A Field Recombinant Strain Derived from Two Type 1 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV-1) Modified Live Vaccines Shows Increased Viremia and Transmission in SPF Pigs

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Viruses
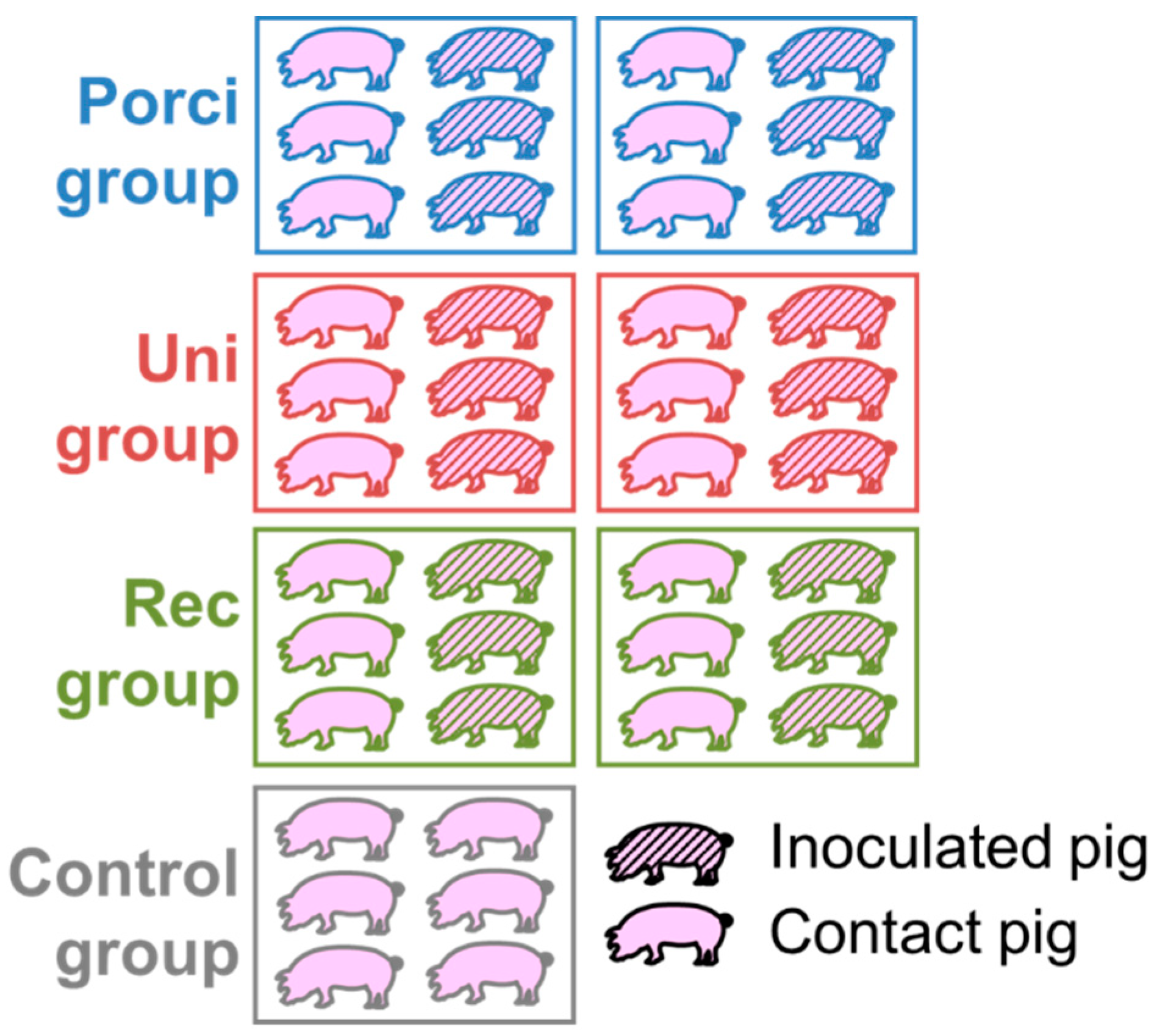
2.2. Animal Experiment
2.3. Quantification of PRRS Viral Genome Load in Sera, Nasal Swabs, and Tissue Samples
2.4. On Farm Follow-Up of the Recombinant Strain
2.5. Statistical Analyses and Transmission Parameters
3. Results
3.1. Clinical Data
3.2. Virological Parameters in Inoculated Pigs
3.2.1. Viremia
3.2.2. Nasal Viral Shedding
3.2.3. Viral Load in Tonsils
3.3. Virological Parameters in Contact Pigs
3.4. Transmission Parameters
3.5. Genomic Evolution of the Recombinant Strain Persisting on the Farm
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Albina, E. Porcine reproductive and respiratory syndrome: Ten years of experience (1986–1996) with this undesirable viral infection. Vet. Res. 1997, 28, 305–352. [Google Scholar] [PubMed]
- Nathues, H.; Alarcon, P.; Rushton, J.; Jolie, R.; Fiebig, K.; Jimenez, M.; Geurts, V.; Nathues, C. Cost of porcine reproductive and respiratory syndrome virus at individual farm level—An economic disease model. Prev. Vet. Med. 2017, 142, 16–29. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.J. Emerging and re-emerging swine viruses. Transbound. Emerg. Dis. 2012, 59 (Suppl. 1), 85–102. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Yang, H. Porcine reproductive and respiratory syndrome in China. Virus Res. 2010, 154, 31–37. [Google Scholar] [CrossRef]
- Balasuriya, U.B.; Carossino, M. Reproductive effects of arteriviruses: Equine arteritis virus and porcine reproductive and respiratory syndrome virus infections. Curr. Opin. Virol. 2017, 27, 57–70. [Google Scholar] [CrossRef]
- Guo, Z.; Chen, X.X.; Li, R.; Qiao, S.; Zhang, G. The prevalent status and genetic diversity of porcine reproductive and respiratory syndrome virus in China: A molecular epidemiological perspective. Virol. J. 2018, 15, 2. [Google Scholar] [CrossRef]
- Allende, R.; Lewis, T.L.; Lu, Z.; Rock, D.L.; Kutish, G.F.; Ali, A.; Doster, A.R.; Osorio, F.A. North American and European porcine reproductive and respiratory syndrome viruses differ in non-structural protein coding regions. J. Gen. Virol. 1999, 80 Pt 2, 307–315. [Google Scholar] [CrossRef]
- Martinez-Lobo, F.J.; Diez-Fuertes, F.; Segales, J.; Garcia-Artiga, C.; Simarro, I.; Castro, J.M.; Prieto, C. Comparative pathogenicity of type 1 and type 2 isolates of porcine reproductive and respiratory syndrome virus (PRRSV) in a young pig infection model. Vet. Microbiol. 2011, 154, 58–68. [Google Scholar] [CrossRef]
- Nan, Y.; Wu, C.; Gu, G.; Sun, W.; Zhang, Y.J.; Zhou, E.M. Improved Vaccine against PRRSV: Current Progress and Future Perspective. Front. Microbiol. 2017, 8, 1635. [Google Scholar] [CrossRef]
- Charerntantanakul, W. Porcine reproductive and respiratory syndrome virus vaccines: Immunogenicity, efficacy and safety aspects. World J. Virol. 2012, 1, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Kappes, M.A.; Faaberg, K.S. PRRSV structure, replication and recombination: Origin of phenotype and genotype diversity. Virology 2015, 479–480, 475–486. [Google Scholar] [CrossRef]
- Murtaugh, M.P.; Stadejek, T.; Abrahante, J.E.; Lam, T.T.; Leung, F.C. The ever-expanding diversity of porcine reproductive and respiratory syndrome virus. Virus Res. 2010, 154, 18–30. [Google Scholar] [CrossRef]
- Yun, S.I.; Lee, Y.M. Overview: Replication of porcine reproductive and respiratory syndrome virus. J. Microbiol. 2013, 51, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Simon-Loriere, E.; Holmes, E.C. Why do RNA viruses recombine? Nat. Rev. Microbiol. 2011, 9, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Snijder, E.J.; Kikkert, M.; Fang, Y. Arterivirus molecular biology and pathogenesis. J. Gen. Virol. 2013, 94 Pt 10, 2141–2163. [Google Scholar] [CrossRef]
- Wang, H.M.; Liu, Y.G.; Tang, Y.D.; Liu, T.X.; Zheng, L.L.; Wang, T.Y.; Liu, S.G.; Wang, G.; Cai, X.H. A natural recombinant PRRSV between HP-PRRSV JXA1-like and NADC30-like strains. Transbound. Emerg. Dis. 2018, 65, 1078–1086. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhou, X.; Zhai, J.; Wei, C.; Dai, A.; Yang, X.; Luo, M. Recombination in JXA1-R vaccine and NADC30-like strain of porcine reproductive and respiratory syndrome viruses. Vet. Microbiol. 2017, 204, 110–120. [Google Scholar] [CrossRef]
- Chen, N.; Yu, X.; Wang, L.; Wu, J.; Zhou, Z.; Ni, J.; Li, X.; Zhai, X.; Tian, K. Two natural recombinant highly pathogenic porcine reproductive and respiratory syndrome viruses with different pathogenicities. Virus Genes 2013, 46, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Wang, Y.; Yu, L.; Zhang, P.; Liu, X.; Zhang, L.; Liu, Y.; Liang, P.; Wang, L.; Song, C. Pathogenicity of a newly emerged recombined porcine reproductive and respiratory syndrome virus strain (subgenotype III) in China. Vet. Microbiol. 2017, 210, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Fang, L.; Xu, Z.; Liu, S.; Gao, J.; Jiang, Y.; Chen, H.; Xiao, S. Recombination in vaccine and circulating strains of porcine reproductive and respiratory syndrome viruses. Emerg. Infect. Dis. 2009, 15, 2032–2035. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Wei, Z.; Zhang, G.; Li, Z.; Ma, J.; Xie, Q.; Sun, B.; Bi, Y. Complete genome sequence of a novel variant porcine reproductive and respiratory syndrome virus (PRRSV) strain: Evidence for recombination between vaccine and wild-type PRRSV strains. J. Virol. 2012, 86, 9543. [Google Scholar]
- Zhao, H.; Han, Q.; Zhang, L.; Zhang, Z.; Wu, Y.; Shen, H.; Jiang, P. Emergence of mosaic recombinant strains potentially associated with vaccine JXA1-R and predominant circulating strains of porcine reproductive and respiratory syndrome virus in different provinces of China. Virol. J. 2017, 14, 67. [Google Scholar] [CrossRef] [PubMed]
- Madsen, K.G.; Hansen, C.M.; Madsen, E.S.; Strandbygaard, B.; Botner, A.; Sorensen, K.J. Sequence analysis of porcine reproductive and respiratory syndrome virus of the American type collected from Danish swine herds. Arch. Virol. 1998, 143, 1683–1700. [Google Scholar] [CrossRef] [PubMed]
- Bian, T.; Sun, Y.; Hao, M.; Zhou, L.; Ge, X.; Guo, X.; Han, J.; Yang, H. A recombinant type 2 porcine reproductive and respiratory syndrome virus between NADC30-like and a MLV-like: Genetic characterization and pathogenicity for piglets. Infect. Genet. Evol. 2017, 54, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.H.; Tun, H.M.; Sun, B.L.; Mo, J.; Zhou, Q.F.; Deng, Y.X.; Xie, Q.M.; Bi, Y.Z.; Leung, F.C.; Ma, J.Y. Re-emerging of porcine respiratory and reproductive syndrome virus (lineage 3) and increased pathogenicity after genomic recombination with vaccine variant. Vet. Microbiol. 2015, 175, 332–340. [Google Scholar] [CrossRef]
- Frossard, J.P.; Hughes, G.J.; Westcott, D.G.; Naidu, B.; Williamson, S.; Woodger, N.G.; Steinbach, F.; Drew, T.W. Porcine reproductive and respiratory syndrome virus: Genetic diversity of recent British isolates. Vet. Microbiol. 2013, 162, 507–518. [Google Scholar] [CrossRef]
- Chen, N.; Liu, Q.; Qiao, M.; Deng, X.; Chen, X.; Sun, M. Whole genome characterization of a novel porcine reproductive and respiratory syndrome virus 1 isolate: Genetic evidence for recombination between Amervac vaccine and circulating strains in mainland China. Infect. Genet. Evol. 2017, 54, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Renson, P.; Touzain, F.; Lebret, A.; Le Dimna, M.; Quenault, H.; Normand, V.; Claude, J.B.; Pez, F.; Rose, N.; Blanchard, Y.; et al. Complete Genome Sequence of a Recombinant Porcine Reproductive and Respiratory Syndrome Virus Strain from Two Genotype 1 Modified Live Virus Vaccine Strains. Genome Announc. 2017, 5, e00454-17. [Google Scholar] [CrossRef] [PubMed]
- OIE. Porcine Reproductive and Respiratory Syndrome. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; OIE: Paris, France, 2015. [Google Scholar]
- Charpin, C.; Mahe, S.; Keranflec’h, A.; Belloc, C.; Cariolet, R.; Le Potier, M.F.; Rose, N. Infectiousness of pigs infected by the Porcine Reproductive and Respiratory Syndrome virus (PRRSV) is time-dependent. Vet. Res. 2012, 43, 69. [Google Scholar] [CrossRef]
- Rose, N.; Renson, P.; Andraud, M.; Paboeuf, F.; Le Potier, M.F.; Bourry, O. Porcine reproductive and respiratory syndrome virus (PRRSv) modified-live vaccine reduces virus transmission in experimental conditions. Vaccine 2015, 33, 2493–2499. [Google Scholar] [CrossRef]
- Loving, C.L.; Brockmeier, S.L.; Vincent, A.L.; Lager, K.M.; Sacco, R.E. Differences in clinical disease and immune response of pigs challenged with a high-dose versus low-dose inoculum of porcine reproductive and respiratory syndrome virus. Viral Immunol. 2008, 21, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Haiwick, G.; Hermann, J.; Roof, M.; Fergen, B.; Philips, R.; Patterson, A. Examination of viraemia and clinical signs after challenge with a heterologous PRRSV strain in PRRS Type 2 MLV vaccinated pigs: A challenge-dose study. PLoS ONE 2018, 13, e0209784. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Lobo, F.J.; de Lome, L.C.; Diez-Fuertes, F.; Segales, J.; Garcia-Artiga, C.; Simarro, I.; Castro, J.M.; Prieto, C. Safety of Porcine Reproductive and Respiratory Syndrome Modified Live Virus (MLV) vaccine strains in a young pig infection model. Vet. Res. 2013, 44, 115. [Google Scholar] [CrossRef] [PubMed]
- Scortti, M.; Prieto, C.; Martinez-Lobo, F.J.; Simarro, I.; Castro, J.M. Effects of two commercial European modified-live vaccines against porcine reproductive and respiratory syndrome viruses in pregnant gilts. Vet. J. 2006, 172, 506–514. [Google Scholar] [CrossRef]
- Chang, C.C.; Yoon, K.J.; Zimmerman, J.J.; Harmon, K.M.; Dixon, P.M.; Dvorak, C.M.; Murtaugh, M.P. Evolution of porcine reproductive and respiratory syndrome virus during sequential passages in pigs. J. Virol. 2002, 76, 4750–4763. [Google Scholar] [CrossRef] [PubMed]
- Murtaugh, M.P. Use and interpretation of sequencing in PRRSv control programs. In Proceedings of the Veterinary Continuing Education, Allen D. Leman Swine Conference, St. Paul, MN, USA, 15–18 September 2012; p. 39. [Google Scholar]
- Botner, A.; Nielsen, J.; Oleksiewicz, M.B.; Storgaard, T. Heterologous challenge with porcine reproductive and respiratory syndrome (PRRS) vaccine virus: No evidence of reactivation of previous European-type PRRS virus infection. Vet. Microbiol. 1999, 68, 187–195. [Google Scholar] [CrossRef]
- Mengeling, W.L.; Vorwald, A.C.; Lager, K.M.; Clouser, D.F.; Wesley, R.D. Identification and clinical assessment of suspected vaccine-related field strains of porcine reproductive and respiratory syndrome virus. Am. J. Vet. Res. 1999, 60, 334–340. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TRANSMISSION PARAMETERS | Porci | Uni | Rec |
|---|---|---|---|
| Duration of viral shedding (days) | 23.3 | 30.1 | 34.7 |
| (20.6; 26.3) * | (26.2; 34.7) | (30.6; 39.3) | |
| Duration of latency (days) | 0.5 | 1.0 | 0.5 |
| (0.3; 0.9) | (0.6; 2.0) | (0.3; 1.0) | |
| Daily transmission rate (number of infected pigs by an infectious pig per day) | 0.11 | 0.08 | 0.57 |
| (0.05; 0.22) | (0.03; 0.16) | (0.23; 1.23) |
| PRRS-FR-2014-56-11-1 Recombinant Strain Isolated in 2014 | PRRS-FR-2016-56-11-1 Recombinant Strain Isolated in 2016 | ||
|---|---|---|---|
| Event No. 1 | Segment length and position | 500–1370 nt | 510–1408 nt |
| % similarity with Porcilis | 99.5% | 98.0% | |
| p value RDP | 3.372.10−34 | 9.727.10−25 | |
| Event No. 2 | Segment length and position | 3646–4272 nt | 3646–4272 nt |
| % similarity with Porcilis | 99.5% | 98.7% | |
| p value RDP | 5.790.10−17 | 9.431.10−14 | |
| Event No. 3 | Segment length and position | 4972–8430 nt | 4994–8376 nt |
| % similarity with Porcilis | 99.5% | 98.4% | |
| p value RDP | 3.894.10−63 | 6.115.10−47 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eclercy, J.; Renson, P.; Lebret, A.; Hirchaud, E.; Normand, V.; Andraud, M.; Paboeuf, F.; Blanchard, Y.; Rose, N.; Bourry, O. A Field Recombinant Strain Derived from Two Type 1 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV-1) Modified Live Vaccines Shows Increased Viremia and Transmission in SPF Pigs. Viruses 2019, 11, 296. https://doi.org/10.3390/v11030296
Eclercy J, Renson P, Lebret A, Hirchaud E, Normand V, Andraud M, Paboeuf F, Blanchard Y, Rose N, Bourry O. A Field Recombinant Strain Derived from Two Type 1 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV-1) Modified Live Vaccines Shows Increased Viremia and Transmission in SPF Pigs. Viruses. 2019; 11(3):296. https://doi.org/10.3390/v11030296
Chicago/Turabian StyleEclercy, Julie, Patricia Renson, Arnaud Lebret, Edouard Hirchaud, Valérie Normand, Mathieu Andraud, Frédéric Paboeuf, Yannick Blanchard, Nicolas Rose, and Olivier Bourry. 2019. "A Field Recombinant Strain Derived from Two Type 1 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV-1) Modified Live Vaccines Shows Increased Viremia and Transmission in SPF Pigs" Viruses 11, no. 3: 296. https://doi.org/10.3390/v11030296
APA StyleEclercy, J., Renson, P., Lebret, A., Hirchaud, E., Normand, V., Andraud, M., Paboeuf, F., Blanchard, Y., Rose, N., & Bourry, O. (2019). A Field Recombinant Strain Derived from Two Type 1 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV-1) Modified Live Vaccines Shows Increased Viremia and Transmission in SPF Pigs. Viruses, 11(3), 296. https://doi.org/10.3390/v11030296