Genetic Characterization of a Recombinant Myxoma Virus in the Iberian Hare (Lepus granatensis)

 ,
,  , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
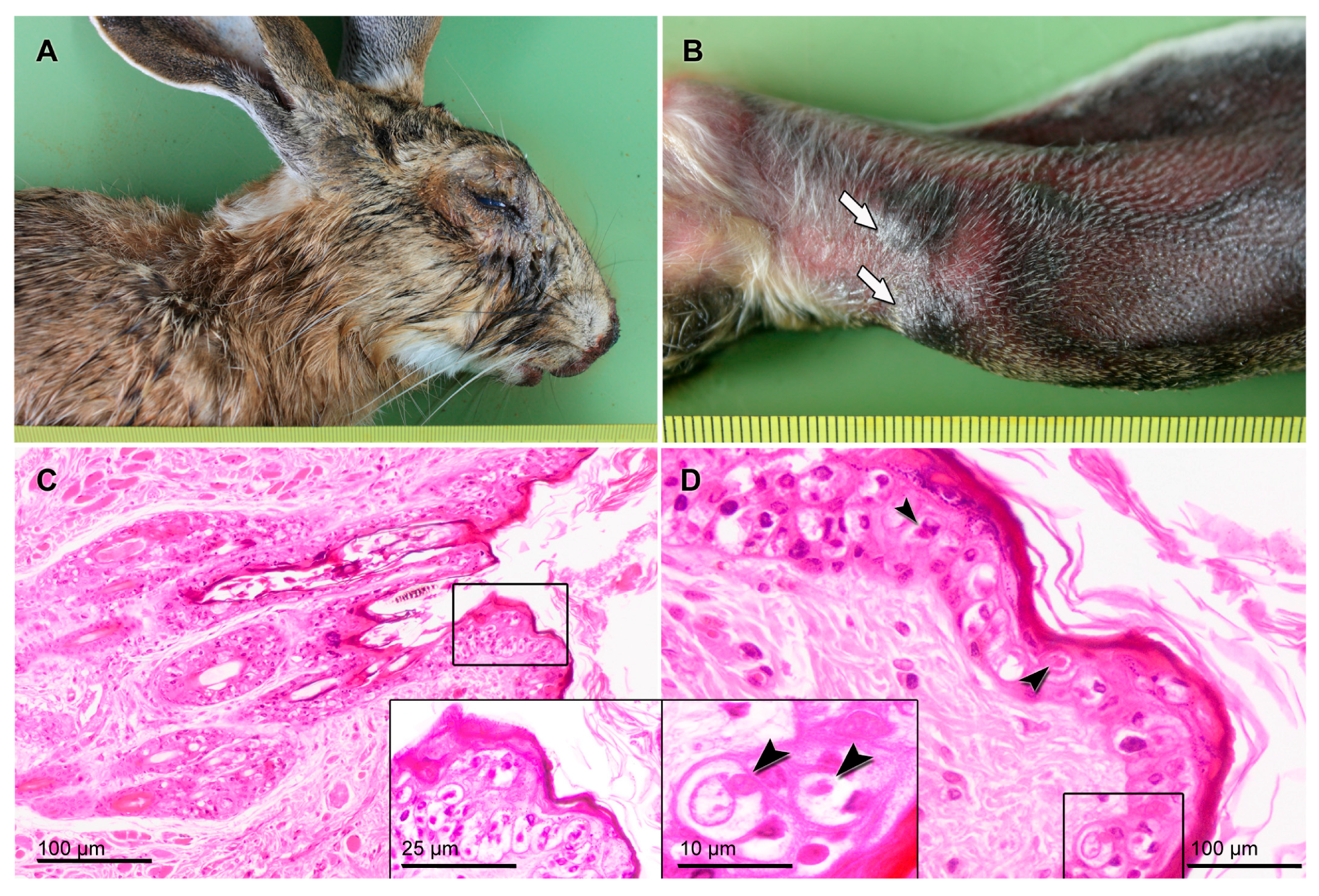
2.1. Sampling and Pathology
2.2. Cell Lines
2.2.1. Isolation, Replication, and Purification of the New Myxoma Virus (MYXV Toledo)
2.2.2. Viral Nucleic Acid Extraction, Illumina Sequencing, and De novo Assembly of the Genome
2.3. Genome Analysis
3. Results and Discussion
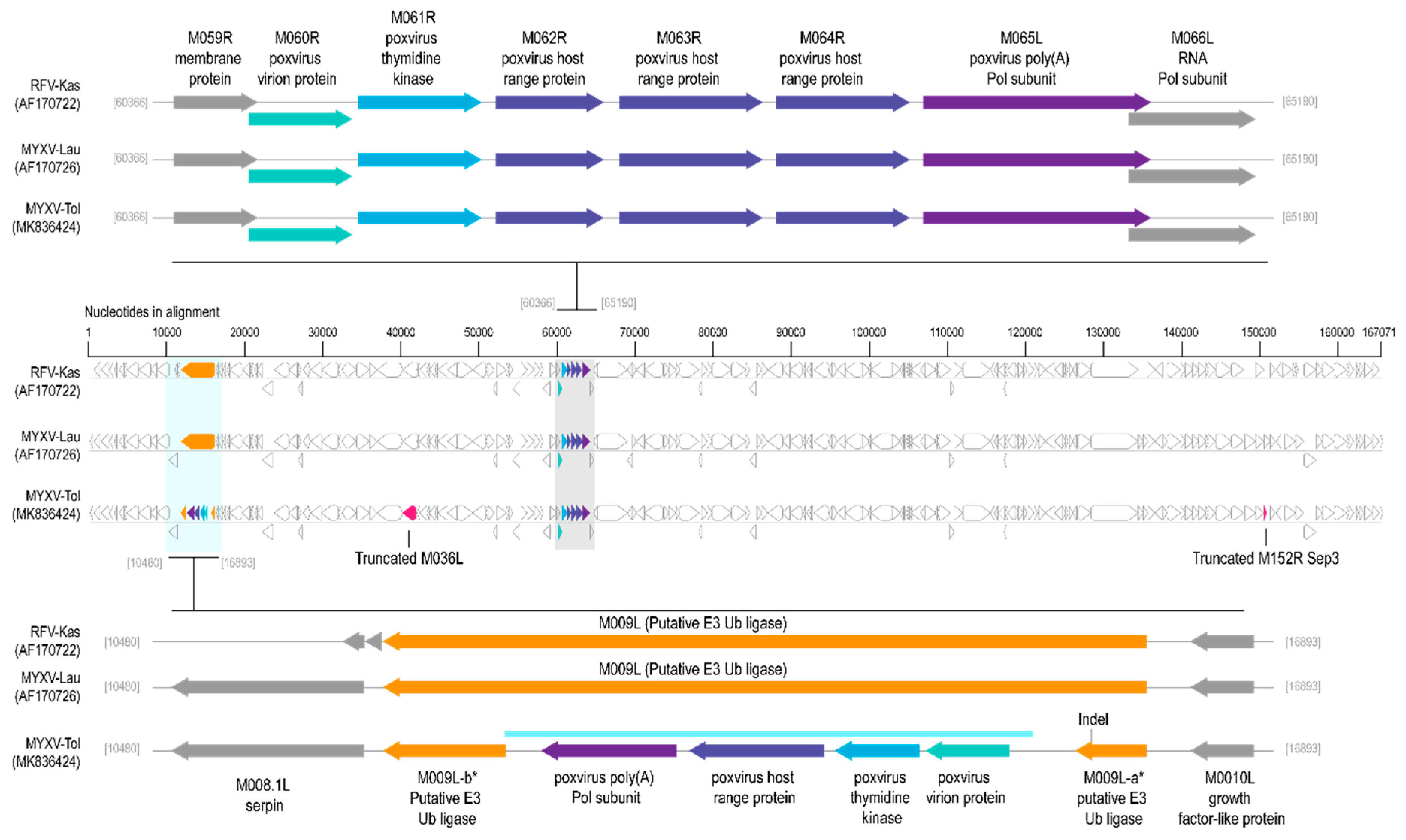
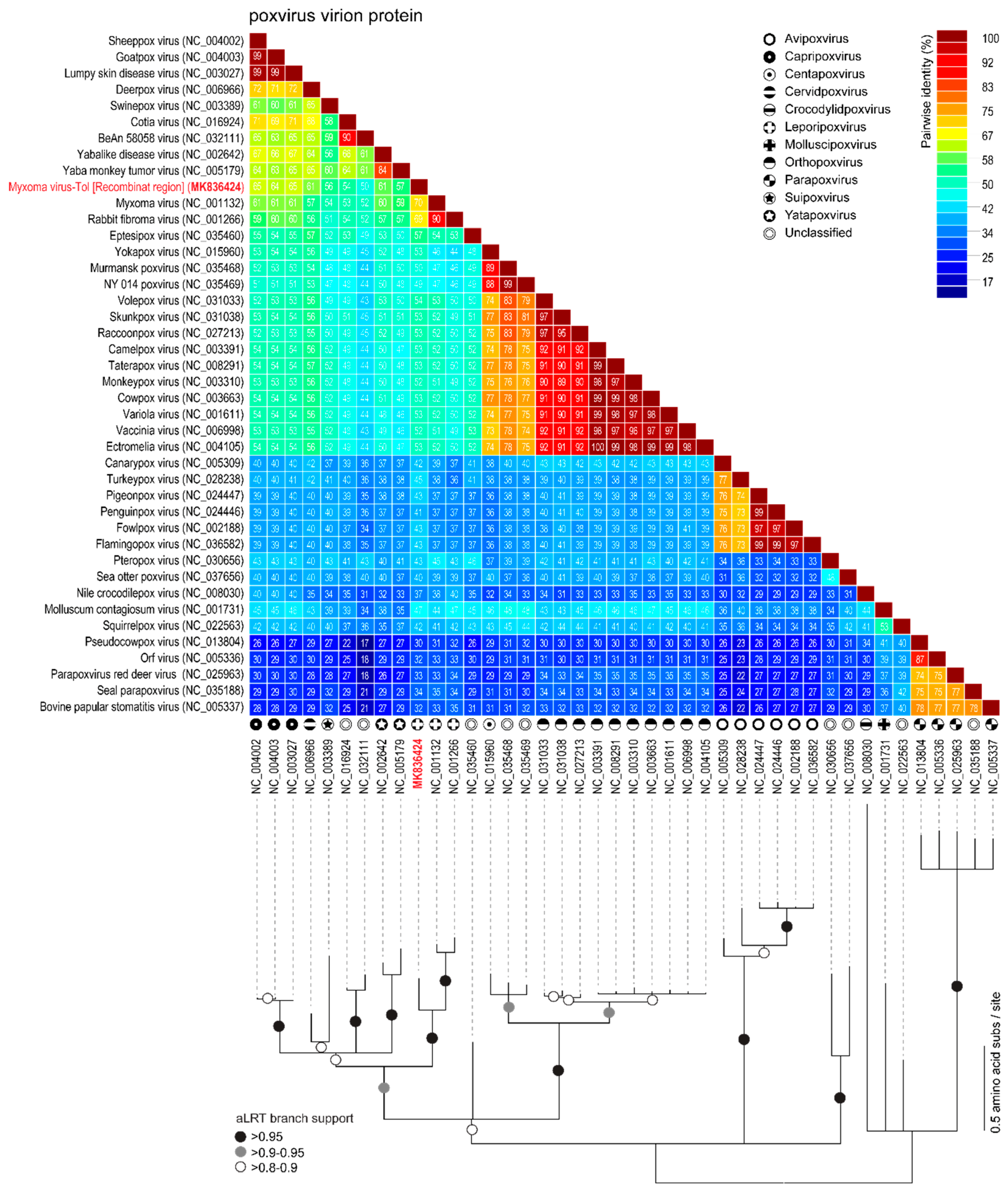
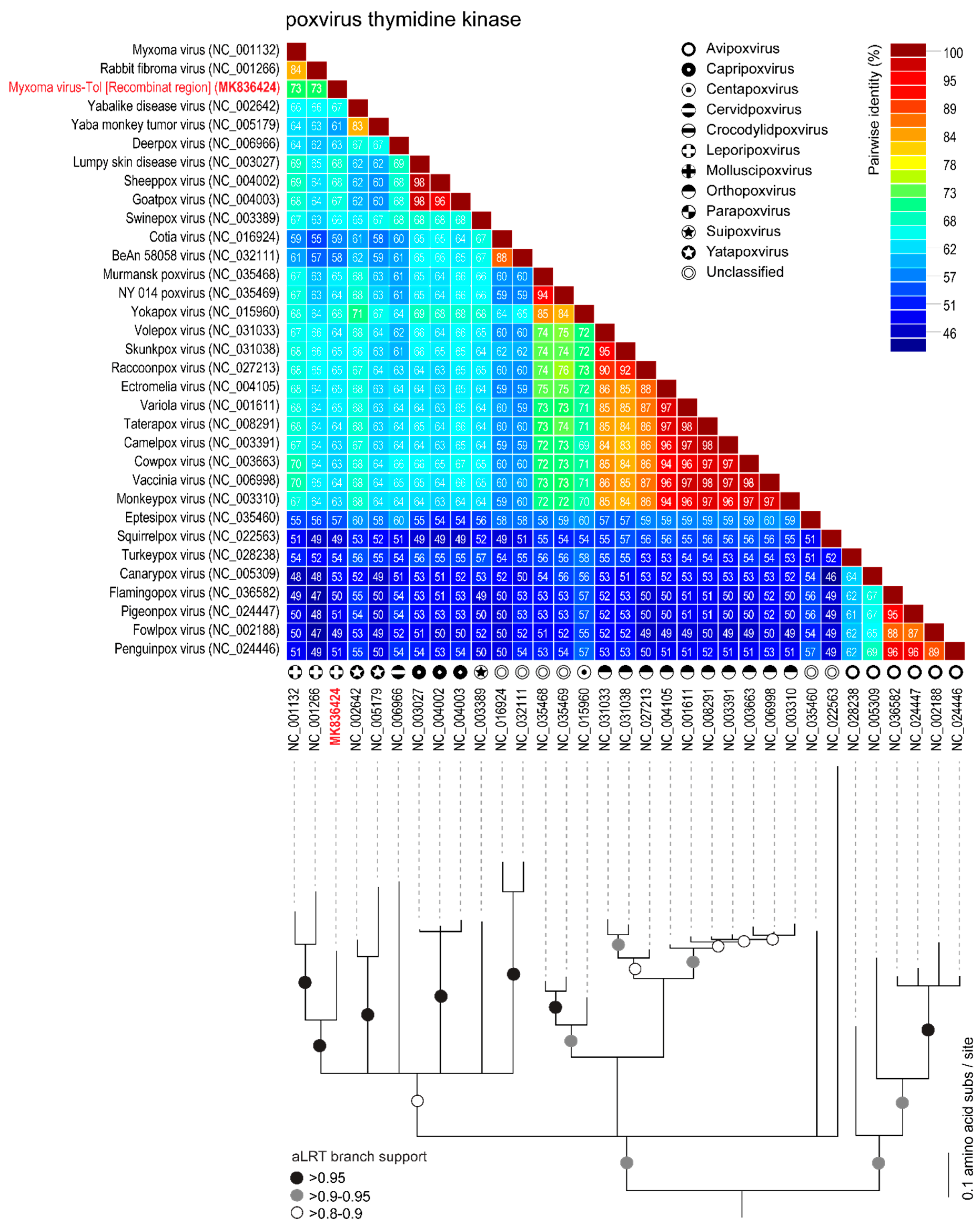
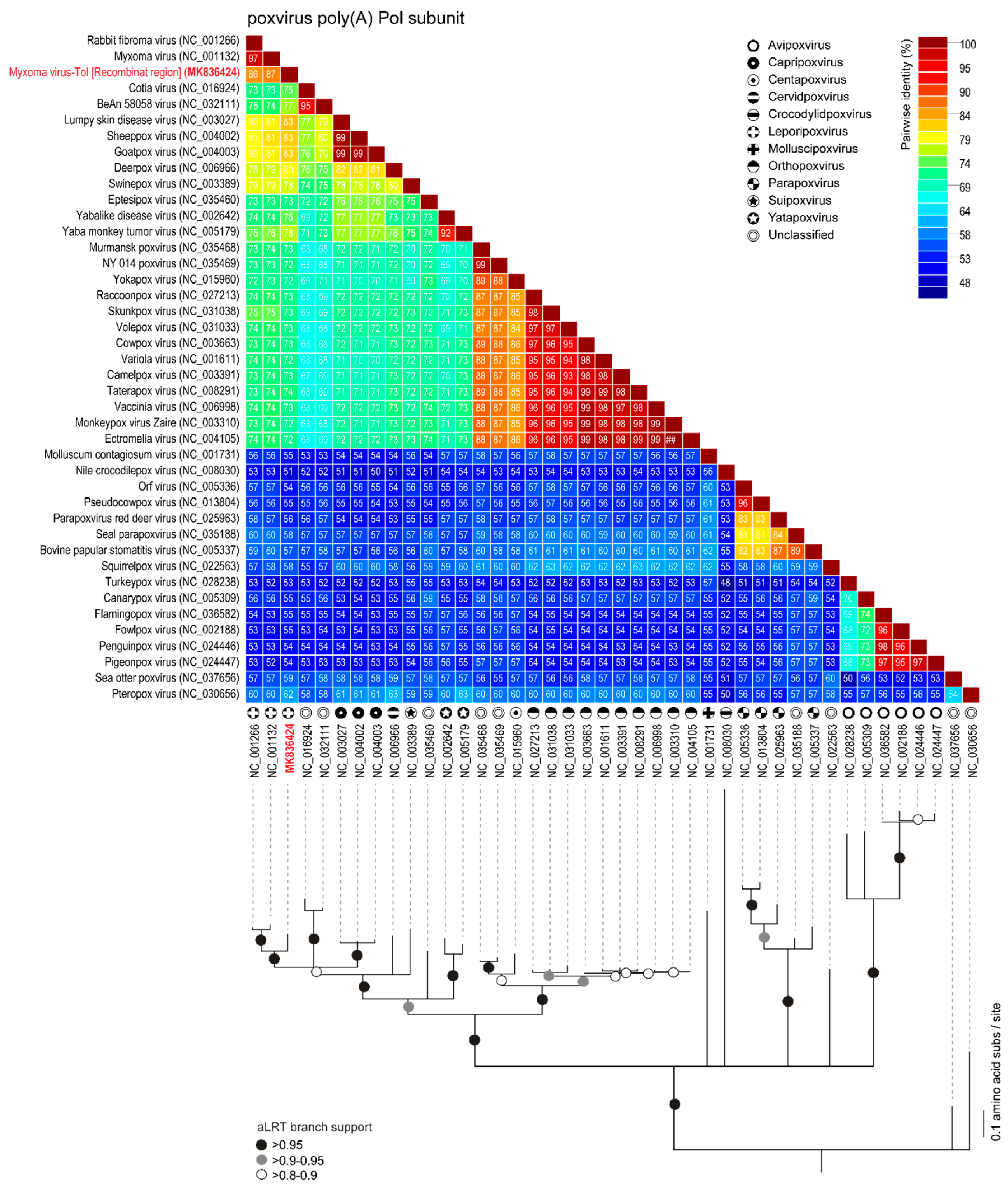
3.1. Comparison of MYXV Lausanne variant with the Newly Discovered MYXV Toledo Variant and RFV Kasza
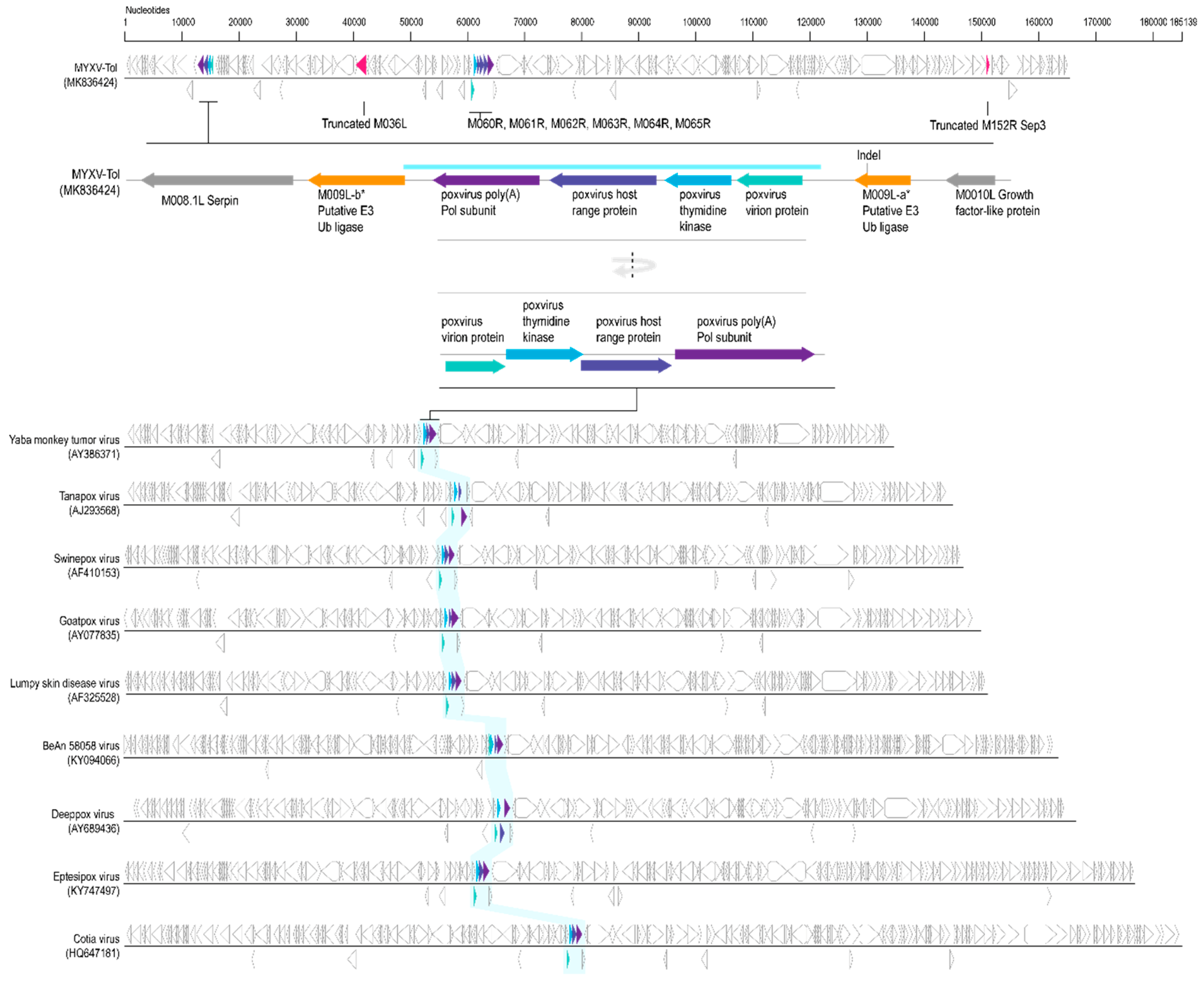
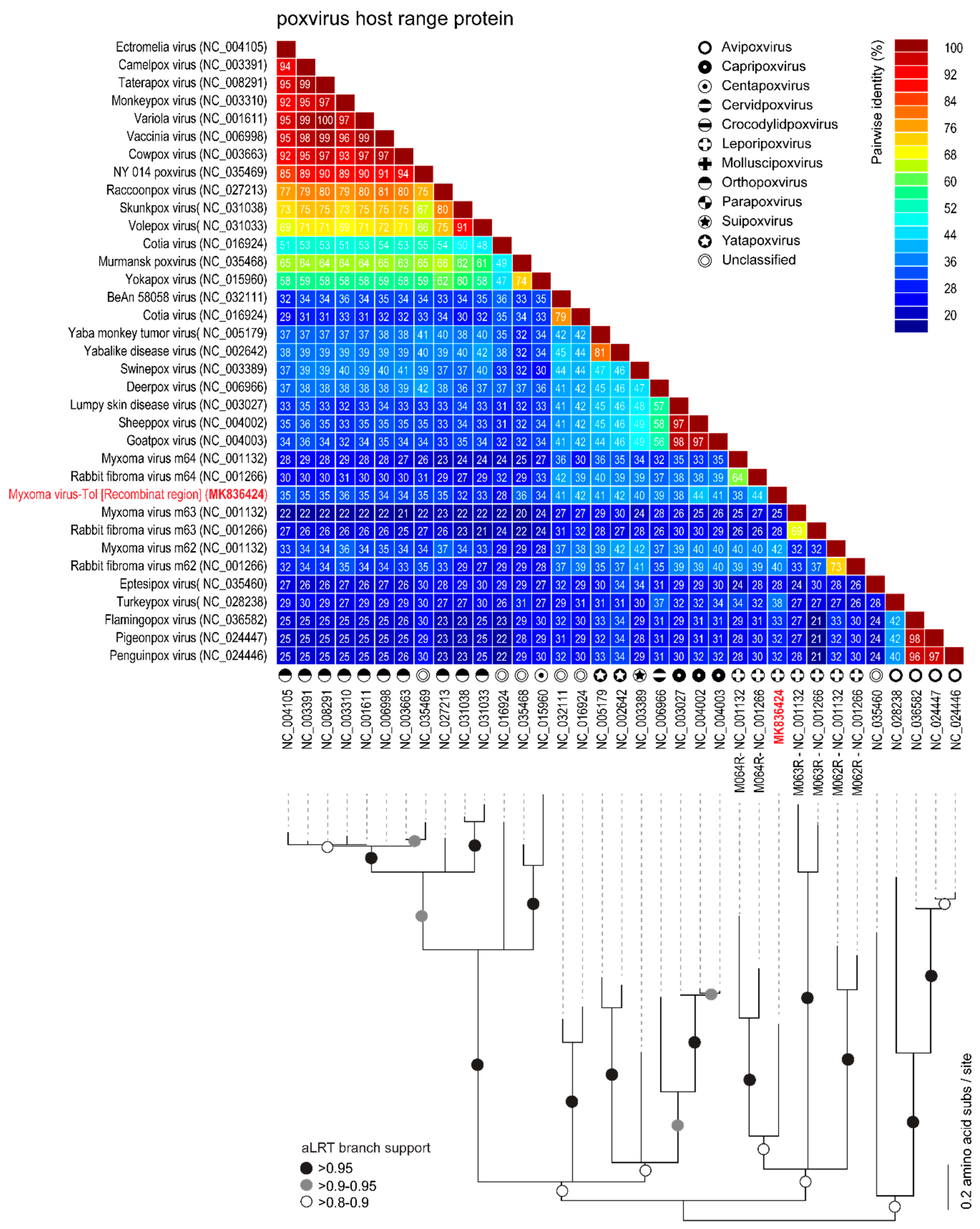
3.2. Viral Genes Disrupted in the New MYXV-Tol Isolate
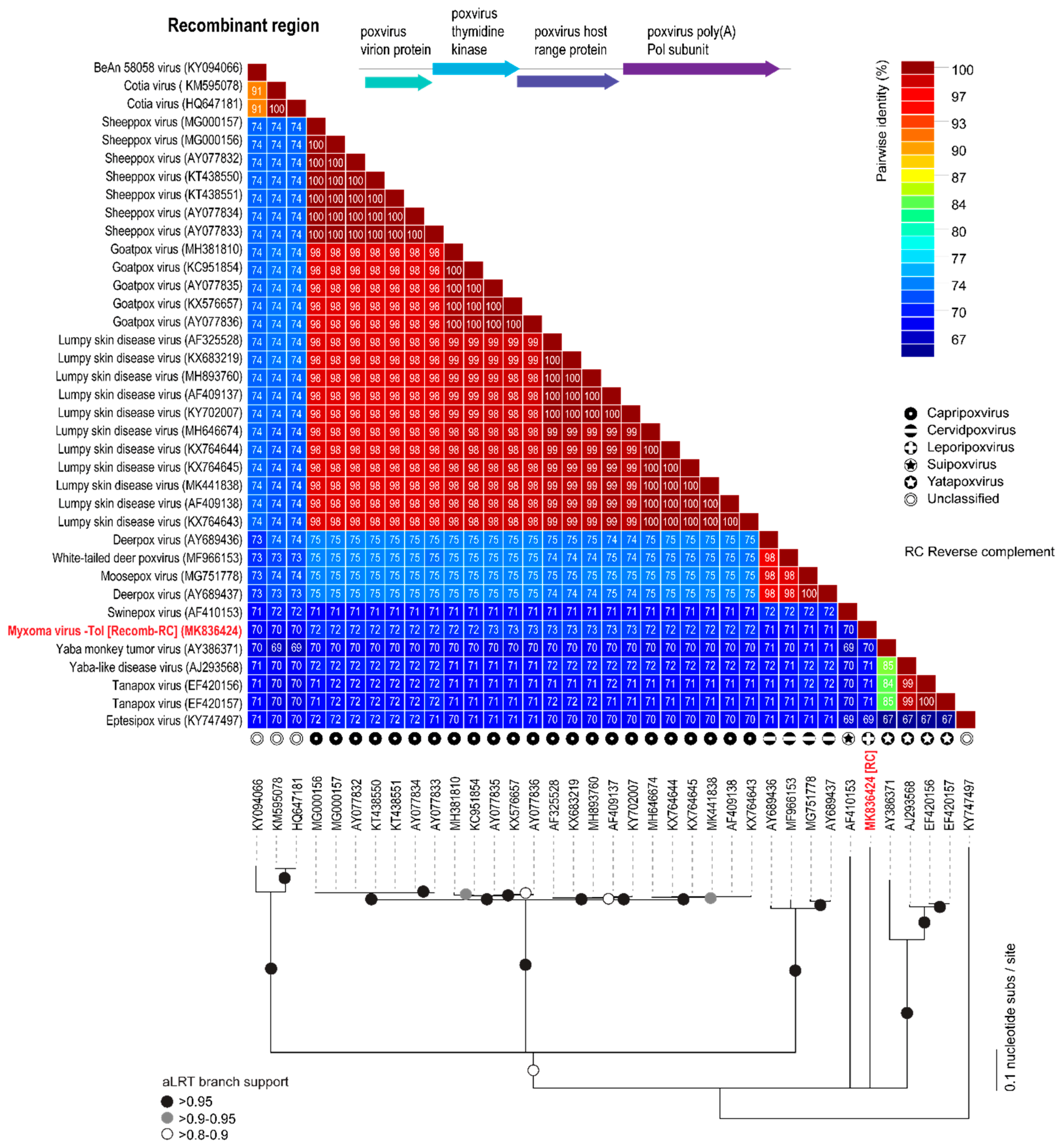
3.3. Analyses of the New Recombinant Region of the MYXV-Tol Isolate
4. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fenner, F.; Ratcliffe, F.N. Myxomatosis; Cambridge University Press: Cambridge, UK, 1965. [Google Scholar]
- Kerr, P.; Liu, J.; Cattadori, I.; Ghedin, E.; Read, A.; Holmes, E. Myxoma virus and the leporipoxviruses: An evolutionary paradigm. Viruses 2015, 7, 1020–1061. [Google Scholar] [CrossRef] [PubMed]
- Stanford, M.M.; Werden, S.J.; McFadden, G. Myxoma virus in the european rabbit: Interactions between the virus and its susceptible host. Veterin. Res. 2007, 38, 299–318. [Google Scholar] [CrossRef] [PubMed]
- Kerr, P.J. Myxomatosis in australia and europe: A model for emerging infectious diseases. Antivir. Res. 2012, 93, 387–415. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, L. Biological control of vertebrate pests: The history of myxomatosis, an experiment in evolution. Med. Hist. 2001, 45, 139–140. [Google Scholar] [CrossRef]
- Ross, J.; Sanders, M.F. The development of genetic resistance to myxomatosis in wild rabbits in britain. J. Hyg. 1984, 92, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Marshall, I.D.; Fenner, F. Studies in the epidemiology of infectious myxomatosis of rabbits. V. Changes in the innate resistance of australian wild rabbits exposed to myxomatosis. J. Hyg. 1958, 56, 288–302. [Google Scholar] [CrossRef] [PubMed]
- Alves, J.M.; Carneiro, M.; Cheng, J.Y.; Lemos de Matos, A.; Rahman, M.M.; Loog, L.; Campos, P.F.; Wales, N.; Eriksson, A.; Manica, A.; et al. Parallel adaptation of rabbit populations to myxoma virus. Science 2019, 363, 1319–1326. [Google Scholar] [CrossRef]
- Regnery, D.C.; Marshall, I.D. Studies in the epidemiology of myxomatosis in california: Iv. The susceptibility of six leporid species to californian myxoma virus and the relative infectivity of their tumors for mosquitoes. Am. J. Epidemiol. 1971, 94, 508–513. [Google Scholar] [CrossRef]
- Regnery, D.C. The epidemic potential of brazilian myxoma virus (lausanne strain) for three species of north american cottontails. Am. J. Epidemiol. 1971, 94, 514–519. [Google Scholar] [CrossRef]
- Collins, J. Myxomatosis in the common hare. Irish Veterin. J. 1955, 9, 268. [Google Scholar]
- Barlow, A.; Lawrence, K.; Everest, D.; Dastjerdi, A.; Finnegan, C.; Steinbach, F. Confirmation of myxomatosis in a european brown hare in great britain. Myxomatosis 2014, 175, 75–76. [Google Scholar] [CrossRef] [PubMed]
- Smallwood, S.E.; Rahman, M.M.; Smith, D.W.; McFadden, G. Myxoma virus: Propagation, purification, quantification, and storage. Curr. Protoc. Microbiol. 2010, 17, 14A.11.11–14A.11.20. [Google Scholar] [CrossRef] [PubMed]
- Di Pietro, F.; Ortenzi, F.; Tilio, M.; Concetti, F.; Napolioni, V. Genomic DNA extraction from whole blood stored from 15- to 30-years at −20 c by rapid phenol–chloroform protocol: A useful tool for genetic epidemiology studies. Mol. Cell. Probes 2011, 25, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. Spades: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Bushnell, B. Bbmap Short-Read Aligner, and Other Bioinformatics Tools. 2015. Available online: https://sourceforge.net/projects/bbmap/ (accessed on 18 August 2018).
- Nakamura, T.; Tomii, K.; Yamada, K.D.; Katoh, K. Parallelization of mafft for large-scale multiple sequence alignments. Bioinformatics 2018, 34, 2490–2492. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Muhire, B.M.; Varsani, A.; Martin, D.P. Sdt: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. A simple method to control over-alignment in the mafft multiple sequence alignment program. Bioinformatics 2016, 32, 1933–1942. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of phyml 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. Prottest 3: Fast selection of best-fit models of protein evolution. Bioinformatics 2011, 27, 1164–1165. [Google Scholar] [CrossRef]
- Stöver, B.C.; Müller, K.F. Treegraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 7. [Google Scholar] [CrossRef] [PubMed]
- Kerr, P.J.; Cattadori, I.M.; Liu, J.; Sim, D.G.; Dodds, J.W.; Brooks, J.W.; Kennett, M.J.; Holmes, E.C.; Read, A.F. Next step in the ongoing arms race between myxoma virus and wild rabbits in australia is a novel disease phenotype. Proc. Natl. Acad. Sci. USA 2017, 114, 9397–9402. [Google Scholar] [CrossRef] [PubMed]
- Cameron, C.; Hota-Mitchell, S.; Chen, L.; Barrett, J.; Cao, J.-X.; Macaulay, C.; Willer, D.; Evans, D.; McFadden, G. The complete DNA sequence of myxoma virus. Virology 1999, 264, 298–318. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.W.; McFadden, G. Genus leporipoxvirus. In Poxviruses; Mercer, A.A., Schmidt, A., Weber, O., Eds.; Birkhäuser Basel: Basel, Switzerland, 2007; pp. 183–201. [Google Scholar]
- Kerr, P.J.; Rogers, M.B.; Fitch, A.; DePasse, J.V.; Cattadori, I.M.; Twaddle, A.C.; Hudson, P.J.; Tscharke, D.C.; Read, A.F.; Holmes, E.C.; et al. Genome scale evolution of myxoma virus reveals host-pathogen adaptation and rapid geographic spread. J. Virol. 2013, 87, 12900–12915. [Google Scholar] [CrossRef] [PubMed]
- Kerr, P.J.; Ghedin, E.; DePasse, J.V.; Fitch, A.; Cattadori, I.M.; Hudson, P.J.; Tscharke, D.C.; Read, A.F.; Holmes, E.C. Evolutionary history and attenuation of myxoma virus on two continents. PLoS Pathog. 2012, 8, e1002950. [Google Scholar] [CrossRef] [PubMed]
- Kerr, P.J.; Cattadori, I.M.; Rogers, M.B.; Fitch, A.; Geber, A.; Liu, J.; Sim, D.G.; Boag, B.; Eden, J.-S.; Ghedin, E.; et al. Genomic and phenotypic characterization of myxoma virus from great britain reveals multiple evolutionary pathways distinct from those in australia. PLoS Pathog. 2017, 13, e1006252. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Rothenburg, S.; McFadden, G. The poxvirus c7l host range factor superfamily. Curr. Opin. Virol. 2012, 2, 764–772. [Google Scholar] [CrossRef]
- Saint, K.; French, N.; Kerr, P. Genetic variation in australian isolates of myxoma virus: An evolutionary and epidemiological study. Arch. Virol. 2001, 146, 1105–1123. [Google Scholar] [CrossRef]
- Guerin, J.-L.; Gelfi, J.; Camus, C.; Delverdier, M.; Whisstock, J.C.; Amardeihl, M.-F.; Py, R.; Bertagnoli, S.; Messud-Petit, F. Characterization and functional analysis of serp3: A novel myxoma virus-encoded serpin involved in virulence. J. Gen. Virol. 2001, 82, 1407–1417. [Google Scholar] [CrossRef]
- Upton, C.; Macen, J.; Wishart, D.; McFadden, G. Myxoma virus and malignant rabbit fibroma virus encode a serpin-like protein important for virus virulence. Virology 1990, 179, 618–631. [Google Scholar] [CrossRef]
- Viswanathan, K.; Liu, L.; Vaziri, S.; Dai, E.; Richardson, J.; Togonu-Bickersteth, B.; Vatsya, P.; Christov, A.; Lucas, A.R. Myxoma viral serpin, serp-1, a unique interceptor of coagulation and innate immune pathways. Thromb. Haemost. 2006, 95, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Macen, J.L.; Upton, C.; Nation, N.; McFadden, G. Serp1, a serine proteinase inhibitor encoded by myxoma virus, is a secreted glycoprotein that interferes with inflammation. Virology 1993, 195, 348–363. [Google Scholar] [CrossRef] [PubMed]
- Messud-Petit, F.; Gelfi, J.; Delverdier, M.; Amardeilh, M.-F.; Py, R.; Sutter, G.; Bertagnoli, S. Serp2, an inhibitor of the interleukin-1β-converting enzyme, is critical in the pathobiology of myxoma virus. J. Virol. 1998, 72, 7830–7839. [Google Scholar] [PubMed]
- Kerr, P.J.; Rogers, M.B.; Fitch, A.; Depasse, J.V.; Cattadori, I.M.; Hudson, P.J.; Tscharke, D.C.; Holmes, E.C.; Ghedin, E. Comparative analysis of the complete genome sequence of the california msw strain of myxoma virus reveals potential host adaptations. J. Virol. 2013, 87, 12080–12089. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Haller, S.L.; Rahman, M.M.; McFadden, G.; Rothenburg, S. Myxoma virus m156 is a specific inhibitor of rabbit pkr but contains a loss-of-function mutation in australian virus isolates. Proc. Natl. Acad. Sci. USA 2016, 113, 3855–3860. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wennier, S.; Zhang, L.; McFadden, G. M062 is a host range factor essential for myxoma virus pathogenesis and functions as an antagonist of host samd9 in human cells. J. Virol. 2011, 85, 3270–3282. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wennier, S.; Moussatche, N.; Reinhard, M.; Condit, R.; McFadden, G. Myxoma virus m064 is a novel member of the poxvirus c7l superfamily of host range factors that controls the kinetics of myxomatosis in european rabbits. J. Virol. 2012, 86, 5371–5375. [Google Scholar] [CrossRef]
- Barrett, J.W.; Shun Chang, C.; Wang, G.; Werden, S.J.; Shao, Z.; Barrett, C.; Gao, X.; Belsito, T.A.; Villenevue, D.; McFadden, G. Myxoma virus m063r is a host range gene essential for virus replication in rabbit cells. Virology 2007, 361, 123–132. [Google Scholar] [CrossRef]
- Werden, S.J.; Rahman, M.M.; McFadden, G. Poxvirus host range genes. In Advances in Virus Research; Academic Press: Cambridge, MA, USA, 2008; Volume 71, pp. 135–171. [Google Scholar]
- Oliveira, G.; Rodrigues, R.; Lima, M.; Drumond, B.; Abrahão, J. Poxvirus host range genes and virus–host spectrum: A critical review. Viruses 2017, 9, 331. [Google Scholar] [CrossRef]
- Merchant, J.C.; Kerr, P.J.; Simms, N.G.; Robinson, A.J. Monitoring the spread of myxoma virus in rabbit oryctolagus cuniculus populations on the southern tablelands of new south wales, australia. I. Natural occurrence of myxomatosis. Epidemiol. Infect. 2003, 130, 113–121. [Google Scholar] [CrossRef]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Águeda-Pinto, A.; Lemos de Matos, A.; Abrantes, M.; Kraberger, S.; Risalde, M.A.; Gortázar, C.; McFadden, G.; Varsani, A.; Esteves, P.J. Genetic Characterization of a Recombinant Myxoma Virus in the Iberian Hare (Lepus granatensis). Viruses 2019, 11, 530. https://doi.org/10.3390/v11060530
Águeda-Pinto A, Lemos de Matos A, Abrantes M, Kraberger S, Risalde MA, Gortázar C, McFadden G, Varsani A, Esteves PJ. Genetic Characterization of a Recombinant Myxoma Virus in the Iberian Hare (Lepus granatensis). Viruses. 2019; 11(6):530. https://doi.org/10.3390/v11060530
Chicago/Turabian StyleÁgueda-Pinto, Ana, Ana Lemos de Matos, Mário Abrantes, Simona Kraberger, Maria A. Risalde, Christian Gortázar, Grant McFadden, Arvind Varsani, and Pedro J. Esteves. 2019. "Genetic Characterization of a Recombinant Myxoma Virus in the Iberian Hare (Lepus granatensis)" Viruses 11, no. 6: 530. https://doi.org/10.3390/v11060530
APA StyleÁgueda-Pinto, A., Lemos de Matos, A., Abrantes, M., Kraberger, S., Risalde, M. A., Gortázar, C., McFadden, G., Varsani, A., & Esteves, P. J. (2019). Genetic Characterization of a Recombinant Myxoma Virus in the Iberian Hare (Lepus granatensis). Viruses, 11(6), 530. https://doi.org/10.3390/v11060530