Evolutionary and Functional Diversity of the 5′ Untranslated Region of Enterovirus D68: Increased Activity of the Internal Ribosome Entry Site of Viral Strains during the 2010s


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Genomic Data
2.2. Phylogenetic Analysis
2.3. Secondary Structure
2.4. Cell Lines
2.5. Viruses
2.6. Plasmids
2.7. Luciferase Assay
2.8. Viral Growth Assay
2.9. Statistical Analysis
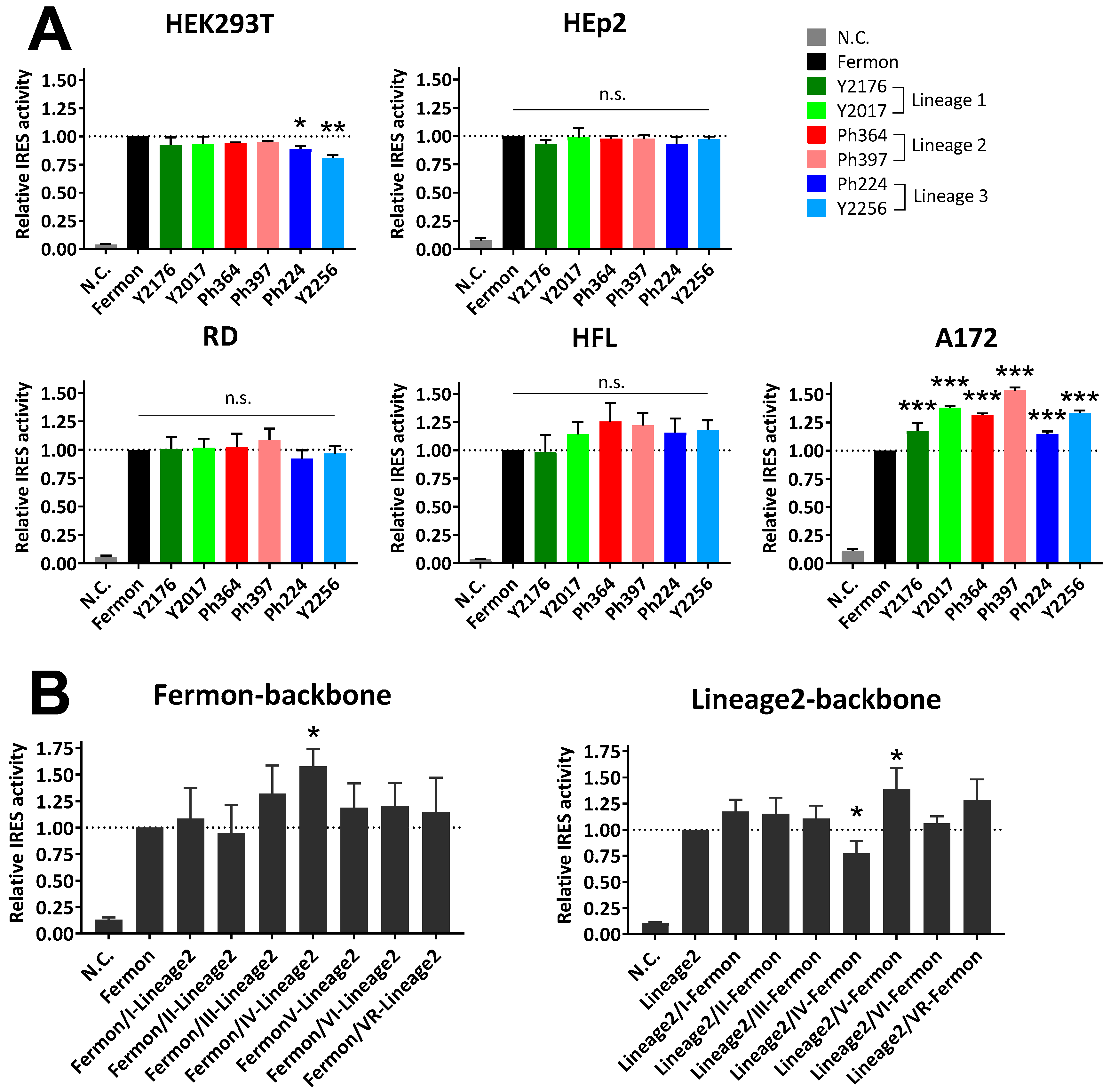
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tapparel, C.; Siegrist, F.; Petty, T.J.; Kaiser, L. Picornavirus and enterovirus diversity with associated human diseases. Infect. Genet. Evol. 2013, 14, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Rossmann, M.G.; Arnold, E.; Erickson, J.W.; Frankenberger, E.A.; Griffith, J.P.; Hecht, H.J.; Johnson, J.E.; Kamer, G.; Luo, M.; Mosser, A.G. Structure of a human common cold virus and functional relationship to other picornaviruses. Nature 1985, 317, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Pulli, T.; Lankinen, H.; Roivainen, M.; Hyypiä, T. Antigenic sites of coxsackievirus A9. Virology 1998, 240, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Imamura, T.; Okamoto, M.; Nakakita, S.; Suzuki, A.; Saito, M.; Tamaki, R.; Lupisan, S.; Roy, C.N.; Hiramatsu, H.; Sugawara, K.; et al. Antigenic and receptor binding properties of enterovirus 68. J. Virol. 2014, 88, 2374–2384. [Google Scholar] [CrossRef] [PubMed]
- Trono, D.; Pelletier, J.; Sonenberg, N.; Baltimore, D. Translation in mammalian cells of a gene linked to the poliovirus 5′ noncoding region. Science 1988, 241, 445–448. [Google Scholar] [CrossRef] [PubMed]
- Dutkiewicz, M.; Stachowiak, A.; Swiatkowska, A.; Ciesiołka, J. Structure and function of RNA elements present in enteroviral genomes. Acta Biochim. Pol. 2016, 63, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Salas, E. The impact of RNA structure on picornavirus IRES activity. Trends Microbiol. 2008, 16, 230–237. [Google Scholar] [CrossRef]
- Yeh, M.T.; Wang, S.W.; Yu, C.K.; Lin, K.H.; Lei, H.Y.; Su, I.J.; Wang, J.R. A single nucleotide in stem loop II of 5′-untranslated region contributes to virulence of Enterovirus 71 in mice. PLoS ONE 2011, 6. [Google Scholar] [CrossRef]
- Gromeier, M.; Alexander, L.; Wimmer, E. Internal ribosomal entry site substitution eliminates neurovirulence in intergeneric poliovirus recombinants. Proc. Natl. Acad. Sci. USA 1996, 93, 2370–2375. [Google Scholar] [CrossRef]
- Rinehart, J.E.; Gómez, R.M.; Roos, R.P. Molecular determinants for virulence in coxsackievirus B1 infection. J. Virol. 1997, 71, 3986–3991. [Google Scholar]
- Centers for Disease Control and Prevention (CDC). Clusters of acute respiratory illness associated with human enterovirus 68—Asia, Europe, and United States, 2008–2010. Morb. Mortal. Wkly. Rep. 2011, 60, 1301–1304. [Google Scholar]
- Holm-Hansen, C.C.; Midgley, S.E.; Fischer, T.K. Global emergence of enterovirus D68: A systematic review. Lancet Infect. Dis. 2016. [Google Scholar] [CrossRef]
- Furuse, Y.; Chaimongkol, N.; Okamoto, M.; Imamura, T.; Saito, M.; Tamaki, R.; Saito, M.; Lupisan, S.P.; Oshitani, H. Molecular epidemiology of enterovirus D68 from 2013 to 2014 in Philippines. J. Clin. Microbiol. 2015, 53, 1015–1018. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cao, J.; Zhang, S.; Lee, A.J.; Sun, G.; Larsen, C.N.; Zhao, H.; Gu, Z.; He, S.; Klem, E.B.; et al. Genetic changes found in a distinct clade of Enterovirus D68 associated with paralysis during the 2014 outbreak. Virus Evol. 2016, 2, vew015. [Google Scholar] [CrossRef] [PubMed]
- Greninger, A.L.; Naccache, S.N.; Messacar, K.; Clayton, A.; Yu, G.; Somasekar, S.; Federman, S.; Stryke, D.; Anderson, C.; Yagi, S.; et al. A novel outbreak enterovirus D68 strain associated with acute flaccid myelitis cases in the USA (2012-14): A retrospective cohort study. Lancet Infect. Dis. 2015, 15, 671–682. [Google Scholar] [CrossRef]
- Yip, C.; Lo, J.; Sridhar, S.; Lung, D.; Luk, S.; Chan, K.-H.; Chan, J.; Cheng, V.; Woo, P.; Yuen, K.-Y.; et al. First Report of a Fatal Case Associated with EV-D68 Infection in Hong Kong and Emergence of an Interclade Recombinant in China Revealed by Genome Analysis. Int. J. Mol. Sci. 2017, 18, 1065. [Google Scholar] [CrossRef] [PubMed]
- Simmonds, P. SSE: A nucleotide and amino acid sequence analysis platform. BMC Res. Notes 2012, 5, 50. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Reuter, J.S.; Mathews, D.H. RNAstructure: Software for RNA secondary structure prediction and analysis. BMC Bioinform. 2010, 11, 129. [Google Scholar] [CrossRef]
- Ikeda, T.; Mizuta, K.; Abiko, C.; Aoki, Y.; Itagaki, T.; Katsushima, F.; Katsushima, Y.; Matsuzaki, Y.; Fuji, N.; Imamura, T.; et al. Acute respiratory infections due to enterovirus 68 in Yamagata, Japan between 2005 and 2010. Microbiol. Immunol. 2012, 56, 139–143. [Google Scholar] [CrossRef]
- Imamura, T.; Suzuki, A.; Lupisan, S.; Okamoto, M.; Aniceto, R.; Egos, R.J.; Daya, E.E.; Tamaki, R.; Saito, M.; Fuji, N.; et al. Molecular Evolution of Enterovirus 68 Detected in the Philippines. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Imamura, T.; Fuji, N.; Suzuki, A.; Tamaki, R.; Saito, M.; Aniceto, R.; Galang, H.; Sombrero, L.; Lupisan, S.; Oshitani, H. Enterovirus 68 among children with severe acute respiratory infection, the Philippines. Emerg. Infect. Dis. 2011, 17, 1430–1435. [Google Scholar] [CrossRef] [PubMed]
- Dierssen, U.; Rehren, F.; Henke-Gendo, C.; Harste, G.; Heim, A. Rapid routine detection of enterovirus RNA in cerebrospinal fluid by a one-step real-time RT-PCR assay. J. Clin. Virol. 2008, 42, 58–64. [Google Scholar] [CrossRef]
- Tokarz, R.; Firth, C.; Madhi, S.A.; Howie, S.R.C.; Wu, W.; Sall, A.A.; Haq, S.; Briese, T.; Ian Lipkin, W. Worldwide emergence of multiple clades of enterovirus 68. J. Gen. Virol. 2012, 93, 1952–1958. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Hassan, F.; Schuster, J.E.; Simenauer, A.; Selvarangan, R.; Halpin, R.A.; Lin, X.; Fedorova, N.; Stockwell, T.B.; Tsan-Yuk Lam, T.; et al. Molecular evolution and intra-clade recombination of Enterovirus D68 during the 2014 outbreak in the United States. J. Virol. 2015, 90, JVI.02418-15. [Google Scholar] [CrossRef]
- Hixon, A.M.; Yu, G.; Leser, J.S.; Yagi, S.; Clarke, P.; Chiu, C.Y.; Tyler, K.L. A mouse model of paralytic myelitis caused by enterovirus D68. PLoS Pathog. 2017, 13. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.M.; Hixon, A.M.; Oldfield, L.M.; Zhang, Y.; Novotny, M.; Wang, W.; Das, S.R.; Shabman, R.S.; Tyler, K.L.; Scheuermann, R.H. Contemporary Circulating Enterovirus D68 Strains Have Acquired the Capacity for Viral Entry and Replication in Human Neuronal Cells. MBio 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Morrey, J.D.; Wang, H.; Hurst, B.L.; Zukor, K.; Siddharthan, V.; van Wettere, A.J.; Sinex, D.G.; Tarbet, E.B. Causation of acute flaccid paralysis by myelitis and myositis in enterovirus-D68 infected mice deficient in interferon αβ/γ receptor deficient mice. Viruses 2018, 10, 33. [Google Scholar] [CrossRef] [PubMed]
- Dyda, A.; Stelzer-Braid, S.; Adam, D.; Chughtai, A.A.; Macintyre, C.R. The association between acute flaccid myelitis (AFM) and enterovirus D68 (EV-D68)—What is the evidence for causation? Eurosurveillance 2018. [Google Scholar] [CrossRef]
- Hixon, A.M.; Clarke, P.; Tyler, K.L. Contemporary circulating enterovirus D68 strains infect and undergo retrograde axonal transport in spinal motor neurons independent of sialic acid. J. Virol. 2019. [Google Scholar] [CrossRef]
- Sean, P.; Nguyen, J.H.C.; Semler, B.L. Altered interactions between stem-loop IV within the 5′ noncoding region of coxsackievirus RNA and poly(rC) binding protein 2: Effects on IRES-mediated translation and viral infectivity. Virology 2009, 389, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Blyn, L.B.; Chen, R.; Semler, B.L.; Ehrenfeld, E. Host cell proteins binding to domain IV of the 5′ noncoding region of poliovirus RNA. J. Virol. 1995, 69, 4381–4389. [Google Scholar] [PubMed]
- Gamarnik, A.; Andino, R. Two functional complexes formed by KH domain containing proteins with the 5′ noncoding region of poliovirus RNA. RNA 1997, 3, 882–892. [Google Scholar] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furuse, Y.; Chaimongkol, N.; Okamoto, M.; Oshitani, H. Evolutionary and Functional Diversity of the 5′ Untranslated Region of Enterovirus D68: Increased Activity of the Internal Ribosome Entry Site of Viral Strains during the 2010s. Viruses 2019, 11, 626. https://doi.org/10.3390/v11070626
Furuse Y, Chaimongkol N, Okamoto M, Oshitani H. Evolutionary and Functional Diversity of the 5′ Untranslated Region of Enterovirus D68: Increased Activity of the Internal Ribosome Entry Site of Viral Strains during the 2010s. Viruses. 2019; 11(7):626. https://doi.org/10.3390/v11070626
Chicago/Turabian StyleFuruse, Yuki, Natthawan Chaimongkol, Michiko Okamoto, and Hitoshi Oshitani. 2019. "Evolutionary and Functional Diversity of the 5′ Untranslated Region of Enterovirus D68: Increased Activity of the Internal Ribosome Entry Site of Viral Strains during the 2010s" Viruses 11, no. 7: 626. https://doi.org/10.3390/v11070626
APA StyleFuruse, Y., Chaimongkol, N., Okamoto, M., & Oshitani, H. (2019). Evolutionary and Functional Diversity of the 5′ Untranslated Region of Enterovirus D68: Increased Activity of the Internal Ribosome Entry Site of Viral Strains during the 2010s. Viruses, 11(7), 626. https://doi.org/10.3390/v11070626