Mastomys Species as Model Systems for Infectious Diseases


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
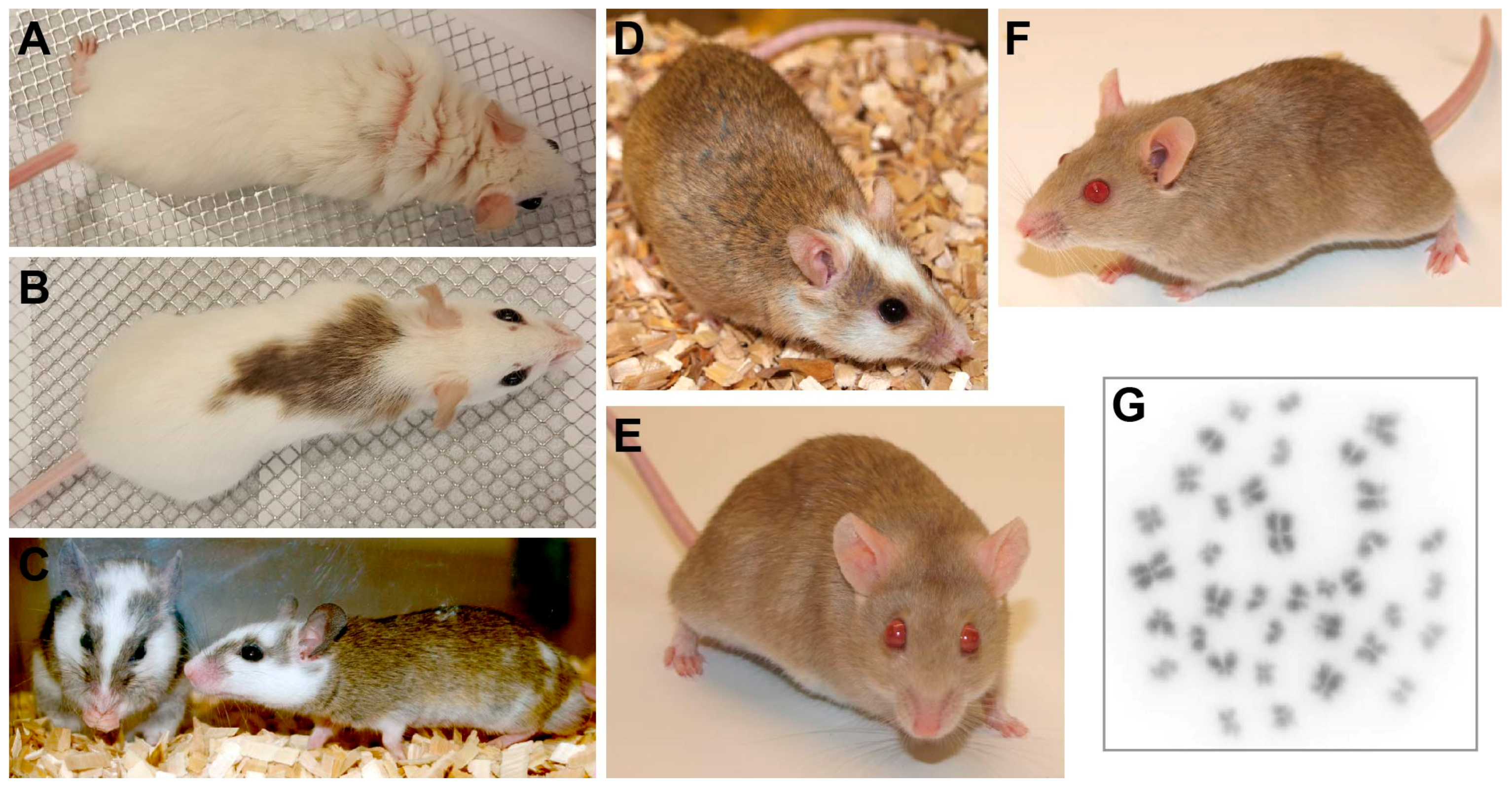
2. Characteristics of Mastomys Species
3. Housing of Mastomys in the Laboratory
4. Mastomys as Model Systems in Biomedical Research—Historical Flashback
5. Mastomys coucha as a Preclinical Model in Papillomavirus Research
5.1. History of Mastomys coucha in Papillomavirus Research
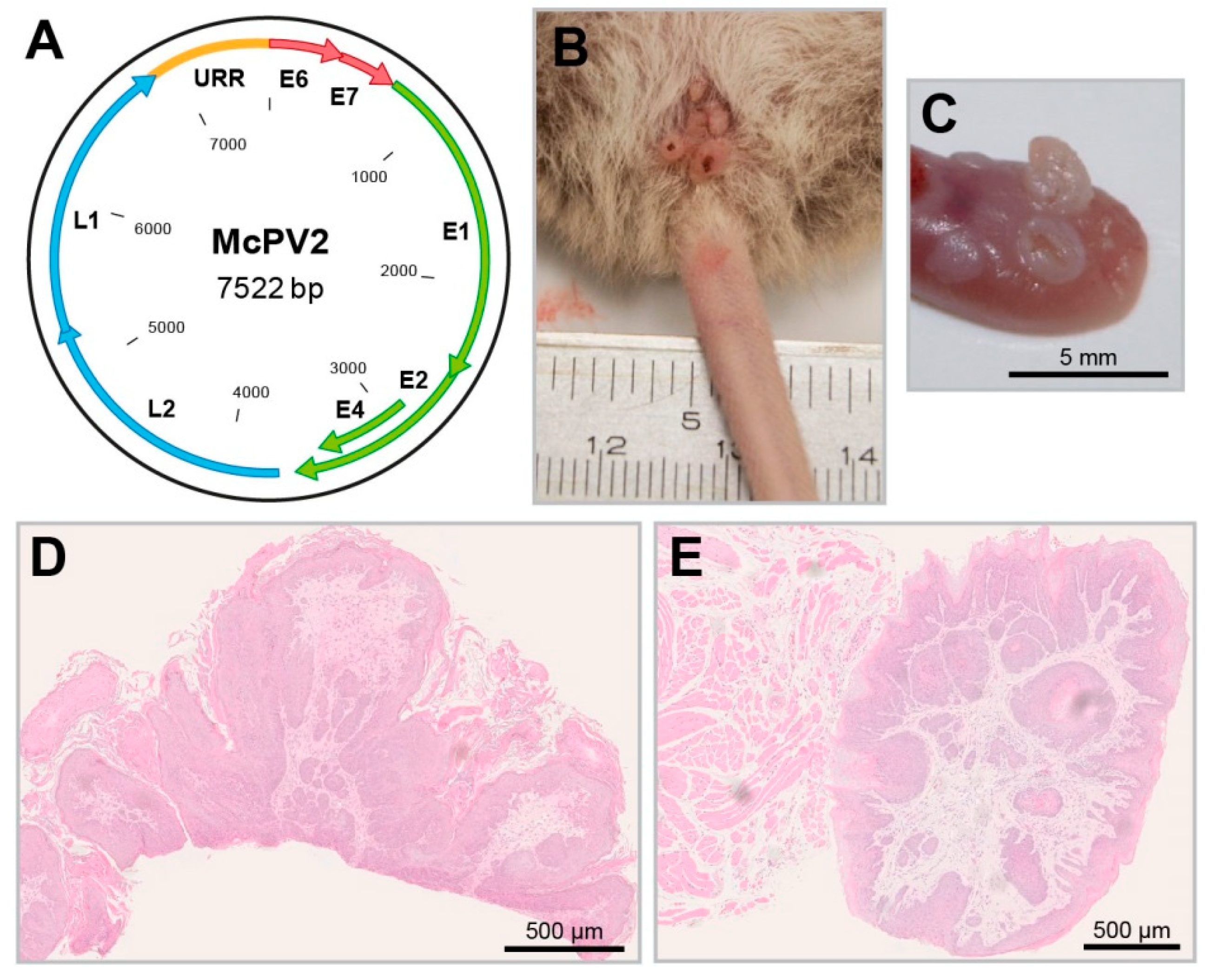
5.2. Recent Contributions of Mastomys coucha to PV Research as a Preclinical Model
6. Vaccination and Tumor Prevention in Mastomys coucha
7. The Role of UV Exposure and Papillomavirus Infection in NMSC Development
8. Summary
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rheinberger, H.J. Preparations, models, and simulations. Hist. Philos. Life Sci. 2015, 36, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Wright, A. Milestone 3: (1925) the Spinning Electron. Available online: https://www.nature.com/milestones/milespin/pdf/milespin03.pdf (accessed on 22 January 2019).
- Van Regenmortel, M.H. Reductionism and complexity in molecular biology. Scientists now have the tools to unravel biological and overcome the limitations of reductionism. EMBO Rep. 2004, 5, 1016–1020. [Google Scholar] [PubMed]
- Sverdlov, E.D. Unsolvable problems of biology: It is impossible to create two identical organisms, to defeat cancer, or to map organisms onto their genomes. Biochemistry 2018, 83, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Massoud, T.F.; Hademenos, G.J.; Young, W.L.; Gao, E.; Pile-Spellman, J.; Vinuela, F. Principles and philosophy of modeling in biomedical research. FASEB J. 1998, 12, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Shamir, E.R.; Ewald, A.J. Three-dimensional organotypic culture: Experimental models of mammalian biology and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 647–664. [Google Scholar] [CrossRef] [PubMed]
- Horvath, P.; Aulner, N.; Bickle, M.; Davies, A.M.; Nery, E.D.; Ebner, D.; Montoya, M.C.; Ostling, P.; Pietiainen, V.; Price, L.S.; et al. Screening out irrelevant cell-based models of disease. Nat. Rev. Drug Discov. 2016, 15, 751–769. [Google Scholar] [CrossRef] [PubMed]
- Day, C.P.; Merlino, G.; van Dyke, T. Preclinical mouse cancer models: A maze of opportunities and challenges. Cell 2015, 163, 39–53. [Google Scholar] [CrossRef]
- Dolgos, H.; Trusheim, M.; Gross, D.; Halle, J.P.; Ogden, J.; Osterwalder, B.; Sedman, E.; Rossetti, L. Translational medicine guide transforms drug development processes: The recent merck experience. Drug Discov. Today 2016, 21, 517–526. [Google Scholar] [CrossRef]
- Delaney, A.; Angus, D.C.; Bellomo, R.; Cameron, P.; Cooper, D.J.; Finfer, S.; Harrison, D.A.; Huang, D.T.; Myburgh, J.A.; Peake, S.L.; et al. Bench-to-bedside review: The evaluation of complex interventions in critical care. Crit. Care 2008, 12, 210. [Google Scholar] [CrossRef]
- Hobin, J.A.; Galbraith, R.A. Engaging basic scientists in translational research. FASEB J. 2012, 26, 2227–2230. [Google Scholar] [CrossRef]
- Lazebnik, Y. Are scientists a workforce?—Or, how Dr. Frankenstein made biomedical research sick: A proposed plan to rescue us biomedical research from its current “malaise” will not be effective as it misdiagnoses the root cause of the disease. EMBO Rep. 2015, 16, 1592–1600. [Google Scholar] [CrossRef] [PubMed]
- Bentley, P.J.; Gulbrandsen, M.; Kyvik, S. The relationship between basic and applied research in universities. High Educ. 2015, 70, 689–709. [Google Scholar] [CrossRef]
- Reeves, M.Q.; Kandyba, E.; Harris, S.; Del Rosario, R.; Balmain, A. Multicolour lineage tracing reveals clonal dynamics of squamous carcinoma evolution from initiation to metastasis. Nat. Cell Biol. 2018, 20, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Dougherty, E.R.; Shmulevich, I. On the limitations of biological knowledge. Curr. Genom. 2012, 13, 574–587. [Google Scholar] [CrossRef]
- Anisimov, V.N.; Ukraintseva, S.V.; Yashin, A.I. Cancer in rodents: Does it tell us about cancer in humans? Nat. Rev. Cancer 2005, 5, 807–819. [Google Scholar] [CrossRef]
- Moher, D.; Glasziou, P.; Chalmers, I.; Nasser, M.; Bossuyt, P.M.; Korevaar, D.A.; Graham, I.D.; Ravaud, P.; Boutron, I. Increasing value and reducing waste in biomedical research: Who’s listening? Lancet 2016, 387, 1573–1586. [Google Scholar] [CrossRef]
- Chalmers, I.; Bracken, M.B.; Djulbegovic, B.; Garattini, S.; Grant, J.; Gulmezoglu, A.M.; Howells, D.W.; Ioannidis, J.P.; Oliver, S. How to increase value and reduce waste when research priorities are set. Lancet 2014, 383, 156–165. [Google Scholar] [CrossRef]
- Schneierson, S.S.; Perlman, E. Toxicity of penicillin for the syrian hamster. Proc. Soc. Exp. Biol. Med. 1956, 91, 229–230. [Google Scholar] [CrossRef] [PubMed]
- Green, R.H. The association of viral activation with penicillin toxicity in guinea pigs and hamsters. Yale J. Biol. Med. 1974, 47, 166–181. [Google Scholar]
- Rand, M.S. Selection of biomedical animal models. In Sourcebook of Models for Biomedical Research; Conn, P.M., Ed.; Humana Press Inc.: Totowa, NJ, USA, 2008; pp. 9–16. [Google Scholar]
- Sarasamma, S.; Lai, Y.H.; Liang, S.T.; Liu, K.; Hsiao, C.D. The power of fish models to elucidate skin cancer pathogenesis and impact the discovery of new therapeutic opportunities. Int. J. Mol. Sci. 2018, 19, 3929. [Google Scholar] [CrossRef]
- Zhao, S.; Huang, J.; Ye, J. A fresh look at zebrafish from the perspective of cancer research. J. Exp. Clin. Cancer Res. 2015, 34, 80. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.; Orme, I.M. Animal models of tuberculosis: An overview. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Christensen, N.D.; Budgeon, L.R.; Cladel, N.M.; Hu, J. Recent advances in preclinical model systems for papillomaviruses. Virus Res. 2017, 231, 108–118. [Google Scholar] [CrossRef] [PubMed]
- McGovern, J.A.; Griffin, M.; Hutmacher, D.W. Animal models for bone tissue engineering and modelling disease. Dis. Model. Mech. 2018, 11, dmm033084. [Google Scholar] [CrossRef]
- Workman, P.; Aboagye, E.O.; Balkwill, F.; Balmain, A.; Bruder, G.; Chaplin, D.J.; Double, J.A.; Everitt, J.; Farningham, D.A.; Glennie, M.J.; et al. Guidelines for the welfare and use of animals in cancer research. Br. J. Cancer 2010, 102, 1555–1577. [Google Scholar] [CrossRef] [PubMed]
- Denayer, T.; Stöhr, T.; van Roy, M. Animal models in translational medicine: Validation and prediction. New Horiz. Transl. Med. 2014, 2, 5–11. [Google Scholar] [CrossRef]
- Khaled, W.T.; Liu, P. Cancer mouse models: Past, present and future. Semin. Cell Dev. Biol. 2014, 27, 54–60. [Google Scholar] [CrossRef]
- Justice, M.J.; Dhillon, P. Using the mouse to model human disease: Increasing validity and reproducibility. Dis. Model. Mech. 2016, 9, 101–103. [Google Scholar] [CrossRef]
- Beck, J.A.; Lloyd, S.; Hafezparast, M.; Lennon-Pierce, M.; Eppig, J.T.; Festing, M.F.; Fisher, E.M. Genealogies of mouse inbred strains. Nat. Genet. 2000, 24, 23–25. [Google Scholar] [CrossRef]
- Hau, J. Animal models for human diseases. In Sourcebook of Models for Biomedical Research; Conn, P.M., Ed.; Humana Press Inc.: Totowa, NJ, USA, 2008; pp. 3–8. [Google Scholar]
- Pelleitier, M.; Montplaisir, S. The nude mouse: A model of deficient t-cell function. Methods Achiev. Exp. Pathol. 1975, 7, 149–166. [Google Scholar]
- Geissler, E.K. Post-transplantation malignancies: Here today, gone tomorrow? Nat. Rev. Clin. Oncol. 2015, 12, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Arroyo Muhr, L.S.; Bzhalava, Z.; Hortlund, M.; Lagheden, C.; Nordqvist Kleppe, S.; Bzhalava, D.; Hultin, E.; Dillner, J. Viruses in cancers among the immunosuppressed. Int. J. Cancer. 2017, 141, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Abolins, S.R.; Pocock, M.J.; Hafalla, J.C.; Riley, E.M.; Viney, M.E. Measures of immune function of wild mice, mus musculus. Mol. Ecol. 2011, 20, 881–892. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Wu, J.; Jin, D.; Wang, B.; Cao, H. Fecal microbiota transplantation in cancer management: Current status and perspectives. Int. J. Cancer. 2018. [Google Scholar] [CrossRef] [PubMed]
- Hamada, T.; Nowak, J.A.; Milner, D.A., Jr.; Song, M.; Ogino, S. Integration of microbiology, molecular pathology, and epidemiology: A new paradigm to explore the pathogenesis of microbiome-driven neoplasms. J. Pathol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.Z.; Ravindran, R.; Chassaing, B.; Carvalho, F.A.; Maddur, M.S.; Bower, M.; Hakimpour, P.; Gill, K.P.; Nakaya, H.I.; Yarovinsky, F.; et al. Tlr5-mediated sensing of gut microbiota is necessary for antibody responses to seasonal influenza vaccination. Immunity 2014, 41, 478–492. [Google Scholar] [CrossRef]
- Kennedy, E.A.; King, K.Y.; Baldridge, M.T. Mouse microbiota models: Comparing germ-free mice and antibiotics treatment as tools for modifying gut bacteria. Front. Physiol. 2018, 9, 1534. [Google Scholar] [CrossRef]
- Conti, F.; Abnave, P.; Ghigo, E. Unconventional animal models: A booster for new advances in host-pathogen interactions. Front. Cell Infect. Microbiol. 2014, 4, 142. [Google Scholar] [CrossRef]
- Gladfelter, A.S. How nontraditional model systems can save us. Mol. Biol. Cell 2015, 26, 3687–3689. [Google Scholar] [CrossRef]
- Gutekunst, J.; Andriantsoa, R.; Falckenhayn, C.; Hanna, K.; Stein, W.; Rasamy, J.; Lyko, F. Clonal genome evolution and rapid invasive spread of the marbled crayfish. Nat. Ecol. Evol. 2018, 2, 567–573. [Google Scholar] [CrossRef]
- Petruseva, I.O.; Evdokimov, A.N.; Lavrik, O.I. Genome stability maintenance in naked mole-rat. Acta Nat. 2017, 9, 31–41. [Google Scholar]
- Zhao, Y.; Tyshkovskiy, A.; Munoz-Espin, D.; Tian, X.; Serrano, M.; de Magalhaes, J.P.; Nevo, E.; Gladyshev, V.N.; Seluanov, A.; Gorbunova, V. Naked mole rats can undergo developmental, oncogene-induced and DNA damage-induced cellular senescence. Proc. Natl. Acad. Sci. USA 2018, 115, 1801–1806. [Google Scholar] [CrossRef]
- Jansa, S.A.; Giarla, T.C.; Lim, B.K. The phylogenetic position of the rodent genus typhlomys and the geographic origin of muroidea. J. Mammal. 2009, 90, 1083–1094. [Google Scholar] [CrossRef]
- Lecompte, E.; Granjon, L.; Denys, C. The phylogeny of the praomys complex (rodentia: Muridae) and its phylogeographic implications. J. Zool. Syst. Evol. Res. 2002, 40, 8–25. [Google Scholar] [CrossRef]
- Chevret, P.; Granjon, L.; Duplantier, J.M.; Denys, C.; Catzeflis, F.M. Molecular phylogeny of the praomys complex (rodentia, murinae)—A study based on DNA/DNA hybridization experiments. Zool. J. Linn. Soc.-Lond. 1994, 112, 425–442. [Google Scholar] [CrossRef]
- Granjon, L.; Duplantier, J.M.; Catalan, J.; BrittonDavidian, J. Systematics of the genus mastomys (thomas, 1915) (rodentia: Muridae)—A review. Belg. J. Zool. 1997, 127, 7–18. [Google Scholar]
- Solleveld, H.A. The multimammate mouse. In The Ufaw Handbook on the Care and Management of Laboratory Animals, Subsequent edition (1 June 1987) ed.; Churchill Livingstone Inc.: New York, NY, USA, 1987; p. 948. [Google Scholar]
- Weiss, J.; Becker, K.; Bernsmann, E.; Chourbaji, S.; Dietrich, H. Versuchstierkunde: Tierpflege in Forschung und klinik, 4th ed.; Enke: Stuttgart, Germany, 2014. [Google Scholar]
- Smit, A.; van der Bank, H.; Falk, T.; de Castro, A. Biochemical genetic markers to identify two morphologically similar south african mastomys species (rodentia: Muridae). Biochem. Syst. Ecol. 2001, 29, 21–30. [Google Scholar] [CrossRef]
- Smit, A.A.; van der Bank, H.F. Isozyme and allozyme markers distinguishing two morphologically similar, medically important mastomys species (rodentia: Muridae). BMC Genet. 2001, 2, 15. [Google Scholar] [CrossRef]
- Kruppa, T.F.; Iglauer, F.; Ihnen, E.; Miller, K.; Kunstyr, I. Mastomys natalensis or mastomys coucha. Correct species designation in animal experiments. Trop. Med. Parasitol. 1990, 41, 219–220. [Google Scholar]
- Britton-Davidian, J.; Catalan, J.; Granjon, L.; Duplantier, J.M. Chromosomal phylogeny and evolution in the genus mastomys (mammalia, rodentia). J. Mammal. 1995, 76, 248–262. [Google Scholar] [CrossRef]
- Hallett, J.M. Cytological and Cytogenetical Studies on the Multimammate Mouse Praomys (Mastomys) Natalensis. Ph.D. Thesis, University of the Witwatersrand, Johannesburg, South Africa, 1977. [Google Scholar]
- Davis, D.H.S.; Heisch, R.B.; Mcneill, D.; Meyer, K.F. Serological survey of plague in rodents and other small mammals in kenya. Trans. R. Soc. Trop. Med. Hyg. 1968, 62, 838–861. [Google Scholar] [CrossRef]
- Pugh, A.O.; Parker, D.A. Plague: Rhodesia’s first recorded outbreak. Cent. Afr. J. Med. 1975, 21, 93–96. [Google Scholar]
- Hallett, A.F. Evaluation of live attenuated plague vaccines in praomys (mastomys) natalensis. Infect. Immun. 1977, 18, 8–13. [Google Scholar] [PubMed]
- Matthey, R. Cytogénétique et taxonomie des rats appartenant au sous-genre mastomys thomas (rodentia-muridae). Mammalia 1966, 30, 105–119. [Google Scholar] [CrossRef]
- Lyons, N.F.; Green, C.R.; Gordon, D.H.; Walters, C.R. G-banding chromosome analysis of Praomys natalensis-(smith) (Rodentia muridae) from rhodesia. 1. 36-chromosome population. Heredity 1977, 38, 197. [Google Scholar] [CrossRef]
- Isaacson, M.; Taylor, P.; Arntzen, L. Ecology of plague in africa: Response of indigenous wild rodents to experimental plague infection. Bull. World Health Organ. 1983, 61, 339–344. [Google Scholar]
- Arntzen, L.; Wadee, A.A.; Isaacson, M. Immune responses of two mastomys sibling species to yersinia pestis. Infect. Immun. 1991, 59, 1966–1971. [Google Scholar]
- Green, C.A.; Keogh, H.; Gordon, D.H.; Pinto, M.; Hartwig, E.K. The distribution, identification, and naming of the mastomys natalensis species complex in southern africa (rodentia: Muridae). J. Zool. 1980, 192, 17–23. [Google Scholar] [CrossRef]
- Frame, J.D.; Baldwin, J.M., Jr.; Gocke, D.J.; Troup, J.M. Lassa fever, a new virus disease of man from west africa. I. Clinical description and pathological findings. Am. J. Trop. Med. Hyg. 1970, 19, 670–676. [Google Scholar] [CrossRef]
- Monath, T.P.; Newhouse, V.F.; Kemp, G.E.; Setzer, H.W.; Cacciapuoti, A. Lassa virus isolation from mastomys natalensis rodents during an epidemic in sierra leone. Science 1974, 185, 263–265. [Google Scholar] [CrossRef]
- Lecompte, E.; Fichet-Calvet, E.; Daffis, S.; Koulemou, K.; Sylla, O.; Kourouma, F.; Dore, A.; Soropogui, B.; Aniskin, V.; Allali, B.; et al. Mastomys natalensis and lassa fever, west africa. Emerg. Infect. Dis. 2006, 12, 1971–1974. [Google Scholar] [CrossRef]
- Gryseels, S.; Baird, S.J.; Borremans, B.; Makundi, R.; Leirs, H.; Gouy de Bellocq, J. When viruses don’t go viral: The importance of host phylogeographic structure in the spatial spread of arenaviruses. PLoS Pathog. 2017, 13, e1006073. [Google Scholar] [CrossRef] [PubMed]
- Mari Saez, A.; Cherif Haidara, M.; Camara, A.; Kourouma, F.; Sage, M.; Magassouba, N.; Fichet-Calvet, E. Rodent control to fight lassa fever: Evaluation and lessons learned from a 4-year study in upper guinea. PLoS Negl. Trop. Dis. 2018, 12, e0006829. [Google Scholar] [CrossRef] [PubMed]
- Lämmler, G.; Zahner, H.; Texdorf, I. Infektionsversuche mit darmnematoden, cestoden und trematoden bei mastomys natalensis (smith, 1834). Zeitschrift für Parasitenkunde 1968, 31, 166–202. [Google Scholar]
- Lammler, G.; Petranyi, G. Chemotherapeutic studies on experimental schistosoma mansoni infection of mastomys natalensis. Bull. World Health Organ. 1971, 44, 739–750. [Google Scholar] [PubMed]
- Kushwaha, V.; Saxena, K.; Verma, R.; Verma, S.K.; Katoch, D.; Kumar, N.; Lal, B.; Murthy, P.K.; Singh, B. Antifilarial activity of diterpenoids from taxodium distichum. Parasites Vectors 2016, 9, 312. [Google Scholar] [CrossRef] [PubMed]
- McNulty, S.N.; Mitreva, M.; Weil, G.J.; Fischer, P.U. Inter and intra-specific diversity of parasites that cause lymphatic filariasis. Infect. Genet. Evol. 2013, 14, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Saunders, M.; Taubert, A.; Dafaalla, T.; Zahner, H. Effect of chemotherapeutic treatment on cytokine (IFN-γ, il-2, il-4, il-5, il-10) gene transcription in response to specific antigens in brugia malayi-infected mastomys coucha. Parasitol. Res. 2008, 103, 1163–1176. [Google Scholar] [CrossRef]
- Singh, P.K.; Kushwaha, S.; Rana, A.K.; Misra-Bhattacharya, S. Cofactor independent phosphoglycerate mutase of brugia malayi induces a mixed Th1/Th2 type immune response and inhibits larval development in the host. BioMed Res. Int. 2014, 2014, 590281. [Google Scholar]
- Verma, S.K.; Joseph, S.K.; Verma, R.; Kushwaha, V.; Parmar, N.; Yadav, P.K.; Thota, J.R.; Kar, S.; Murthy, P.K. Protection against filarial infection by 45–49 kda molecules of brugia malayi via IFN-γ-mediated inos induction. Vaccine 2015, 33, 527–534. [Google Scholar] [CrossRef]
- Verma, S.K.; Kushwaha, V.; Dubey, V.; Saxena, K.; Sharma, A.; Murthy, P.K. Inflammatory mediator release by brugia malayi from macrophages of susceptible host mastomys coucha and Thp-1 and raw 264.7 cell lines. Asian Pac. J. Trop. Med. 2011, 4, 92–96. [Google Scholar] [CrossRef]
- Madhumathi, J.; Prince, P.R.; Rao, D.N.; Karande, A.A.; Reddy, M.V.; Kaliraj, P. Epitope mapping of brugia malayi Alt-2 and the development of a multi-epitope vaccine for lymphatic filariasis. J. Helminthol. 2017, 91, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Kushwaha, S.; Singh, P.K.; Rana, A.K.; Misra-Bhattacharya, S. Immunization of mastomys coucha with brugia malayi recombinant trehalose-6-phosphate phosphatase results in significant protection against homologous challenge infection. PLoS ONE 2013, 8, e72585. [Google Scholar] [CrossRef] [PubMed]
- Kushwaha, V.; Kumar, V.; Verma, S.K.; Sharma, R.; Siddiqi, M.I.; Murthy, P.K. Disorganized muscle protein-1 (dim-1) of filarial parasite brugia malayi: Cdna cloning, expression, purification, structural modeling and its potential as vaccine candidate for human filarial infection. Vaccine 2014, 32, 1693–1699. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.K.; Arora, A.; Murthy, P.K. Recombinant calponin of human filariid brugia malayi: Secondary structure and immunoprophylactic potential. Vaccine 2017, 35, 5201–5208. [Google Scholar] [CrossRef] [PubMed]
- Gupta, J.; Misra, S.; Misra-Bhattacharya, S. Immunization with brugia malayi myosin as heterologous DNA prime protein boost induces protective immunity against b. Malayi infection in mastomys coucha. PLoS ONE 2016, 11, e0164991. [Google Scholar] [CrossRef] [PubMed]
- Modlin, I.M.; Esterline, W.; Kim, H.; Goldenring, J.R. Enterochromaffin-like cells and gastric argyrophil carcinoidosis. Acta Oncol. 1991, 30, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, O.; Wangberg, B.; Johansson, L.; Modlin, I.M.; Ahlman, H. Praomys (mastomys) natalensis: A model for gastric carcinoid formation. Yale J. Biol. Med. 1992, 65, 741–751, discussion 827–749. [Google Scholar]
- Oettle, A.G. Spontaneous carcinoma of the glandular stomach in rattus (mastomys) natalensis, an african rodent. Br. J. Cancer 1957, 11, 415–433. [Google Scholar] [CrossRef]
- Randeria, J.D. Animal model: Carcinoids and adenocarcinoma of the glandular stomach of praomys (mastomys) natalensis. Am. J. Pathol. 1979, 96, 359–362. [Google Scholar]
- Wardlaw, R.; Smith, J.W. Gastric carcinoid tumors. Ochsner J. 2008, 8, 191–196. [Google Scholar] [PubMed]
- Simmers, M.H.; Ibsen, K.H.; Berk, J.E. Concerning the incidence of “spontaneous” stomach cancer in praomys (mastomys) natalensis. Cancer Res. 1968, 28, 1573–1576. [Google Scholar] [PubMed]
- Hooi, J.K.Y.; Lai, W.Y.; Ng, W.K.; Suen, M.M.Y.; Underwood, F.E.; Tanyingoh, D.; Malfertheiner, P.; Graham, D.Y.; Wong, V.W.S.; Wu, J.C.Y.; et al. Global prevalence of helicobacter pylori infection: Systematic review and meta-analysis. Gastroenterology 2017, 153, 420–429. [Google Scholar] [CrossRef] [PubMed]
- De Martel, C.; Ferlay, J.; Franceschi, S.; Vignat, J.; Bray, F.; Forman, D.; Plummer, M. Global burden of cancers attributable to infections in 2008: A review and synthetic analysis. Lancet Oncol. 2012, 13, 607–615. [Google Scholar] [CrossRef]
- Moss, S.F. The clinical evidence linking helicobacter pylori to gastric cancer. Cell. Mol. Gastroenterol. Hepatol. 2017, 3, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Koga, T.; Takahashi, K.; Sato, K.; Kikuchi, I.; Okazaki, Y.; Miura, T.; Katsuta, M.; Narita, T. The effect of colonisation by helicobacter pylori in praomys (mastomys) natalensis on the incidence of carcinoids. J. Med. Microbiol. 2002, 51, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Kidd, M.; Modlin, I.M.; Eick, G.N.; Champaneria, M.C. Isolation, functional characterization, and transcriptome of mastomys ileal enterochromaffin cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 291, G778–G791. [Google Scholar] [CrossRef]
- Kidd, M.; Modlin, I.M.; Eick, G.N.; Camp, R.L.; Mane, S.M. Role of CCN2/CTGF in the proliferation of mastomys enterochromaffin-like cells and gastric carcinoid development. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G191–G200. [Google Scholar] [CrossRef]
- Kidd, M.; Tang, L.H.; Schmid, S.; Lauffer, J.; Louw, J.A.; Modlin, I.M. Helicobacter pylori lipopolysaccharide alters ecl cell DNA synthesis via a CD14 receptor and polyamine pathway in mastomys. Digestion 2000, 62, 217–224. [Google Scholar] [CrossRef]
- Shen, Z.; Xu, S.; Dewhirst, F.E.; Paster, B.J.; Pena, J.A.; Modlin, I.M.; Kidd, M.; Fox, J.G. A novel enterohepatic helicobacter species “helicobacter mastomyrinus” isolated from the liver and intestine of rodents. Helicobacter 2005, 10, 59–70. [Google Scholar] [CrossRef]
- Eaton, K.A.; Opp, J.S.; Gray, B.M.; Bergin, I.L.; Young, V.B. Ulcerative typhlocolitis associated with helicobacter mastomyrinus in telomerase-deficient mice. Vet. Pathol. 2011, 48, 713–725. [Google Scholar] [CrossRef] [PubMed]
- Harvey, J.J. Susceptibility of praomys (mastomys) natalensis to the murine sarcoma virus-harvey (MSV-h). Int. J. Cancer. 1968, 3, 634–643. [Google Scholar] [CrossRef]
- Haag, A.; Wayss, K.; Rommelaere, J.; Cornelis, J.J. Experimentally induced infection with autonomous parvoviruses, minute virus of mice and h-1, in the african multimammate mouse (mastomys coucha). Comp. Med. 2000, 50, 613–621. [Google Scholar] [PubMed]
- Orba, Y.; Kobayashi, S.; Nakamura, I.; Ishii, A.; Hang’ombe, B.M.; Mweene, A.S.; Thomas, Y.; Kimura, T.; Sawa, H. Detection and characterization of a novel polyomavirus in wild rodents. J. Gen. Virol. 2011, 92, 789–795. [Google Scholar] [CrossRef] [PubMed]
- Gardner, S.D.; Field, A.M.; Coleman, D.V.; Hulme, B. New human papovavirus (b.K.) isolated from urine after renal transplantation. Lancet 1971, 1, 1253–1257. [Google Scholar] [CrossRef]
- Padgett, B.L.; Walker, D.L.; ZuRhein, G.M.; Eckroade, R.J.; Dessel, B.H. Cultivation of papova-like virus from human brain with progressive multifocal leucoencephalopathy. Lancet 1971, 1, 1257–1260. [Google Scholar] [CrossRef]
- Feng, H.; Shuda, M.; Chang, Y.; Moore, P.S. Clonal integration of a polyomavirus in human merkel cell carcinoma. Science 2008, 319, 1096–1100. [Google Scholar] [CrossRef]
- Polyomaviridae Study Group of the International Committee on Taxonomy of Viruses; Calvignac-Spencer, S.; Feltkamp, M.C.; Daugherty, M.D.; Moens, U.; Ramqvist, T.; Johne, R.; Ehlers, B. A taxonomy update for the family polyomaviridae. Arch. Virol. 2016, 161, 1739–1750. [Google Scholar] [CrossRef]
- Nafz, J.; Kohler, A.; Ohnesorge, M.; Nindl, I.; Stockfleth, E.; Rösl, F. Persistence of mastomys natalensis papillomavirus in multiple organs identifies novel targets for infection. J. Gen. Virol. 2007, 88, 2670–2678. [Google Scholar] [CrossRef]
- Burtscher, H.; Grunberg, W.; Meingassner, G. Infectious keratoacanthomas of the epidermis in praomys (mastomys) natalensis. Die Nat. 1973, 60, 209–210. [Google Scholar] [CrossRef]
- Amtmann, E.; Wayss, K. Papillomaviruses and carcinogenic progression ii. In The Papovaviridae: The Papillomaviruses; Salzman, N.P., Howley, P.M., Eds.; Springer: Boston, MA, USA, 1987; pp. 187–198. [Google Scholar]
- Inglis, T.J. Principia aetiologica: Taking causality beyond koch’s postulates. J. Med. Microbiol. 2007, 56, 1419–1422. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, R.; Hundeiker, M. Keratoacanthomas in mastomys natalensis. Arch. Dermatol. Res. 1975, 254, 239–243. [Google Scholar] [CrossRef]
- Rudolph, R.; Müller, H. Induktion von epidermalem tumorwachstum in der haut von mastomys natalensis durch übertragung virushaltigen tumorgewebes eines plattenepithelkarzinoms. Zentralblatt für Veterinärmedizin Reihe B 1976, 23, 143–150. [Google Scholar] [CrossRef]
- Rudolph, R.; Thiel, W. Pathological anatomy and histology of spontaneous, epithelial skin tumors in mastomys natalensis. Zentralblatt für Veterinärmedizin Reihe A 1976, 23, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Müller, H.; Gissmann, L. Mastomys natalensis papilloma virus (mnpv), the causative agent of epithelial proliferations: Characterization of the virus particle. J. Gen. Virol. 1978, 41, 315–323. [Google Scholar] [CrossRef]
- Reinacher, M.; Müller, H.; Thiel, W.; Rudolph, R.L. Localization of papillomavirus and virus-specific antigens in the skin of tumor-bearing mastomys natalensis (gra giessen). Med. Microbiol. Immunol. 1978, 165, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.H.; Tachezy, R.; Van Ranst, M.; Chan, S.Y.; Bernard, H.U.; Burk, R.D. The mastomys natalensis papillomavirus: Nucleotide sequence, genome organization, and phylogenetic relationship of a rodent papillomavirus involved in tumorigenesis of cutaneous epithelia. Virology 1994, 198, 534–541. [Google Scholar] [CrossRef] [PubMed]
- De Villiers, E.M.; Fauquet, C.; Broker, T.R.; Bernard, H.U.; zur Hausen, H. Classification of papillomaviruses. Virology 2004, 324, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, J.C.; Calonje, E. Cutaneous manifestations of human papillomaviruses: A review. Acta Dermatovenerol. Alp. Pannon. Adriat. 2011, 20, 145–154. [Google Scholar]
- Wang, J.; Aldabagh, B.; Yu, J.; Arron, S.T. Role of human papillomavirus in cutaneous squamous cell carcinoma: A meta-analysis. J. Am. Acad. Dermatol. 2014, 70, 621–629. [Google Scholar] [CrossRef]
- Pfister, H.; Fuchs, P.G.; Majewski, S.; Jablonska, S.; Pniewska, I.; Malejczyk, M. High prevalence of epidermodysplasia verruciformis-associated human papillomavirus DNA in actinic keratoses of the immunocompetent population. Arch. Dermatol. Res. 2003, 295, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Takai, T. Advances in histopathological diagnosis of keratoacanthoma. J. Dermatol. 2017, 44, 304–314. [Google Scholar] [CrossRef] [PubMed]
- Hannigan, G.D.; Meisel, J.S.; Tyldsley, A.S.; Zheng, Q.; Hodkinson, B.P.; SanMiguel, A.J.; Minot, S.; Bushman, F.D.; Grice, E.A. The human skin double-stranded DNA virome: Topographical and temporal diversity, genetic enrichment, and dynamic associations with the host microbiome. MBio 2015, 6, e01578-15. [Google Scholar] [CrossRef] [PubMed]
- Antonsson, A.; Karanfilovska, S.; Lindqvist, P.G.; Hansson, B.G. General acquisition of human papillomavirus infections of skin occurs in early infancy. J. Clin. Microbiol. 2003, 41, 2509–2514. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, K.; Neumann, J.; Waterboer, T.; Rösl, F. Serological markers for papillomavirus infection and skin tumour development in the rodent model mastomys coucha. J. Gen. Virol. 2011, 92, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Wayss, K.; Reyes-Mayes, D.; Volm, M. Chemical carcinogenesis by the two-stage protocol in the skin ofmastomys natalensis (muridae) using topical initiation with 7, 12-dimethylbenz (a) anthracene and topical promotion with 12-0-tetradecanoylphorbol-13-acetate. Virchows Arch. B Cell Pathol. 1981, 38, 13–21. [Google Scholar] [CrossRef]
- Amtmann, E.; Volm, M.; Wayss, K. Tumour induction in the rodent mastomys natalensis by activation of endogenous papilloma virus genomes. Nature 1984, 308, 291–292. [Google Scholar] [CrossRef] [PubMed]
- Helfrich, I.; Chen, M.; Schmidt, R.; Furstenberger, G.; Kopp-Schneider, A.; Trick, D.; Grone, H.J.; Zur Hausen, H.; Rösl, F. Increased incidence of squamous cell carcinomas in mastomys natalensis papillomavirus e6 transgenic mice during two-stage skin carcinogenesis. J. Virol. 2004, 78, 4797–4805. [Google Scholar] [CrossRef]
- Abel, E.L.; Angel, J.M.; Kiguchi, K.; DiGiovanni, J. Multi-stage chemical carcinogenesis in mouse skin: Fundamentals and applications. Nat. Protoc. 2009, 4, 1350–1362. [Google Scholar] [CrossRef]
- Holderfield, M.; Lorenzana, E.; Weisburd, B.; Lomovasky, L.; Boussemart, L.; Lacroix, L.; Tomasic, G.; Favre, M.; Vagner, S.; Robert, C.; et al. Vemurafenib cooperates with hpv to promote initiation of cutaneous tumors. Cancer Res. 2014, 74, 2238–2245. [Google Scholar] [CrossRef]
- Hasche, D.; Vinzon, S.E.; Rosl, F. Cutaneous papillomaviruses and non-melanoma skin cancer: Causal agents or innocent bystanders? Front. Microbiol. 2018, 9, 874. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, M.; Munzel, P.A.; Braeuning, A. Non-melanoma skin cancer in mouse and man. Arch. Toxicol. 2013, 87, 783–798. [Google Scholar] [CrossRef]
- Siegsmund, M.; Wayss, K.; Amtmann, E. Activation of latent papillomavirus genomes by chronic mechanical irritation. J. Gen. Virol. 1991, 72, 2787–2789. [Google Scholar] [CrossRef] [PubMed]
- Hufbauer, M.; Cooke, J.; van der Horst, G.T.; Pfister, H.; Storey, A.; Akgül, B. Human papillomavirus mediated inhibition of DNA damage sensing and repair drives skin carcinogenesis. Mol. Cancer 2015, 14, 183. [Google Scholar] [CrossRef]
- Viarisio, D.; Müller-Decker, K.; Accardi, R.; Robitaille, A.; Durst, M.; Beer, K.; Jansen, L.; Flechtenmacher, C.; Bozza, M.; Harbottle, R.; et al. Beta hpv38 oncoproteins act with a hit-and-run mechanism in ultraviolet radiation-induced skin carcinogenesis in mice. PLoS Pathog. 2018, 14, e1006783. [Google Scholar] [CrossRef] [PubMed]
- Uberoi, A.; Yoshida, S.; Frazer, I.H.; Pitot, H.C.; Lambert, P.F. Role of ultraviolet radiation in papillomavirus-induced disease. PLoS Pathog. 2016, 12, e1005664. [Google Scholar] [CrossRef] [PubMed]
- Hasche, D.; Stephan, S.; Braspenning-Wesch, I.; Mikulec, J.; Niebler, M.; Gröne, H.J.; Flechtenmacher, C.; Akgül, B.; Rösl, F.; Vinzón, S.E. The interplay of UV and cutaneous papillomavirus infection in skin cancer development. PLoS Pathog. 2017, 13, e1006723. [Google Scholar] [CrossRef]
- Marcuzzi, G.P.; Hufbauer, M.; Kasper, H.U.; Weissenborn, S.J.; Smola, S.; Pfister, H. Spontaneous tumour development in human papillomavirus type 8 e6 transgenic mice and rapid induction by UV-light exposure and wounding. J. Gen. Virol. 2009, 90, 2855–2864. [Google Scholar] [CrossRef]
- Salvermoser, M.; Chotewutmontri, S.; Braspenning-Wesch, I.; Hasche, D.; Rösl, F.; Vinzon, S.E. Transcriptome analysis of mastomys natalensis papillomavirus in productive lesions after natural infection. J. Gen. Virol. 2016, 97, 1658–1669. [Google Scholar] [CrossRef]
- Hasche, D.; Stephan, S.; Savelyeva, L.; Westermann, F.; Rösl, F.; Vinzón, S.E. Establishment of an immortalized skin keratinocyte cell line derived from the animal model mastomys coucha. PLoS ONE 2016, 11, e0161283. [Google Scholar] [CrossRef]
- Nafz, J.; Schäfer, K.; Chen, S.F.; Bravo, I.G.; Ibberson, M.; Nindl, I.; Stockfleth, E.; Rösl, F. A novel rodent papillomavirus isolated from anogenital lesions in its natural host. Virology 2008, 374, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, K.; Waterboer, T.; Rösl, F. A capture elisa for monitoring papillomavirus-induced antibodies in mastomys coucha. J. Virol. Methods 2010, 163, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Andersson, K.; Michael, K.M.; Luostarinen, T.; Waterboer, T.; Gislefoss, R.; Hakulinen, T.; Forslund, O.; Pawlita, M.; Dillner, J. Prospective study of human papillomavirus seropositivity and risk of nonmelanoma skin cancer. Am. J. Epidemiol. 2012, 175, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Ahituv, N. Welcome to the Mastomys Genome Project. Available online: http://mastomys.ucsf.edu (accessed on 12 February 2019).
- Vinzón, S.E.; Rösl, F. Hpv vaccination for prevention of skin cancer. Hum. Vaccines Immunother. 2015, 11, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Garrett, G.L.; Blanc, P.D.; Boscardin, J.; Lloyd, A.A.; Ahmed, R.L.; Anthony, T.; Bibee, K.; Breithaupt, A.; Cannon, J.; Chen, A.; et al. Incidence of and risk factors for skin cancer in organ transplant recipients in the united states. JAMA Dermatol. 2017, 153, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Garrett, G.L.; Yuan, J.T.; Shin, T.M.; Arron, S.T.; Transplant Skin Cancer, N. Validity of skin cancer malignancy reporting to the organ procurement transplant network: A cohort study. J. Am. Acad. Dermatol. 2018, 78, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Rangwala, S.; Tsai, K.Y. Roles of the immune system in skin cancer. Br. J. Dermatol. 2011, 165, 953–965. [Google Scholar] [CrossRef]
- Neale, R.E.; Weissenborn, S.; Abeni, D.; Bavinck, J.N.; Euvrard, S.; Feltkamp, M.C.; Green, A.C.; Harwood, C.; de Koning, M.; Naldi, L.; et al. Human papillomavirus load in eyebrow hair follicles and risk of cutaneous squamous cell carcinoma. Cancer Epidemiol. Prev. Biomark. 2013, 22, 719–727. [Google Scholar] [CrossRef]
- Buck, C.B.; Pastrana, D.V.; Lowy, D.R.; Schiller, J.T. Generation of hpv pseudovirions using transfection and their use in neutralization assays. Methods Mol. Med. 2005, 119, 445–462. [Google Scholar]
- Vinzón, S.E.; Braspenning-Wesch, I.; Muller, M.; Geissler, E.K.; Nindl, I.; Grone, H.J.; Schafer, K.; Rösl, F. Protective vaccination against papillomavirus-induced skin tumors under immunocompetent and immunosuppressive conditions: A preclinical study using a natural outbred animal model. PLoS. Pathog. 2014, 10, e1003924. [Google Scholar] [CrossRef]
- Nagarajan, P.; Asgari, M.M.; Green, A.C.; Guhan, S.M.; Arron, S.T.; Proby, C.M.; Rollison, D.E.; Harwood, C.A.; Toland, A.E. Keratinocyte carcinomas: Current concepts and future research priorities. Clin. Cancer Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Andersson, K.; Waterboer, T.; Kirnbauer, R.; Slupetzky, K.; Iftner, T.; de Villiers, E.M.; Forslund, O.; Pawlita, M.; Dillner, J. Seroreactivity to cutaneous human papillomaviruses among patients with nonmelanoma skin cancer or benign skin lesions. Cancer Epidemiol. Prev. Biomark. 2008, 17, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Missero, C.; Antonini, D. Crosstalk among p53 family members in cutaneous carcinoma. Exp. Dermatol. 2014, 23, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Flores, A.; Grant, W.; White, A.C.; Scumpia, P.; Takahashi, R.; Lowry, W.E. Tumor suppressor identity can contribute to heterogeneity of phenotype in hair follicle stem cell induced squamous cell carcinoma. Exp. Dermatol. 2016, 25, 733. [Google Scholar] [CrossRef] [PubMed]
- Muller, P.A.; Vousden, K.H.; Norman, J.C. P53 and its mutants in tumor cell migration and invasion. J. Cell Biol. 2011, 192, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Schellenbacher, C.; Roden, R.B.S.; Kirnbauer, R. Developments in l2-based human papillomavirus (HPV) vaccines. Virus Res. 2017, 231, 166–175. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasche, D.; Rösl, F. Mastomys Species as Model Systems for Infectious Diseases. Viruses 2019, 11, 182. https://doi.org/10.3390/v11020182
Hasche D, Rösl F. Mastomys Species as Model Systems for Infectious Diseases. Viruses. 2019; 11(2):182. https://doi.org/10.3390/v11020182
Chicago/Turabian StyleHasche, Daniel, and Frank Rösl. 2019. "Mastomys Species as Model Systems for Infectious Diseases" Viruses 11, no. 2: 182. https://doi.org/10.3390/v11020182
APA StyleHasche, D., & Rösl, F. (2019). Mastomys Species as Model Systems for Infectious Diseases. Viruses, 11(2), 182. https://doi.org/10.3390/v11020182