Tropism of the Chikungunya Virus

 , , ,
, , , 
Abstract
1. Introduction
2. Invertebrate and Vertebrate Animal Hosts
2.1. Invetebrate Vectors
2.2. Vertebrate Animal Hosts Spectrum
3. CHIKV Cellular Receptors
4. Human Infection
4.1. Stages and Clinical Signs of Chikungunya Infection
4.2. Host Pathogen Interaction
4.3. Cellular and Tissue Tropism in Human Infections
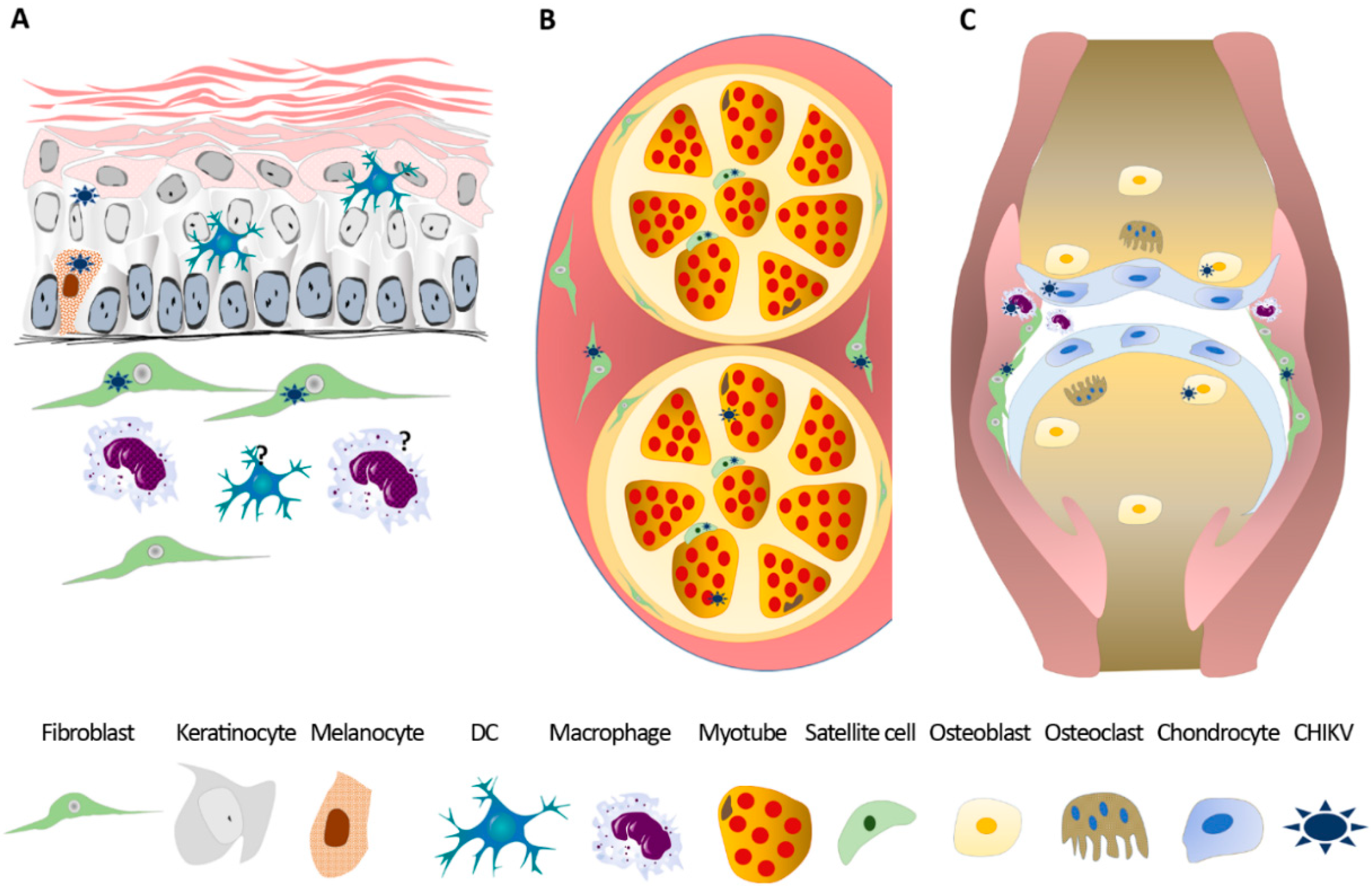
4.3.1. Infection of Skin and Blood Cells
4.3.2. Infection of Muscle, Joint, and Bone
4.3.3. Infection of the Nervous System
4.3.4. Infection of other Target Organs
4.3.5. Target Organs upon Non-Arthropod-Borne Transmission
4.4. Preventive and Therapeutic Strategies
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ross, R.W. The Newala epidemic. III. The virus: Isolation, pathogenic properties and relationship to the epidemic. J. Hyg. 1956, 54, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Weinbren, M.P. The occurrence of Chikungunya virus in Uganda. II. In man on the Entebbe peninsula. Trans. R. Soc. Trop. Med. Hyg. 1958, 52, 258–259. [Google Scholar] [PubMed]
- Powers, A.M.; Brault, A.C.; Tesh, R.B.; Weaver, S.C. Re-emergence of chikungunya and o’nyong-nyong viruses: Evidence for distinct geographical lineages and distant evolutionary relationships. J. Gen. Virol. 2000, 81, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Volk, S.M.; Chen, R.; Tsetsarkin, K.A.; Adams, A.P.; Garcia, T.I.; Sall, A.A.; Nasar, F.; Schuh, A.J.; Holmes, E.C.; Higgs, S.; et al. Genome-Scale Phylogenetic Analyses of Chikungunya Virus Reveal Independent Emergences of Recent Epidemics and Various Evolutionary Rates. J. Virol. 2010, 84, 6497–6504. [Google Scholar] [CrossRef] [PubMed]
- Schuffenecker, I.; Iteman, I.; Michault, A.; Murri, S.; Frangeul, L.; Vaney, M.-C.; Lavenir, R.; Pardigon, N.; Reynes, J.-M.; Pettinelli, F.; et al. Genome Microevolution of Chikungunya Viruses Causing the Indian Ocean Outbreak. PLoS Med. 2006, 3, e263. [Google Scholar] [CrossRef] [PubMed]
- Rezza, G.; Nicoletti, L.; Angelini, R.; Romi, R.; Finarelli, A.; Panning, M.; Cordioli, P.; Fortuna, C.; Boros, S.; Magurano, F.; et al. Infection with chikungunya virus in Italy: An outbreak in a temperate region. Lancet 2007, 370, 1840–1846. [Google Scholar] [CrossRef]
- Aubry, M.; Teissier, A.; Roche, C.; Richard, V.; Yan, A.S.; Zisou, K.; Rouault, E.; Maria, V.; Lastère, S.; Cao-Lormeau, V.-M.; et al. Chikungunya Outbreak, French Polynesia, 2014. Emerg. Infect. Dis. 2015, 21, 724–726. [Google Scholar] [CrossRef] [PubMed]
- Burt, F.J.; Chen, W.; Miner, J.J.; Lenschow, D.J.; Merits, A.; Schnettler, E.; Kohl, A.; Rudd, P.A.; Taylor, A.; Herrero, L.J.; et al. Chikungunya virus: An update on the biology and pathogenesis of this emerging pathogen. Lancet Infect. Dis. 2017, 17, e107–e117. [Google Scholar] [CrossRef]
- Nunes, M.R.T.; Faria, N.R.; de Vasconcelos, J.M.; Golding, N.; Kraemer, M.U.; de Oliveira, L.F.; Azevedo, R.; da Silva Azevedo, R.D.S.; da Silva, D.E.A.; da Silva, E.V.P.; et al. Emergence and potential for spread of Chikungunya virus in Brazil. BMC Med. 2015, 13, 102. [Google Scholar] [CrossRef]
- Mohamed, N.; Magzoub, M.; Mohamed, R.E.H.; Aleanizy, F.S.; Alqahtani, F.Y.; Nour, B.Y.M.; Alkarsany, M.M.S. Prevalence and identification of arthropod- transmitted viruses in Kassala state, Eastern Sudan. Libyan J. Med. 2019, 14, 1564511. [Google Scholar] [CrossRef]
- WHO Chikungunya—Sudan Disease Outbreak News. Available online: https://www.who.int/csr/don/15-october-2018-chikungunya-sudan/en/ (accessed on 19 February 2019).
- Zeller, H.; Van Bortel, W.; Sudre, B. Chikungunya: Its history in Africa and Asia and its spread to new regions in 2013–2014. J. Infect. Dis. 2016, 214, S436–S440. [Google Scholar] [CrossRef] [PubMed]
- Higgs, S.; Vanlandingham, D. Chikungunya Virus and Its Mosquito Vectors. Vector-Borne Zoonotic Dis. 2015, 15, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Savage, H.M.; Ledermann, J.P.; Yug, L.; Burkhalter, K.L.; Marfel, M.; Hancock, W.T. Incrimination of Aedes (Stegomyia) hensilli Farner as an epidemic vector of Chikungunya virus on Yap Island, Federated States of Micronesia, 2013. Am. J. Trop. Med. Hyg. 2015, 92, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Tesh, R.B.; Gubler, D.J.; Rosen, L. Variation among goegraphic strains of Aedes albopictus in susceptibility to infection with chikungunya virus. Am. J. Trop. Med. Hyg. 1976, 25, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Tsetsarkin, K.A.; Chen, R.; Sherman, M.B.; Weaver, S.C. Chikungunya virus: Evolution and genetic determinants of emergence. Curr. Opin. Virol. 2011, 1, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Tsetsarkin, K.A.; Vanlandingham, D.L.; McGee, C.E.; Higgs, S. A single mutation in chikungunya virus affects vector specificity and epidemic potential. PLoS Pathog. 2007, 3, e201. [Google Scholar] [CrossRef] [PubMed]
- Thiberville, S.-D.; Moyen, N.; Dupuis-Maguiraga, L.; Nougairede, A.; Gould, E.A.; Roques, P.; de Lamballerie, X. Chikungunya fever: Epidemiology, clinical syndrome, pathogenesis and therapy. Antiviral Res. 2013, 99, 345–370. [Google Scholar] [CrossRef]
- Vega-Rua, A.; Zouache, K.; Caro, V.; Diancourt, L.; Delaunay, P.; Grandadam, M.; Failloux, A.-B. High Efficiency of Temperate Aedes albopictus to Transmit Chikungunya and Dengue Viruses in the Southeast of France. PLoS ONE 2013, 8, e59716. [Google Scholar] [CrossRef]
- Grandadam, M.; Caro, V.; Plumet, S.; Thiberge, J.-M.; Souarès, Y.; Failloux, A.-B.; Tolou, H.J.; Budelot, M.; Cosserat, D.; Leparc-Goffart, I.; et al. Chikungunya Virus, Southeastern France. Emerg. Infect. Dis. 2011, 17, 910–913. [Google Scholar] [CrossRef]
- Bordi, L.; Carletti, F.; Lalle, E.; Colavita, F.; Meschi, S.; Di Caro, A.; Nicastri, E.; Scognamiglio, P.; Vairo, F.; Di Lallo, D.; et al. Molecular characterization of autochthonous chikungunya cluster in latium region, Italy. Emerg. Infect. Dis. 2018, 24, 178–180. [Google Scholar] [CrossRef]
- Tsetsarkin, K.A.; Weaver, S.C. Sequential Adaptive Mutations Enhance Efficient Vector Switching by Chikungunya Virus and Its Epidemic Emergence. PLoS Pathog. 2011, 7, e1002412. [Google Scholar] [CrossRef] [PubMed]
- Tsetsarkin, K.A.; Chen, R.; Yun, R.; Rossi, S.L.; Plante, K.S.; Guerbois, M.; Forrester, N.; Perng, G.C.; Sreekumar, E.; Leal, G.; et al. Multi-peaked adaptive landscape for chikungunya virus evolution predicts continued fitness optimization in Aedes albopictus mosquitoes. Nat. Commun. 2014, 5, 4084. [Google Scholar] [CrossRef] [PubMed]
- Arias-Goeta, C.; Mousson, L.; Rougeon, F.; Failloux, A.B. Dissemination and Transmission of the E1-226V Variant of Chikungunya Virus in Aedes albopictus Are Controlled at the Midgut Barrier Level. PLoS ONE 2013, 8, e57548. [Google Scholar] [CrossRef] [PubMed]
- Jupp, P.G.; McIntosh, B.M. Aedes furcifer and other mosquitoes as vectors of chikungunya virus at Mica, northeastern Transvaal, South Africa. J. Am. Mosq. Control Assoc. 1990, 6, 415–420. [Google Scholar] [PubMed]
- Jupp, P.G.; McIntosh, B.M.; Dos Santos, I.; DeMoor, P. Laboratory vector studies on six mosquito and one tick species with chikungunya virus. Trans. R. Soc. Trop. Med. Hyg. 1981, 75, 15–19. [Google Scholar] [CrossRef]
- Diallo, M.; Thonnon, J.; Traore-lamizana, M.; Fontenille, D. Vectors of Chikungunya virus in Senegal: Current data and transmission cycles. Am. J. Trop. Med. Hyg. 1999, 60, 281–286. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, B.M.; Paterson, H.E.; McGillivray, G.; Desousa, J. Further studies on the chikungunya outbreak in Rhodesia. I. Mosquitoes, wild primates and birds in relation to the epidemic. Ann. Trop. Med. Parasitol. 1964, 58, 45–51. [Google Scholar] [CrossRef]
- Weinbren, M.P.; Haddow, A.J.; Williams, M.C. The occurrence of Chikungunya virus in Uganda. I. Isolation from mosquitoes. Trans. R. Soc. Trop. Med. Hyg. 1958, 52, 253–257. [Google Scholar] [CrossRef]
- Diallo, D.; Dia, I.; Diagne, C.T.; Gaye, A.; Diallo, M. Emergences of Chikungunya and Zika in Africa. In Chikungunya and Zika Viruses; Higgs, S., Vanlandingham, D.L., Powers, A.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 87–133. ISBN 9780128118658. [Google Scholar]
- Brès, P.; Camicas, J.L.; Cornet, M.; Robin, Y.; Taufflieb, R. Epidemiology of arbovirus diseases in Senegal. Bull. Soc. Pathol. Exot. Filiales 1969, 62, 253–259. [Google Scholar]
- Konstantinov, O.K. Ticks of the Ixodidae family as reservoir of arboviruses in the Republic of Guinea. II. Arboviruses. Rev. Elev. Med. Vet. Pays Trop. 1990, 43, 15–22. [Google Scholar]
- Coffey, L.L.; Failloux, A.B.; Weaver, S.C. Chikungunya virus–vector interactions. Viruses 2014, 6, 4628–4663. [Google Scholar] [CrossRef] [PubMed]
- Mulwa, F.; Lutomiah, J.; Chepkorir, E.; Okello, S.; Eyase, F.; Tigoi, C.; Kahato, M.; Sang, R. Vector competence of Aedes bromeliae and Aedes vitattus mosquito populations from Kenya for chikungunya virus. PLoS Negl. Trop. Dis. 2018, 12, e0006746. [Google Scholar] [CrossRef] [PubMed]
- Richard, V.; Paoaafaite, T.; Cao-Lormeau, V.-M. Vector Competence of Aedes aegypti and Aedes polynesiensis Populations from French Polynesia for Chikungunya Virus. PLoS Negl. Trop. Dis. 2016, 10, e0004694. [Google Scholar] [CrossRef] [PubMed]
- Mendenhall, I.H.; Manuel, M.; Moorthy, M.; Lee, T.T.M.; Low, D.H.W.; Missé, D.; Gubler, D.J.; Ellis, B.R.; Ooi, E.E.; Pompon, J. Peridomestic Aedes malayensis and Aedes albopictus are capable vectors of arboviruses in cities. PLoS Negl. Trop. Dis. 2017, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Lourenço-de-Oliveira, R.; Failloux, A.B. High risk for chikungunya virus to initiate an enzootic sylvatic cycle in the tropical Americas. PLoS Negl. Trop. Dis. 2017, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- van den Hurk, A.F.; Hall-Mendelin, S.; Pyke, A.T.; Smith, G.A.; Mackenzie, J.S. Vector Competence of Australian Mosquitoes for Chikungunya Virus. Vector-Borne Zoonotic Dis. 2010, 10, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Sudeep, A.B.; Ghodke, Y.S.; George, R.P.; Ingale, V.S.; Dhaigude, S.D.; Gokhale, M.D. Vectorial capacity of Culex gelidus (Theobald) mosquitoes to certain viruses of public health importance in India. J. Vector Borne Dis. 2015, 52, 153–158. [Google Scholar] [PubMed]
- Ciocchetta, S.; Prow, N.A.; Darbro, J.M.; Frentiu, F.D.; Savino, S.; Montarsi, F.; Capelli, G.; Aaskov, J.G.; Devine, G.J. The new European invader Aedes (Finlaya) koreicus: A potential vector of chikungunya virus. Pathog. Glob. Health 2018, 112, 107–114. [Google Scholar] [CrossRef]
- Fros, J.J.; Geertsema, C.; Zouache, K.; Baggen, J.; Domeradzka, N.; Van Leeuwen, D.M.; Flipse, J.; Vlak, J.M.; Failloux, A.B.; Pijlman, G.P. Mosquito Rasputin interacts with chikungunya virus nsP3 and determines the infection rate in Aedes albopictus. Parasites Vectors 2015, 8, 1–15. [Google Scholar] [CrossRef]
- Pesko, K.; Westbrook, C.J.; Mores, C.N.; Lounibos, L.P.; Reiskind, M.H. Effects of infectious virus dose and bloodmeal delivery method on susceptibility of Aedes aegypti and Aedes albopictus to chikungunya virus. J. Med. Entomol. 2009, 46, 395–399. [Google Scholar] [CrossRef]
- Lee, R.C.H.; Hapuarachchi, H.C.; Chen, K.C.; Hussain, K.M.; Chen, H.; Low, S.L.; Ng, L.C.; Lin, R.; Ng, M.M.L.; Chu, J.J.H. Mosquito Cellular Factors and Functions in Mediating the Infectious entry of Chikungunya Virus. PLoS Negl. Trop. Dis. 2013, 7, e2050. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Balaraman, V.; Kantor, A.M.; Lin, J.; Grant, D.A.G.; Held, N.L.; Franz, A.W.E. Chikungunya virus dissemination from the midgut of Aedes aegypti is associated with temporal basal lamina degradation during bloodmeal digestion. PLoS Negl. Trop. Dis. 2017, 11, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Kantor, A.M.; Lin, J.; Passarelli, A.L.; Clem, R.J.; Franz, A.W.E. Infection pattern and transmission potential of chikungunya virus in two New World laboratory-adapted Aedes aegypti strains. Sci. Rep. 2016, 6, 24729. [Google Scholar] [CrossRef]
- Kantor, A.M.; Grant, D.G.; Balaraman, V.; White, T.A.; Franz, A.W.E. Ultrastructural analysis of chikungunya virus dissemination from the midgut of the yellow fever mosquito, aedes aegypti. Viruses 2018, 10, 571. [Google Scholar] [CrossRef] [PubMed]
- Bowers, D.F.; Coleman, C.G.; Brown, D.T. Sindbis virus-associated pathology in Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 2003, 40, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Romoser, W.S.; Wasieloski, L.P.; Pushko, P.; Kondig, J.P.; Lerdthusnee, K.; Neira, M.; Ludwig, G.V. Evidence for arbovirus dissemination conduits from the mosquito (Diptera: Culicidae) midgut. J. Med. Entomol. 2004, 41, 467–475. [Google Scholar] [CrossRef]
- Salazar, M.I.; Richardson, J.H.; Sánchez-Vargas, I.; Olson, K.E.; Beaty, B.J. Dengue virus type 2: Replication and tropisms in orally infected Aedes aegypti mosquitoes. BMC Microbiol. 2007, 7, 9. [Google Scholar] [CrossRef]
- Ziegler, S.A.; Nuckols, J.; McGee, C.E.; Huang, Y.S.; Vanlandingham, D.L.; Tesh, R.B.; Higgs, S. In Vivo Imaging of Chikungunya Virus in Mice and Aedes Mosquitoes Using a Renilla Luciferase Clone. Vector-Borne Zoonotic Dis. 2011, 11, 1471–1477. [Google Scholar] [CrossRef]
- Vega-Rúa, A.; Schmitt, C.; Bonne, I.; Locker, J.K.; Failloux, A.B. Chikungunya virus replication in salivary glands of the mosquito aedes albopictus. Viruses 2015, 7, 5902–5907. [Google Scholar] [CrossRef]
- Tchankouo-Nguetcheu, S.; Bourguet, E.; Lenormand, P.; Rousselle, J.-C.; Namane, A.; Choumet, V. Infection by chikungunya virus modulates the expression of several proteins in Aedes aegypti salivary glands. Parasit. Vectors 2012, 5, 264. [Google Scholar] [CrossRef]
- Le Coupanec, A.; Tchankouo-Nguetcheu, S.; Roux, P.; Khun, H.; Huerre, M.; Morales-Vargas, R.; Enguehard, M.; Lavillette, D.; Missé, D.; Choumet, V. Co-infection of mosquitoes with chikungunya and dengue viruses reveals modulation of the replication of both viruses in midguts and salivary glands of Aedes aegypti mosquitoes. Int. J. Mol. Sci. 2017, 18, 1708. [Google Scholar] [CrossRef] [PubMed]
- Janzen, H.G.; Rhodes, A.J.; Doane, F.W. Chikungunya virus in salivary glands of Aedes aegypti (L.): An electron microscope study. Can. J. Microbiol. 1970, 16, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.-N.; Rodriguez, S.D.; Carpenter, V.K.; Vulcan, J.; Bailey, C.D.; Nageswara-Rao, M.; Li, Y.; Attardo, G.M.; Hansen, I.A. Fat Body Organ Culture System in Aedes Aegypti, a Vector of Zika Virus. J. Vis. Exp. 2017. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.V.; Chan, Y.F.; Sam, I.-C.; Sulaiman, W.Y.W.; Vythilingam, I. Chikungunya Virus Infection of Aedes Mosquitoes. In Chikungunya Virus. Methods in Molecular Biology; Humana Press: New York, NY, USA, 2016; Volume 1426. [Google Scholar]
- Sirisena, P.D.N.N.; Kumar, A.; Sunil, S. Evaluation of Aedes aegypti (Diptera: Culicidae) Life Table Attributes Upon Chikungunya Virus Replication Reveals Impact on Egg-Laying Pathways. J. Med. Entomol. 2018, 55, 1580–1587. [Google Scholar] [CrossRef] [PubMed]
- Thavara, U.; Tawatsin, A.; Pengsakul, T.; Bhakdeenuan, P.; Chanama, S.; Anantapreecha, S.; Molito, C.; Chompoosri, J.; Thammapalo, S.; Sawanpanyalert, P.; et al. Outbreak of chikungunya fever in Thailand and virus detection in field population of vector mosquitoes, Aedes aegypti (L.) and Aedes albopictus Skuse (Diptera: Culicidae). Southeast Asian J. Trop. Med. Public Health 2009, 40, 951–962. [Google Scholar] [PubMed]
- Jain, J.; Kushwah, R.B.S.; Singh, S.S.; Sharma, A.; Adak, T.; Singh, O.P.; Bhatnagar, R.K.; Subbarao, S.K.; Sunil, S. Evidence for natural vertical transmission of chikungunya viruses in field populations of Aedes aegypti in Delhi and Haryana states in India—A preliminary report. Acta Trop. 2016, 162, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Chompoosri, J.; Thavara, U.; Tawatsin, A.; Boonserm, R.; Phumee, A.; Sangkitporn, S.; Siriyasatien, P. Vertical transmission of Indian Ocean Lineage of chikungunya virus in Aedes aegypti and Aedes albopictus mosquitoes. Parasit. Vectors 2016, 9, 227. [Google Scholar] [CrossRef]
- Agarwal, A.; Dash, P.K.; Singh, A.K.; Sharma, S.; Gopalan, N.; Rao, P.V.L.; Parida, M.M.; Reiter, P. Evidence of Experimental Vertical Transmission of Emerging Novel ECSA Genotype of Chikungunya Virus in Aedes aegypti. PLoS Negl. Trop. Dis. 2014, 8, e2990. [Google Scholar] [CrossRef]
- Wong, H.V.; Vythilingam, I.; Sulaiman, W.Y.W.; Lulla, A.; Merits, A.; Chan, Y.F.; Sam, I.C. Detection of persistent chikungunya virus RNA but not infectious virus in experimental vertical transmission in aedes aegypti from Malaysia. Am. J. Trop. Med. Hyg. 2016, 94, 182–186. [Google Scholar] [CrossRef]
- Rosen, L.; Shroyer, D.A.; Tesh, R.B.; Freier, J.E.; Lien, J.C. Transovarial transmission of dengue viruses by mosquitoes: Aedes albopictus and Aedes aegypti. Am. J. Trop. Med. Hyg. 1983, 32, 1108–1119. [Google Scholar] [CrossRef]
- Hardy, J.L.; Houk, E.J.; Kramer, L.D.; Reeves, W.C. Intrinsic factors affecting vector competence of mosquitoes for arboviruses. Annu. Rev. Entomol. 1983, 28, 229–262. [Google Scholar] [CrossRef] [PubMed]
- Mascarenhas, M.; Garasia, S.; Berthiaume, P.; Corrin, T.; Greig, J.; Ng, V.; Young, I.; Waddell, L. A scoping review of published literature on chikungunya virus. PLoS ONE 2018, 13, e0207554. [Google Scholar] [CrossRef] [PubMed]
- Althouse, B.M.; Guerbois, M.; Cummings, D.A.T.; Diop, O.M.; Faye, O.; Faye, A.; Diallo, D.; Sadio, B.D.; Sow, A.; Faye, O.; et al. Role of monkeys in the sylvatic cycle of chikungunya virus in Senegal. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Eastwood, G.; Sang, R.C.; Guerbois, M.; Taracha, E.L.N.; Weaver, S.C. Enzootic circulation of chikungunya virus in East Africa: Serological evidence in non-human Kenyan primates. Am. J. Trop. Med. Hyg. 2017, 97, 1399–1404. [Google Scholar] [CrossRef] [PubMed]
- Osterrieth, P.; Rathe, E.; Deleplanque-Liegeois, P. [Simultaneous isolation of the yellow fever and Chikungunya viruses at Bili (Haut-Uele, Belgian Congo)]. Ann. la Soc. Belge Med. Trop. 1961, 41, 207–212. [Google Scholar]
- Vourc’H, G.; Halos, L.; Desvars, A.; Boué, F.; Pascal, M.; Lecollinet, S.; Zientara, S.; Duval, T.; Nzonza, A.; Brémont, M. Chikungunya antibodies detected in non-human primates and rats in three Indian Ocean islands after the 2006 ChikV outbreak. Vet. Res. 2014, 45, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Kading, R.C.; Borland, E.M.; Cranfield, M.; Powers, A.M. Prevalence of antibodies to alphaviruses and flaviviruses in free-ranging game animals and Non human primates in the greater Congo basin. J. Wildl. Dis. 2013, 49, 587–599. [Google Scholar] [CrossRef]
- McCrae, A.W.; Henderson, B.E.; Kirya, B.G.; Sempala, S.D. Chikungunya virus in the Entebbe area of Uganda: Isolations and epidemiology. Trans. R. Soc. Trop. Med. Hyg. 1971, 65, 152–168. [Google Scholar] [CrossRef]
- Guilherme, J.M.; Gonella-Legall, C.; Legall, F.; Nakoume, E.; Vincent, J. Seroprevalence of five arboviruses in Zebu cattle in the Central African Republic. Trans. R. Soc. Trop. Med. Hyg. 1996, 90, 31–33. [Google Scholar] [CrossRef]
- Wolfe, N.D.; Kilbourn, A.M.; Karesh, W.B.; Rahman, H.A.; Bosi, E.J.; Cropp, B.C.; Andau, M.; Spielman, A.; Gubler, D.J. Sylvatic transmission of arboviruses among bornean orangutans. Am. J. Transplant. 2001, 64, 310–316. [Google Scholar] [CrossRef]
- Sam, I.C.; Chua, C.L.; Rovie-Ryan, J.J.; Fu, J.Y.L.; Tong, C.; Sitam, F.T.; Chan, Y.F. Chikungunya virus in macaques, Malaysia. Emerg. Infect. Dis. 2015, 21, 1683–1685. [Google Scholar] [CrossRef]
- Nakgoi, K.; Nitatpattana, N.; Wajjwalku, W.; Pongsopawijit, P.; Kaewchot, S.; Yoksan, S.; Siripolwat, V.; Souris, M.; Gonzalez, J.P. Dengue, Japanese encephalitis and Chikungunya virus antibody prevalence among captive monkey (Macaca nemestrina) colonies of Northern Thailand. Am. J. Primatol. 2014, 76, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Marchette, N.J.; Rudnick, A.; Garcia, R.; MacVean, D.W. Alphaviruses in Peninusular Malaysia: I. Virus isolations and animal serology. Southeast Asian J. Trop. Med. Public Health 1978, 9, 317–329. [Google Scholar] [PubMed]
- Inoue, S.; Morita, K.; Rr, M.; Jv, T.; Rrg, R. Distribution of three arbovirus antibodies among monkeys in the Philippines.pdf. J. Med. Primatol. 2003, 32, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Apandi, Y.; Nazni, W.A.; Azleen, Z.A.N.; Vythilingam, I.; Noorazian, M.Y.; Azahari, A.H.; Zainah, S.; Lee, H.L. The first isolation of chikungunya virus from non-human primates in Malaysia. J. Gen. Mol. Virol. 2009, 1, 35–39. [Google Scholar]
- Moreira-Soto, A.; Carneiro, I.D.O.; Fischer, C.; Feldmann, M.; Kümmerer, B.M.; Silva, N.S.; Santos, U.G.; Souza, B.F.D.C.D.; Liborio, F.D.A.; Valença-Montenegro, M.M.; et al. Limited Evidence for Infection of Urban and Peri-urban Nonhuman Primates with Zika and Chikungunya Viruses in Brazil. mSphere 2018, 3, e00523-17. [Google Scholar] [CrossRef] [PubMed]
- Bosco-Lauth, A.M.; Hartwig, A.E.; Bowen, R.A. Reptiles and amphibians as potential reservoir hosts of chikungunya virus. Am. J. Trop. Med. Hyg. 2018, 98, 841–844. [Google Scholar] [CrossRef] [PubMed]
- Bosco-Lauth, A.M.; Nemeth, N.M.; Kohler, D.J.; Bowen, R.A. Viremia in North American mammals and birds after experimental infection with chikungunya viruses. Am. J. Trop. Med. Hyg. 2016, 94, 504–506. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, B.M. Antibody against Chikungunya virus in wild primates in Southern Africa. S. Afr. J. Med. Sci. 1970, 35, 65–74. [Google Scholar]
- van Duijl-Richter, M.K.S.; Hoornweg, T.E.; Rodenhuis-Zybert, I.A.; Smit, J.M. Early events in chikungunya virus infection—From virus cell binding to membrane fusion. Viruses 2015, 7, 3647–3674. [Google Scholar] [CrossRef]
- Wintachai, P.; Wikan, N.; Kuadkitkan, A.; Jaimipuk, T.; Ubol, S.; Pulmanausahakul, R.; Auewarakul, P.; Kasinrerk, W.; Weng, W.Y.; Panyasrivanit, M.; et al. Identification of prohibitin as a Chikungunya virus receptor protein. J. Med. Virol. 2012, 84, 1757–1770. [Google Scholar] [CrossRef]
- Wintachai, P.; Thuaud, F.; Basmadjian, C.; Roytrakul, S.; Ubol, S.; Désaubry, L.; Smith, D.R. Assessment of flavaglines as potential chikungunya virus entry inhibitors. Microbiol. Immunol. 2015, 59, 129–141. [Google Scholar] [CrossRef]
- Zhang, R.; Kim, A.S.; Fox, J.M.; Nair, S.; Basore, K.; Klimstra, W.B.; Rimkunas, R.; Fong, R.H.; Lin, H.; Poddar, S.; et al. Mxra8 is a receptor for multiple arthritogenic alphaviruses. Nature 2018, 557, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Desai, A.; Ravi, V.; Narayanappa, G.; Tyagi, B.K. Chikungunya Virus Interacts with Heat Shock Cognate 70 Protein to Facilitate Its Entry into Mosquito Cell Line. Intervirology 2018, 60, 247–262. [Google Scholar] [CrossRef]
- Fongsaran, C.; Jirakanwisal, K.; Kuadkitkan, A.; Wikan, N.; Wintachai, P.; Thepparit, C.; Ubol, S.; Phaonakrop, N.; Roytrakul, S.; Smith, D.R. Involvement of ATP synthase β subunit in chikungunya virus entry into insect cells. Arch. Virol. 2014, 159, 3353–3364. [Google Scholar] [CrossRef] [PubMed]
- Haby, M.M.; Pinart, M.; Elias, V.; Reveiz, L. Prevalence of asymptomatic Zika virus infection: A systematic review. Bull. World Health Organ. 2018, 96, 402. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Ayu, S.M.; Lai, L.R.; Chan, Y.F.; Hatim, A.; Hairi, N.N.; Ayob, A.; Sam, I.-C. Seroprevalence survey of Chikungunya virus in Bagan Panchor, Malaysia. Am. J. Trop. Med. Hyg. 2010, 83, 1245–1248. [Google Scholar] [CrossRef]
- Dupuis-Maguiraga, L.; Noret, M.; Brun, S.; Le Grand, R.; Gras, G.; Roques, P. Chikungunya disease: Infection-associated markers from the acute to the chronic phase of arbovirus-induced arthralgia. PLoS Negl. Trop. Dis. 2012, 6, e1446. [Google Scholar] [CrossRef] [PubMed]
- WHO. Chikungunya Virus Fact Sheet. Available online: https://www.who.int/news-room/fact-sheets/detail/chikungunya (accessed on 19 February 2019).
- Rahman, M.; Jakaria, S.K.; Sayed, B.; Kabir, A.K.M.H.; Mallik, U.; Hasan, R.; Siddique, A.B.; Hossain, A.; Uddin, N.; Hassan, M.; et al. Clinical and Laboratory Characteristics of an Acute Chikungunya Outbreak in Bangladesh in 2017. Am. J. Trop. Med. Hyg. 2019, 100, 405–410. [Google Scholar] [CrossRef]
- De Andrade, G.C.; Ventura, C.V.; Mello Filho, P.A.D.A.; Maia, M.; Vianello, S.; Rodrigues, E.B. Arboviruses and the eye. Int. J. Retin. Vitr. 2017, 3, 4. [Google Scholar] [CrossRef] [PubMed]
- Fourie, E.D.; Morrison, J.G. Rheumatoid arthritic syndrome after chikungunya fever. S. Afr. Med. J. 1979, 56, 130–132. [Google Scholar] [PubMed]
- Aalst, V.; Nelen, C.M.; Goorhuis, A.; Stijnis, C.; Grobusch, M.P. Long-term sequelae of chikungunya virus disease: A systematic review. Travel Med. Infect. Dis. 2017, 15, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Morales, A.J.; Cardona-Ospina, J.A.; Fernanda Urbano-Garzón, S.; Sebastian Hurtado-Zapata, J. Prevalence of Post-Chikungunya Infection Chronic Inflammatory Arthritis: A Systematic Review and Meta-Analysis. Arthritis Care Res. 2016, 68, 1849–1858. [Google Scholar] [CrossRef] [PubMed]
- Economopoulou, A.; Dominguez, M.; Helynck, B.; Sissoko, D.; Wichmann, O.; Quenel, P.; Germonneau, P.; Quatresous, I. Atypical Chikungunya virus infections: Clinical manifestations, mortality and risk factors for severe disease during the 2005–2006 outbreak on Réunion. Epidemiol. Infect. 2009, 137, 534. [Google Scholar] [CrossRef] [PubMed]
- Colavita, F.; Vita, S.; Lalle, E.; Carletti, F.; Bordi, L.; Vincenti, D.; Pozzetto, I.; Aiuti, M.; Vairo, F.; Capobianchi, M.R.; et al. Overproduction of IL-6 and Type-I IFN in a Lethal Case of Chikungunya Virus Infection in an Elderly Man During the 2017 Italian Outbreak. Open Forum Infect. Dis. 2018, 5, ofy276. [Google Scholar] [CrossRef] [PubMed]
- Lemant, J.; Boisson, V.; Winer, A.; Thibault, L.; André, H.; Tixier, F.; Lemercier, M.; Antok, E.; Cresta, M.P.; Grivard, P.; et al. Serious acute chikungunya virus infection requiring intensive care during the reunion island outbreak in 2005–2006. Crit. Care Med. 2008, 36, 2536–2541. [Google Scholar] [CrossRef] [PubMed]
- Renault, P.; Balleydier, E.; D’Ortenzio, E.; Bâville, M.; Filleul, L. Epidemiology of chikungunya infection on Reunion Island, Mayotte, and neighboring countries. Médecine Mal. Infect. 2012, 42, 93–101. [Google Scholar] [CrossRef]
- Rajapakse, S.; Rodrigo, C.; Rajapakse, A. Atypical manifestations of chikungunya infection. Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 89–96. [Google Scholar] [CrossRef]
- Mercado, M.; Acosta-Reyes, J.; Parra, E.; Guzmán, L.; Beltrán, M.; Gasque, P.; Mejía-García, C.; Viasus, D. Renal involvement in fatal cases of chikungunya virus infection. J. Clin. Virol. 2018, 103, 16–18. [Google Scholar] [CrossRef]
- Garg, T.; Sanke, S.; Ahmed, R.; Chander, R.; Basu, S. Stevens-Johnson syndrome and toxic epidermal necrolysis-like cutaneous presentation of chikungunya fever: A case series. Pediatr. Dermatol. 2018, 35, 392–396. [Google Scholar] [CrossRef] [PubMed]
- Dutta, P.; Sharma, A. A case of atypical ophthalmoplegia after Chikungunya fever. Int. Ophthalmol. 2018, 38, 837–839. [Google Scholar] [CrossRef] [PubMed]
- Mehta, R.; Gerardin, P.; de Brito, C.A.A.; Soares, C.N.; Ferreira, M.L.B.; Solomon, T. The neurological complications of chikungunya virus: A systematic review. Rev. Med. Virol. 2018, 28, e1978. [Google Scholar] [CrossRef] [PubMed]
- Sharda, M.; Meena, H. Neurological complications in Chikungunya fever. J. Assoc. Phys. India 2007, 55, 765–769. [Google Scholar]
- Singh, S.S.; Manimunda, S.P.; Sugunan, A.P.; Vijayachari, P. Four cases of acute flaccid paralysis associated with chikungunya virus infection. Epidemiol. Infect. 2008, 136, 1277–1280. [Google Scholar] [CrossRef] [PubMed]
- Wielanek, A.C.; Monredon, J.D.; Amrani, M.E.; Roger, J.C.; Serveaux, J.P. Guillain-Barré syndrome complicating a chikungunya virus infection. Neurology 2007, 69, 2105–2107. [Google Scholar] [CrossRef]
- Acevedo, N.; Waggoner, J.; Rodriguez, M.; Rivera, L.; Landivar, J.; Pinsky, B.; Zambrano, H. Zika virus, chikungunya virus, and dengue virus in cerebrospinal fluid from adults with neurological manifestations, Guayaquil, Ecuador. Front. Microbiol. 2017, 8, 1–6. [Google Scholar] [CrossRef]
- Mehta, R.; Soares, C.N.; Medialdea-Carrera, R.; Ellul, M.; da Silva, M.T.T.; Rosala-Hallas, A.; Jardim, M.R.; Burnside, G.; Pamplona, L.; Bhojak, M.; et al. The spectrum of neurological disease associated with Zika and chikungunya viruses in adults in Rio de Janeiro, Brazil: A case series. PLoS Negl. Trop. Dis. 2018, 12, e0006212. [Google Scholar] [CrossRef]
- Fros, J.; Pijlman, G. Alphavirus Infection: Host Cell Shut-Off and Inhibition of Antiviral Responses. Viruses 2016, 8, 166. [Google Scholar] [CrossRef]
- Li, S.; Gong, M.; Zhao, F.; Shao, J.; Xie, Y.; Zhang, Y.; Chang, H. Type I Interferons: Distinct Biological Activities and Current Applications for Viral Infection. Cell. Physiol. Biochem. 2018, 51, 2377–2396. [Google Scholar] [CrossRef]
- Schilte, C.; Couderc, T.; Chretien, F.; Sourisseau, M.; Gangneux, N.; Guivel-Benhassine, F.; Kraxner, A.; Tschopp, J.; Higgs, S.; Michault, A.; et al. Type I IFN controls chikungunya virus via its action on nonhematopoietic cells. J. Exp. Med. 2010, 207, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Broeckel, R.; Haese, N.; Messaoudi, I.; Streblow, D. Nonhuman Primate Models of Chikungunya Virus Infection and Disease (CHIKV NHP Model). Pathogens 2015, 4, 662–681. [Google Scholar] [CrossRef] [PubMed]
- Fros, J.J.; Liu, W.J.; Prow, N.A.; Geertsema, C.; Ligtenberg, M.; Vanlandingham, D.L.; Schnettler, E.; Vlak, J.M.; Suhrbier, A.; Khromykh, A.A.; et al. Chikungunya Virus Nonstructural Protein 2 Inhibits Type I/II Interferon-Stimulated JAK-STAT Signaling. J. Virol. 2010, 84, 10877–10887. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.H.; Maric, M.; Madison, M.N.; Maury, W.; Roller, R.J.; Okeoma, C.M. BST-2/tetherin-mediated restriction of chikungunya (CHIKV) VLP budding is counteracted by CHIKV non-structural protein 1 (nsP1). Virology 2013, 438, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Colavita, F.; Musumeci, G.; Caglioti, C. Human osteoblast-like cells are permissive for Zika virus replication. J. Rheumatol. 2018, 45, 443. [Google Scholar] [CrossRef] [PubMed]
- Kvansakul, M. Viral Infection and Apoptosis. Viruses 2017, 9, 356. [Google Scholar] [CrossRef] [PubMed]
- Krejbich-Trotot, P.; Denizot, M.; Hoarau, J.-J.; Jaffar-Bandjee, M.-C.; Das, T.; Gasque, P. Chikungunya virus mobilizes the apoptotic machinery to invade host cell defenses. FASEB J. 2011, 25, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Chow, A.; Her, Z.; Ong, E.K.S.; Chen, J.M.; Dimatatac, F.; Kwek, D.J.C.; Barkham, T.; Yang, H.; Rénia, L.; Leo, Y.S.; et al. Persistent arthralgia induced by Chikungunya virus infection is associated with interleukin-6 and granulocyte macrophage colony-stimulating factor. J. Infect. Dis. 2011, 203, 149–157. [Google Scholar] [CrossRef]
- Kelvin, A.A.; Banner, D.; Silvi, G.; Moro, M.L.; Spataro, N.; Gaibani, P.; Cavrini, F.; Pierro, A.; Rossini, G.; Cameron, M.J.; et al. Inflammatory Cytokine Expression Is Associated with Chikungunya Virus Resolution and Symptom Severity. PLoS Negl. Trop. Dis. 2011, 5, e1279. [Google Scholar] [CrossRef]
- Michlmayr, D.; Pak, T.R.; Rahman, A.H.; Amir, E.D.; Kim, E.; Kim-Schulze, S.; Suprun, M.; Stewart, M.G.; Thomas, G.P.; Balmaseda, A.; et al. Comprehensive innate immune profiling of chikungunya virus infection in pediatric cases. Mol. Syst. Biol. 2018, 14, e7862. [Google Scholar] [CrossRef]
- Poo, Y.S.; Rudd, P.A.; Gardner, J.; Wilson, J.A.C.; Larcher, T.; Colle, M.A.; Le, T.T.; Nakaya, H.I.; Warrilow, D.; Allcock, R.; et al. Multiple Immune Factors Are Involved in Controlling Acute and Chronic Chikungunya Virus Infection. PLoS Negl. Trop. Dis. 2014, 8, e3354. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.; Mani, R.S.; Desai, A.; Ravi, V. Correlation of plasma viral loads and presence of Chikungunya IgM antibodies with cytokine/chemokine levels during acute Chikungunya virus infection. J. Med. Virol. 2014, 86, 1393–1401. [Google Scholar] [CrossRef] [PubMed]
- Venugopalan, A.; Ghorpade, R.P.; Chopra, A. Cytokines in Acute Chikungunya. PLoS ONE 2014, 9, e111305. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.F.P.; Chow, A.; Sun, Y.-J.; Kwek, D.J.C.; Lim, P.-L.; Dimatatac, F.; Ng, L.-C.; Ooi, E.-E.; Choo, K.-H.; Her, Z.; et al. IL-1β, IL-6, and RANTES as Biomarkers of Chikungunya Severity. PLoS ONE 2009, 4, e4261. [Google Scholar] [CrossRef] [PubMed]
- Hoarau, J.J.; Jaffar Bandjee, M.C.; Krejbich Trotot, P.; Das, T.; Li-Pat-Yuen, G.; Dassa, B.; Denizot, M.; Guichard, E.; Ribera, A.; Henni, T.; et al. Persistent Chronic Inflammation and Infection by Chikungunya Arthritogenic Alphavirus in Spite of a Robust Host Immune Response. J. Immunol. 2010, 184, 5914–5927. [Google Scholar] [CrossRef]
- Teng, T.-S.; Kam, Y.-W.; Lee, B.; Hapuarachchi, H.C.; Wimal, A.; Ng, L.-C.; Ng, L.F.P. A Systematic Meta-analysis of Immune Signatures in Patients With Acute Chikungunya Virus Infection. J. Infect. Dis. 2015, 211, 1925–1935. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.; Poddar, S.; Shimak, R.M.; Diamond, M.S. Interferon Regulatory Factor 1 Protects against Chikungunya Virus-Induced Immunopathology by Restricting Infection in Muscle Cells. J. Virol. 2017, 91. [Google Scholar] [CrossRef]
- Pruetz, J.D.; Socha, A.; Kante, D. New Range Record for the Lesser Spot-nosed Guenon (Cercopithecus petaurista) in Southeastern Senegal. Am. J. Primatol. 2010, 7, 64–66. [Google Scholar]
- Lum, F.-M.; Ng, L.F.P. Cellular and molecular mechanisms of chikungunya pathogenesis. Antiviral Res. 2015, 120, 165–174. [Google Scholar] [CrossRef]
- Hawman, D.W.; Stoermer, K.A.; Montgomery, S.A.; Pal, P.; Oko, L.; Diamond, M.S.; Morrison, T.E. Chronic Joint Disease Caused by Persistent Chikungunya Virus Infection Is Controlled by the Adaptive Immune Response. J. Virol. 2013, 87, 13878–13888. [Google Scholar] [CrossRef]
- Teo, T.-H.; Lum, F.-M.; Claser, C.; Lulla, V.; Lulla, A.; Merits, A.; Renia, L.; Ng, L.F.P. A Pathogenic Role for CD4+ T Cells during Chikungunya Virus Infection in Mice. J. Immunol. 2013, 190, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Partidos, C.D.; Weger, J.; Brewoo, J.; Seymour, R.; Borland, E.M.; Ledermann, J.P.; Powers, A.M.; Weaver, S.C.; Stinchcomb, D.T.; Osorio, J.E. Probing the attenuation and protective efficacy of a candidate chikungunya virus vaccine in mice with compromised interferon (IFN) signaling. Vaccine 2011, 29, 3067–3073. [Google Scholar] [CrossRef] [PubMed]
- Mallilankaraman, K.; Shedlock, D.J.; Bao, H.; Kawalekar, O.U.; Fagone, P.; Ramanathan, A.A.; Ferraro, B.; Stabenow, J.; Vijayachari, P.; Sundaram, S.G.; et al. A DNA Vaccine against Chikungunya Virus Is Protective in Mice and Induces Neutralizing Antibodies in Mice and Nonhuman Primates. PLoS Negl. Trop. Dis. 2011, 5, e928. [Google Scholar] [CrossRef]
- Messaoudi, I.; Vomaske, J.; Totonchy, T.; Kreklywich, C.N.; Haberthur, K.; Springgay, L.; Brien, J.D.; Diamond, M.S.; DeFilippis, V.R.; Streblow, D.N. Chikungunya Virus Infection Results in Higher and Persistent Viral Replication in Aged Rhesus Macaques Due to Defects in Anti-Viral Immunity. PLoS Negl. Trop. Dis. 2013, 7, e2343. [Google Scholar] [CrossRef] [PubMed]
- Ekchariyawat, P.; Hamel, R.; Bernard, E.; Wichit, S.; Surasombatpattana, P.; Talignani, L.; Thomas, F.; Choumet, V.; Yssel, H.; Desprès, P.; et al. Inflammasome signaling pathways exert antiviral effect against Chikungunya virus in human dermal fibroblasts. Infect. Genet. Evol. 2015, 32, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Sourisseau, M.; Schilte, C.; Casartelli, N.; Trouillet, C.; Guivel-Benhassine, F.; Rudnicka, D.; Sol-Foulon, N.; Le Roux, K.; Prevost, M.C.; Fsihi, H.; et al. Characterization of reemerging chikungunya virus. PLoS Pathog. 2007, 3, 0804–0817. [Google Scholar] [CrossRef] [PubMed]
- Wichit, S.; Diop, F.; Hamel, R.; Talignani, L.; Ferraris, P.; Cornelie, S.; Liegeois, F.; Thomas, F.; Yssel, H.; Missé, D. Aedes Aegypti saliva enhances chikungunya virus replication in human skin fibroblasts via inhibition of the type I interferon signaling pathway. Infect. Genet. Evol. 2017, 55, 68–70. [Google Scholar] [CrossRef]
- Couderc, T.; Chrétien, F.; Schilte, C.; Disson, O.; Brigitte, M.; Guivel-Benhassine, F.; Touret, Y.; Barau, G.; Cayet, N.; Schuffenecker, I.; et al. A mouse model for Chikungunya: Young age and inefficient type-I interferon signaling are risk factors for severe disease. PLoS Pathog. 2008, 4, e29. [Google Scholar] [CrossRef]
- Puiprom, O.; Morales Vargas, R.E.; Potiwat, R.; Chaichana, P.; Ikuta, K.; Ramasoota, P.; Okabayashi, T. Characterization of chikungunya virus infection of a human keratinocyte cell line: Role of mosquito salivary gland protein in suppressing the host immune response. Infect. Genet. Evol. 2013, 17, 210–215. [Google Scholar] [CrossRef]
- Bernard, E.; Hamel, R.; Neyret, A.; Ekchariyawat, P.; Molès, J.P.; Simmons, G.; Chazal, N.; Desprès, P.; Missé, D.; Briant, L. Human keratinocytes restrict chikungunya virus replication at a post-fusion step. Virology 2015, 476, 1–10. [Google Scholar] [CrossRef]
- Rudd, P.A.; Wilson, J.; Gardner, J.; Larcher, T.; Babarit, C.; Le, T.T.; Anraku, I.; Kumagai, Y.; Loo, Y.-M.; Gale, M.; et al. Interferon Response Factors 3 and 7 Protect against Chikungunya Virus Hemorrhagic Fever and Shock. J. Virol. 2012, 86, 9888–9898. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Yanai, H.; Negishi, H.; Asagiri, M.; Sato, M.; Mizutani, T.; Shimada, N.; Ohba, Y.; Takaoka, A.; Yoshida, N.; et al. IRF-7 is the master regulator of type-I interferon-dependent immune responses. Nature 2005, 434, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Osterlund, P.I.; Pietila, T.E.; Veckman, V.; Kotenko, S.V.; Julkunen, I. IFN Regulatory Factor Family Members Differentially Regulate the Expression of Type III IFN (IFN-) Genes. J. Immunol. 2007, 179, 3434–3442. [Google Scholar] [CrossRef] [PubMed]
- Gasque, P.; Jaffar-Bandjee, M.C. The immunology and inflammatory responses of human melanocytes in infectious diseases. J. Infect. 2015, 71, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Parola, P.; de Lamballerie, X.; Jourdan, J.; Rovery, C.; Vaillant, V.; Minodier, P.; Brouqui, P.; Flahault, A.; Raoult, D.; Charrel, R.N. Novel chikungunya virus variant in travelers returning from Indian Ocean islands. Emerg. Infect. Dis. 2006, 12, 1493–1499. [Google Scholar] [CrossRef] [PubMed]
- Her, Z.; Malleret, B.; Chan, M.; Ong, E.K.S.; Wong, S.C.; Kwek, D.J.C.; Tolou, H.; Lin, R.T.P.; Tambyah, P.A.; Renia, L.; et al. Active Infection of Human Blood Monocytes by Chikungunya Virus Triggers an Innate Immune Response. J. Immunol. 2010, 184, 5903–5913. [Google Scholar] [CrossRef] [PubMed]
- Ruiz Silva, M.; Van Der Ende-Metselaar, H.; Mulder, H.L.; Smit, J.M.; Rodenhuis-Zybert, I.A. Mechanism and role of MCP-1 upregulation upon chikungunya virus infection in human peripheral blood mononuclear cells. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef]
- Ruiz Silva, M.; Aguilar Briseño, J.A.; Upasani, V.; van der Ende-Metselaar, H.; Smit, J.M.; Rodenhuis-Zybert, I.A. Suppression of chikungunya virus replication and differential innate responses of human peripheral blood mononuclear cells during co-infection with dengue virus. PLoS Negl. Trop. Dis. 2017, 11, e0005712. [Google Scholar] [CrossRef]
- Laughhunn, A.; Huang, Y.-J.S.; Vanlandingham, D.L.; Lanteri, M.C.; Stassinopoulos, A. Inactivation of chikungunya virus in blood components treated with amotosalen/ultraviolet A light or amustaline/glutathione. Transfusion 2018, 58, 748–757. [Google Scholar] [CrossRef]
- Tsetsarkin, K.A.; Sampson-Johannes, A.; Sawyer, L.; Kinsey, J.; Higgs, S.; Vanlandingham, D.L. Photochemical Inactivation of Chikungunya Virus in Human Apheresis Platelet Components by Amotosalen and UVA Light. Am. J. Trop. Med. Hyg. 2013, 88, 1163–1169. [Google Scholar] [CrossRef]
- Ozden, S.; Huerre, M.; Riviere, J.P.; Coffey, L.L.; Afonso, P.V.; Mouly, V.; de Monredon, J.; Roger, J.C.; El Amrani, M.; Yvin, J.L.; et al. Human Muscle Satellite Cells as Targets of Chikungunya Virus Infection. PLoS ONE 2007, 2, e527. [Google Scholar] [CrossRef] [PubMed]
- Lohachanakul, J.; Phuklia, W.; Thannagith, M.; Thongsakulprasert, T.; Smith, D.R.; Ubol, S. Differences in response of primary human myoblasts to infection with recent epidemic strains of Chikungunya virus isolated from patients with and without myalgia. J. Med. Virol. 2015, 87, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Deeba, F.; Islam, A.; Kazim, S.N.; Naqvi, I.H.; Broor, S.; Ahmed, A.; Parveen, S. Chikungunya virus: Recent advances in epidemiology, host pathogen interaction and vaccine strategies. Pathog. Dis. 2016, 74, ftv119. [Google Scholar] [CrossRef] [PubMed]
- Manimunda, S.P.; Singh, S.S.; Sugunan, A.P.; Singh, O.; Roy, S.; Shriram, A.N.; Bharadwaj, A.P.; Shah, W.A.; Vijayachari, P. Chikungunya Fever, Andaman and Nicobar Islands, India. Emerg. Infect. Dis. 2007, 13, 1259–1260. [Google Scholar] [CrossRef] [PubMed]
- Phuklia, W.; Kasisith, J.; Modhiran, N.; Rodpai, E.; Thannagith, M.; Thongsakulprasert, T.; Smith, D.R.; Ubol, S. Osteoclastogenesis induced by CHIKV-infected fibroblast-like synoviocytes: A possible interplay between synoviocytes and monocytes/macrophages in CHIKV-induced arthralgia/arthritis. Virus Res. 2013, 177, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Lokireddy, S.; Vemula, S.; Vadde, R. Connective tissue metabolism in chikungunya patients. Virol. J. 2008, 5, 31. [Google Scholar] [CrossRef] [PubMed]
- Herrero, L.J.; Foo, S.-S.; Sheng, K.-C.; Chen, W.; Forwood, M.R.; Bucala, R.; Mahalingam, S. Pentosan Polysulfate: A Novel Glycosaminoglycan-Like Molecule for Effective Treatment of Alphavirus-Induced Cartilage Destruction and Inflammatory Disease. J. Virol. 2015, 89, 8063–8076. [Google Scholar] [CrossRef]
- Kuo, S.C.; Wang, Y.M.; Ho, Y.J.; Chang, T.Y.; Lai, Z.Z.; Tsui, P.Y.; Wu, T.Y.; Lin, C.C. Suramin treatment reduces chikungunya pathogenesis in mice. Antiviral Res. 2016, 134, 89–96. [Google Scholar] [CrossRef]
- Noret, M.; Herrero, L.; Rulli, N.; Rolph, M.; Smith, P.N.; Li, R.W.; Roques, P.; Gras, G.; Mahalingam, S. Interleukin 6, RANKL, and osteoprotegerin expression by chikungunya virus-infected human osteoblasts. J. Infect. Dis. 2012, 206, 455–457. [Google Scholar] [CrossRef]
- Goupil, B.A.; McNulty, M.A.; Martin, M.J.; McCracken, M.K.; Christofferson, R.C.; Mores, C.N. Novel Lesions of Bones and Joints Associated with Chikungunya Virus Infection in Two Mouse Models of Disease: New Insights into Disease Pathogenesis. PLoS ONE 2016, 11, e0155243. [Google Scholar] [CrossRef]
- Chen, W.; Foo, S.; Taylor, A.; Lulla, A.; Merits, A.; Hueston, L.; Forwood, M.R.; Walsh, N.C.; Sims, N.A.; Herrero, L.J.; et al. Bindarit, an Inhibitor of Monocyte Chemotactic Protein Synthesis, Protects against Bone Loss Induced by Chikungunya Virus Infection. J. Virol. 2015, 89, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Labadie, K.; Larcher, T.; Joubert, C.; Mannioui, A.; Delache, B.; Brochard, P.; Guigand, L.; Dubreil, L.; Lebon, P.; Verrier, B.; et al. Chikungunya disease in nonhuman primates involves long-term viral persistence in macrophages. J. Clin. Investig. 2010, 120, 894–906. [Google Scholar] [CrossRef] [PubMed]
- Gardner, J.; Anraku, I.; Le, T.T.; Larcher, T.; Major, L.; Roques, P.; Schroder, W.A.; Higgs, S.; Suhrbier, A. Chikungunya Virus Arthritis in Adult Wild-Type Mice. J. Virol. 2010, 84, 8021–8032. [Google Scholar] [CrossRef] [PubMed]
- Mazaud, R.; Salaün, J.J.; Montabone, H.; Goube, P.; Bazillio, R. Acute neurologic and sensorial disorders in dengue and Chikungunya fever. Bull. Soc. Pathol. Exot. Filiales 1971, 64, 22–30. [Google Scholar] [PubMed]
- Arpino, C.; Curatolo, P.; Rezza, G. Chikungunya and the nervous system: What we do and do not know. Rev. Med. Virol. 2009, 19, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Thiruvengadam, K.V.; Kalyanasundaram, V.; Rajgopal, J. Clinical and pathological studies on chikungunya fever in Madras city. Indian J. Med. Res. 1965, 53, 729–744. [Google Scholar] [PubMed]
- Ganesan, K.; Diwan, A.; Shankar, S.K.; Desai, S.B.; Sainani, G.S.; Katrak, S.M. Chikungunya encephalomyeloradiculitis: Report of 2 cases with neuroimaging and 1 case with autopsy findings. Am. J. Neuroradiol. 2008, 29, 1636–1637. [Google Scholar] [CrossRef] [PubMed]
- Dhanwani, R.; Khan, M.; Bhaskar, A.S.B.; Singh, R.; Patro, I.K.; Rao, P.V.L.; Parida, M.M. Characterization of Chikungunya virus infection in human neuroblastoma SH-SY5Y cells: Role of apoptosis in neuronal cell death. Virus Res. 2012, 163, 563–572. [Google Scholar] [CrossRef]
- Solignat, M.; Gay, B.; Higgs, S.; Briant, L.; Devaux, C. Replication cycle of chikungunya: A re-emerging arbovirus. Virology 2009, 393, 183–197. [Google Scholar] [CrossRef]
- Abraham, R.; Mudaliar, P.; Padmanabhan, A.; Sreekumar, E. Induction of Cytopathogenicity in Human Glioblastoma Cells by Chikungunya Virus. PLoS ONE 2013, 8, e75854. [Google Scholar] [CrossRef]
- Abraham, R.; Singh, S.; Nair, S.R.; Hulyalkar, N.V.; Surendran, A.; Jaleel, A.; Sreekumar, E. Nucleophosmin (NPM1)/B23 in the Proteome of Human Astrocytic Cells Restricts Chikungunya Virus Replication. J. Proteome Res. 2017, 16, 4144–4155. [Google Scholar] [CrossRef] [PubMed]
- Abere, B.; Wikan, N.; Ubol, S.; Auewarakul, P.; Paemanee, A.; Kittisenachai, S.; Roytrakul, S.; Smith, D.R. Proteomic analysis of chikungunya virus infected microgial cells. PLoS ONE 2012, 7, e34800. [Google Scholar] [CrossRef] [PubMed]
- Wikan, N.; Sakoonwatanyoo, P.; Ubol, S.; Yoksan, S.; Smith, D.R. Chikungunya virus infection of cell lines: Analysis of the east, central and south African lineage. PLoS ONE 2012, 7, e31102. [Google Scholar] [CrossRef] [PubMed]
- Chiam, C.W.; Chan, Y.F.; Ong, K.C.; Wong, K.T.; Sam, I.C. Neurovirulence comparison of chikungunya virus isolates of the Asian and East/Central/South African genotypes from Malaysia. J. Gen. Virol. 2015, 96, 3243–3254. [Google Scholar] [CrossRef] [PubMed]
- Das, T.; Hoarau, J.J.; Bandjee, M.C.J.; Maquart, M.; Gasque, P. Multifaceted innate immune responses engaged by astrocytes, microglia and resident dendritic cells against Chikungunya neuroinfection. J. Gen. Virol. 2015, 96, 294–310. [Google Scholar] [CrossRef] [PubMed]
- Fraisier, C.; Koraka, P.; Belghazi, M.; Bakli, M.; Granjeaud, S.; Pophillat, M.; Lim, S.M.; Osterhaus, A.; Martina, B.; Camoin, L.; et al. Kinetic Analysis of Mouse Brain Proteome Alterations Following Chikungunya Virus Infection before and after Appearance of Clinical Symptoms. PLoS ONE 2014, 9, e91397. [Google Scholar] [CrossRef] [PubMed]
- Lim, P.J.; Chu, J.J.H. A Polarized Cell Model for Chikungunya Virus Infection: Entry and Egress of Virus Occurs at the Apical Domain of Polarized Cells. PLoS Negl. Trop. Dis. 2014, 8, e2661. [Google Scholar] [CrossRef] [PubMed]
- Passoni, G.; Langevin, C.; Palha, N.; Mounce, B.C.; Briolat, V.; Affaticati, P.; De Job, E.; Joly, J.-S.; Vignuzzi, M.; Saleh, M.; et al. Imaging of viral neuroinvasion in the zebrafish reveals that Sindbis and chikungunya viruses favour different entry routes. Dis. Model. Mech. 2017, 10, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Mahendradas, P.; Avadhani, K.; Shetty, R. Chikungunya and the eye: A review. J. Ophthalmic Inflamm. Infect. 2013, 3, 1–9. [Google Scholar] [CrossRef]
- Lin, J.; Chen, R.W.S.; Hazan, A.; Weiss, M. Chikungunya Virus Infection Manifesting as Intermediate Uveitis. Ocul. Immunol. Inflamm. 2018, 26, 680–682. [Google Scholar] [CrossRef]
- Salceanu, S.O.; Raman, V. Recurrent chikungunya retinitis. BMJ Case Rep. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Ulloa-Padilla, J.P.; Davila, P.J.; Izquierdo, N.J.; Garcia-Rodriguez, O.; Jimenez, I.Z. Ocular Symptoms and Signs of Chikungunya Fever in Puerto Rico. P. R. Health Sci. J. 2018, 37, 83–87. [Google Scholar] [PubMed]
- Mahendradas, P.; Shetty, R.; Malathi, J.; Madhavan, H.N. Chikungunya virus iridocyclitis in Fuchs’ heterochromic iridocyclitis. Indian J. Ophthalmol. 2010, 58, 545–547. [Google Scholar] [CrossRef] [PubMed]
- Babu, K.; Murthy, G. Chikungunya virus iridocyclitis in Fuchs′ heterochromic iridocyclitis. Indian J. Ophthalmol. 2012, 60, 73. [Google Scholar] [CrossRef] [PubMed]
- Couderc, T.; Gangneux, N.; Chrétien, F.; Caro, V.; Le Luong, T.; Ducloux, B.; Tolou, H.; Lecuit, M.; Grandadam, M. Chikungunya virus infection of corneal grafts. J. Infect. Dis. 2012, 206, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Pal, P.; Fox, J.M.; Hawman, D.W.; Huang, Y.-J.S.; Messaoudi, I.; Kreklywich, C.; Denton, M.; Legasse, A.W.; Smith, P.P.; Johnson, S.; et al. Chikungunya Viruses That Escape Monoclonal Antibody Therapy Are Clinically Attenuated, Stable, and Not Purified in Mosquitoes. J. Virol. 2014, 88, 8213–8226. [Google Scholar] [CrossRef] [PubMed]
- Chua, H.H.; Rashid, K.A.; Law, W.C.; Hamizah, A.; Chem, Y.K.; Khairul, A.H.; Chua, K.B. A fatal case of chikungunya virus infection with liver involvement. Med. J. Malaysia 2010, 65, 83–84. [Google Scholar]
- Davis, J.L.; Hodge, H.M.; Campbell, W.E. Growth of chikungunya virus in baby hamster kidney cell (BHK-21-clone 13) suspension cultures. Appl. Microbiol. 1971, 21, 338–341. [Google Scholar]
- Gardner, J.; Rudd, P.A.; Prow, N.A.; Belarbi, E.; Roques, P.; Larcher, T.; Gresh, L.; Balmaseda, A.; Harris, E.; Schroder, W.A.; et al. Infectious chikungunya virus in the saliva of mice, monkeys and humans. PLoS ONE 2015, 10, 1–15. [Google Scholar] [CrossRef]
- Niedrig, M.; Patel, P.; El Wahed, A.A.; Schädler, R.; Yactayo, S. Find the right sample: A study on the versatility of saliva and urine samples for the diagnosis of emerging viruses. BMC Infect. Dis. 2018, 18, 707. [Google Scholar] [CrossRef]
- Musso, D.; Teissier, A.; Rouault, E.; Teururai, S.; de Pina, J.-J.; Nhan, T.-X. Detection of chikungunya virus in saliva and urine. Virol. J. 2016, 13, 102. [Google Scholar] [CrossRef] [PubMed]
- Contopoulos-Ioannidis, D.; Newman-Lindsay, S.; Chow, C.; LaBeaud, A.D. Mother-to-child transmission of Chikungunya virus: A systematic review and meta-analysis. PLoS Negl. Trop. Dis. 2018, 12, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Gérardin, P.; Barau, G.; Michault, A.; Bintner, M.; Randrianaivo, H.; Choker, G.; Lenglet, Y.; Touret, Y.; Bouveret, A.; Grivard, P.; et al. Multidisciplinary Prospective Study of Mother-to-Child Chikungunya Virus Infections on the Island of La Réunion. PLoS Med. 2008, 5, e60. [Google Scholar] [CrossRef] [PubMed]
- Touret, Y.; Randrianaivo, H.; Michault, A.; Schuffenecker, I.; Kauffmann, E.; Lenglet, Y.; Barau, G.; Fourmaintraux, A. Early maternal-fetal transmission of the Chikungunya virus. Presse Med. 2006, 35, 1656–1658. [Google Scholar] [CrossRef]
- Grivard, P.; Le Roux, K.; Laurent, P.; Fianu, A.; Perrau, J.; Gigan, J.; Hoarau, G.; Grondin, N.; Staikowsky, F.; Favier, F.; et al. Molecular and serological diagnosis of Chikungunya virus infection. Pathol. Biol. 2007, 55, 490–494. [Google Scholar] [CrossRef] [PubMed]
- Prata-Barbosa, A.; Cleto-Yamane, T.L.; Robaina, J.R.; Guastavino, A.B.; de Magalhães-Barbosa, M.C.; Brindeiro, R.D.M.; Medronho, R.A.; da Cunha, A.J.L.A. Co-infection with Zika and Chikungunya viruses associated with fetal death—A case report. Int. J. Infect. Dis. 2018, 72, 25–27. [Google Scholar] [CrossRef] [PubMed]
- Charlier, C.; Beaudoin, M.-C.; Couderc, T.; Lortholary, O.; Lecuit, M. Arboviruses and pregnancy: Maternal, fetal, and neonatal effects. Lancet Child Adolesc. Heal. 2017, 1, 134–146. [Google Scholar] [CrossRef]
- Platt, D.J.; Smith, A.M.; Arora, N.; Diamond, M.S.; Coyne, C.B.; Miner, J.J. Zika virus–related neurotropic flaviviruses infect human placental explants and cause fetal demise in mice. Sci. Transl. Med. 2018, 10, eaao7090. [Google Scholar] [CrossRef]
- Bandeira, A.C.; Campos, G.S.; Rocha, V.F.D.; Souza, B.S.D.F.; Soares, M.B.P.; Oliveira, A.A.; de Abreu, Y.C.; Menezes, G.S.; Sardi, S.I. Prolonged shedding of Chikungunya virus in semen and urine: A new perspective for diagnosis and implications for transmission. IDCases 2016, 6, 100–103. [Google Scholar] [CrossRef]
- Hua, C.; Combe, B. Chikungunya Virus-Associated Disease. Curr. Rheumatol. Rep. 2017, 19, 69. [Google Scholar] [CrossRef]
- Vogels, C.B.F.; Rückert, C.; Cavany, S.M.; Perkins, T.A.; Ebel, G.D.; Grubaugh, N.D. Arbovirus coinfection and co-transmission: A neglected public health concern? PLOS Biol. 2019, 17, e3000130. [Google Scholar] [CrossRef] [PubMed]
- WHO Guidelines on Clinical Management of Chikungunya Fever. Available online: http://www.who.int/iris/handle/10665/205178 (accessed on 19 February 2019).
- Silva, J.V.J.; Ludwig-Begall, L.F.; Oliveira-Filho, E.F.D.; Oliveira, R.A.S.; Durães-Carvalho, R.; Lopes, T.R.R.; Silva, D.E.A.; Gil, L.H.V.G. A scoping review of Chikungunya virus infection: Epidemiology, clinical characteristics, viral co-circulation complications, and control. Acta Trop. 2018, 188, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.A.; Dermody, T.S. Chikungunya virus: Epidemiology, replication, disease mechanisms, and prospective intervention strategies. J. Clin. Investig. 2017, 127, 737–749. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.C.; Reis, P.A.; de Freitas, C.S.; Sacramento, C.Q.; Villas Bôas Hoelz, L.; Bastos, M.M.; Mattos, M.; Rocha, N.; Gomes de Azevedo Quintanilha, I.; da Silva Gouveia Pedrosa, C.; et al. Beyond Members of the Flaviviridae Family, Sofosbuvir Also Inhibits Chikungunya Virus Replication. Antimicrob. Agents Chemother. 2018, 63, e01389-18. [Google Scholar] [CrossRef] [PubMed]
- Franco, E.J.; Rodriquez, J.L.; Pomeroy, J.J.; Hanrahan, K.C.; Brown, A.N. The effectiveness of antiviral agents with broad-spectrum activity against chikungunya virus varies between host cell lines. Antivir. Chem. Chemother. 2018, 26, 204020661880758. [Google Scholar] [CrossRef] [PubMed]
- Chopra, A.; Saluja, M.; Venugopalan, A. Effectiveness of Chloroquine and Inflammatory Cytokine Response in Patients With Early Persistent Musculoskeletal Pain and Arthritis Following Chikungunya Virus Infection. Arthritis Rheumatol. 2014, 66, 319–326. [Google Scholar] [CrossRef] [PubMed]
- de Lamballerie, X.; Ninove, L.; Charrel, R.N. Antiviral treatment of chikungunya virus infection. Infect. Disord. Drug Targets 2009, 9, 101–104. [Google Scholar] [CrossRef]
- Khan, M.; Santhosh, S.R.; Tiwari, M.; Lakshmana Rao, P.V.; Parida, M. Assessment of in vitro prophylactic and therapeutic efficacy of chloroquine against chikungunya virus in vero cells. J. Med. Virol. 2010, 82, 817–824. [Google Scholar] [CrossRef]
- Roques, P.; Thiberville, S.D.; Dupuis-Maguiraga, L.; Lum, F.M.; Labadie, K.; Martinon, F.; Gras, G.; Lebon, P.; Ng, L.F.P.; de Lamballerie, X.; et al. Paradoxical effect of chloroquine treatment in enhancing chikungunya virus infection. Viruses 2018, 10, 268. [Google Scholar] [CrossRef]
- Smith, S.A.; Silva, L.A.; Fox, J.M.; Flyak, A.I.; Kose, N.; Sapparapu, G.; Khomandiak, S.; Ashbrook, A.W.; Kahle, K.M.; Fong, R.H.; et al. Isolation and Characterization of Broad and Ultrapotent Human Monoclonal Antibodies with Therapeutic Activity against Chikungunya Virus. Cell Host Microbe 2015, 18, 86–95. [Google Scholar] [CrossRef]
- Reyes-Sandoval, A. 51 years in of Chikungunya clinical vaccine development: A historical perspective. Hum. Vaccin. Immunother. 2019. [Google Scholar] [CrossRef] [PubMed]
- Plante, K.; Wang, E.; Partidos, C.D.; Weger, J.; Gorchakov, R.; Tsetsarkin, K.; Borland, E.M.; Powers, A.M.; Seymour, R.; Stinchcomb, D.T.; et al. Novel Chikungunya Vaccine Candidate with an IRES-Based Attenuation and Host Range Alteration Mechanism. PLoS Pathog. 2011, 7, e1002142. [Google Scholar] [CrossRef]
- Wang, E.; Kim, D.Y.; Weaver, S.C.; Frolov, I. Chimeric Chikungunya Viruses Are Nonpathogenic in Highly Sensitive Mouse Models but Efficiently Induce a Protective Immune Response. J. Virol. 2011, 85, 9249–9252. [Google Scholar] [CrossRef] [PubMed]
- Nougairede, A.; De Fabritus, L.; Aubry, F.; Gould, E.A.; Holmes, E.C.; de Lamballerie, X. Random Codon Re-encoding Induces Stable Reduction of Replicative Fitness of Chikungunya Virus in Primate and Mosquito Cells. PLoS Pathog. 2013, 9, e1003172. [Google Scholar] [CrossRef] [PubMed]
- Achee, N.L.; Grieco, J.P.; Vatandoost, H.; Seixas, G.; Pinto, J.; Ching-NG, L.; Martins, A.J.; Juntarajumnong, W.; Corbel, V.; Gouagna, C.; et al. Alternative strategies for mosquito-borne arbovirus control. PLoS Negl. Trop. Dis. 2019, 13, e0006822. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| World Region | Species | Common Name | Method of Detection | References |
|---|---|---|---|---|
| NHPs: | ||||
| Africa | Cercopithecus aethiops | Vervet monkey | Isolation and specific antibodies | [27,28,31] |
| Cercopithecus mitis | Blue monkey | Specific antibodies | [67] | |
| Cercopithecus ascanius | Red tailed monkey | Specific antibodies | [67,71] | |
| Galago senegalensis | Senegal bushbaby | Isolation and specific antibodies | [27,31] | |
| Papio papio | Guinea baboon | Isolation and specific antibodies | [27,66] | |
| Papio cynocephalus | Yellow baboon | Specific antibodies | [67] | |
| Papio ursinus | Cape baboon | Specific antibodies | [28,82] | |
| Erythrocebus patas | Patas monkey | Isolation and specific antibodies | [27,66] | |
| Chlorocebus sabaeus | African green monkey | Isolation and specific antibodies | [27,66] | |
| Mandrillus sphinx | Mandrill | Specific antibodies | [70] | |
| Indian Ocean | Eulemur fulvus | Brown lemur | Specific antibodies | [69] |
| Indian Ocean and Asia | Macaca fascicularis | Crab eating macaque | Isolation and specific antibodies | [74,77,78] |
| Asia | Macaca nemestrina | Pig tailed macaque | Specific antibodies | [75] |
| South America | Ateles marginatus | Spider monkey | Specific antibodies | [79] |
| Callithrix jacchus | Common marmoset | Specific antibodies | [79] | |
| Sapajus xanthosternos | Golden-bellied capuchin | Specific antibodies | [79] | |
| Sapajus robustu | Crested capuchin | Specific antibodies | [79] | |
| Sapajus flavius | Capuchin monkey | Specific antibodies | [79] | |
| Mammals: | ||||
| Africa | Xerus erythropus | Palm squirrel | Isolation | [31] |
| Rattus rattus | Ship rat | Specific antibodies | [69] | |
| Scotophillus species | Bat | Isolation | [31] | |
| Syncerus caffer nanus | Buffalo | Specific antibodies | [70] | |
| Loxodonta africana | Elephant | Specific antibodies | [70] | |
| Specimen | Tested | Positive |
|---|---|---|
| Saliva | 5 | 1 |
| Urine | 9 | 1 |
| Ocular swabs | 2 | 0 |
| CVF | 4 | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matusali, G.; Colavita, F.; Bordi, L.; Lalle, E.; Ippolito, G.; Capobianchi, M.R.; Castilletti, C. Tropism of the Chikungunya Virus. Viruses 2019, 11, 175. https://doi.org/10.3390/v11020175
Matusali G, Colavita F, Bordi L, Lalle E, Ippolito G, Capobianchi MR, Castilletti C. Tropism of the Chikungunya Virus. Viruses. 2019; 11(2):175. https://doi.org/10.3390/v11020175
Chicago/Turabian StyleMatusali, Giulia, Francesca Colavita, Licia Bordi, Eleonora Lalle, Giuseppe Ippolito, Maria R. Capobianchi, and Concetta Castilletti. 2019. "Tropism of the Chikungunya Virus" Viruses 11, no. 2: 175. https://doi.org/10.3390/v11020175
APA StyleMatusali, G., Colavita, F., Bordi, L., Lalle, E., Ippolito, G., Capobianchi, M. R., & Castilletti, C. (2019). Tropism of the Chikungunya Virus. Viruses, 11(2), 175. https://doi.org/10.3390/v11020175