 |
| Muscle | - | - | - | - | - | - | 0 |
| Analysis | Finding | N4 | FFV1 | FFV2 | FFV3 | FFV4 | FFV5 | Total Cats |
|---|---|---|---|---|---|---|---|---|
| Histology | Tubular degeneration (+/− atrophy) | - | + | + | - | - | - | 2 |
| TEM | Podocyte effacement | NT | +/+ | ++ | ++ | + | +++ | 5 |
| Cytoplasmic electron dense figures | NT | - | + | + | + | + | 4 | |
| Cytoplasmic myelin figures | NT | + | + | - | + | - | 3 | |
| Cytoplasmic vacuolization | NT | - | + | + | - | - | 2 | |
| Wrinkled glomerular capillary walls | NT | - | - | - | - | + | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ledesma-Feliciano, C.; Troyer, R.M.; Zheng, X.; Miller, C.; Cianciolo, R.; Bordicchia, M.; Dannemiller, N.; Gagne, R.; Beatty, J.; Quimby, J.; et al. Feline Foamy Virus Infection: Characterization of Experimental Infection and Prevalence of Natural Infection in Domestic Cats with and without Chronic Kidney Disease. Viruses 2019, 11, 662. https://doi.org/10.3390/v11070662
Ledesma-Feliciano C, Troyer RM, Zheng X, Miller C, Cianciolo R, Bordicchia M, Dannemiller N, Gagne R, Beatty J, Quimby J, et al. Feline Foamy Virus Infection: Characterization of Experimental Infection and Prevalence of Natural Infection in Domestic Cats with and without Chronic Kidney Disease. Viruses. 2019; 11(7):662. https://doi.org/10.3390/v11070662
Chicago/Turabian StyleLedesma-Feliciano, Carmen, Ryan M. Troyer, Xin Zheng, Craig Miller, Rachel Cianciolo, Matteo Bordicchia, Nicholas Dannemiller, Roderick Gagne, Julia Beatty, Jessica Quimby, and et al. 2019. "Feline Foamy Virus Infection: Characterization of Experimental Infection and Prevalence of Natural Infection in Domestic Cats with and without Chronic Kidney Disease" Viruses 11, no. 7: 662. https://doi.org/10.3390/v11070662
APA StyleLedesma-Feliciano, C., Troyer, R. M., Zheng, X., Miller, C., Cianciolo, R., Bordicchia, M., Dannemiller, N., Gagne, R., Beatty, J., Quimby, J., Löchelt, M., & VandeWoude, S. (2019). Feline Foamy Virus Infection: Characterization of Experimental Infection and Prevalence of Natural Infection in Domestic Cats with and without Chronic Kidney Disease. Viruses, 11(7), 662. https://doi.org/10.3390/v11070662