Camelids and Cattle Are Dead-End Hosts for Peste-des-Petits-Ruminants Virus

 ,
,  , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Virus
2.2. Animals and Study Design
2.3. Serological and Hematological Analyses
2.4. Virological Analyses
2.5. Pathological Analyses
2.6. Comparison of Performance Characteristics of Different Virological Methods
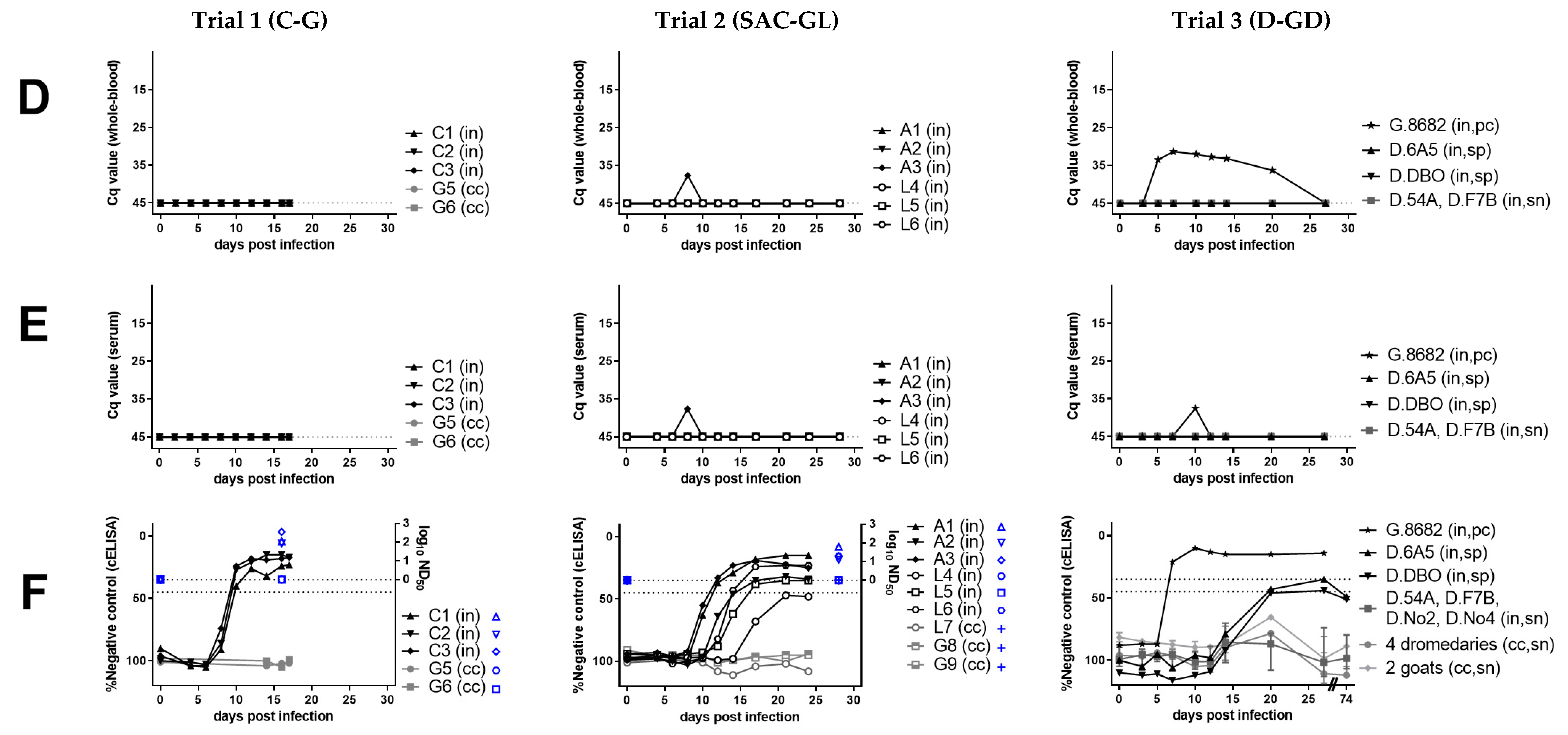
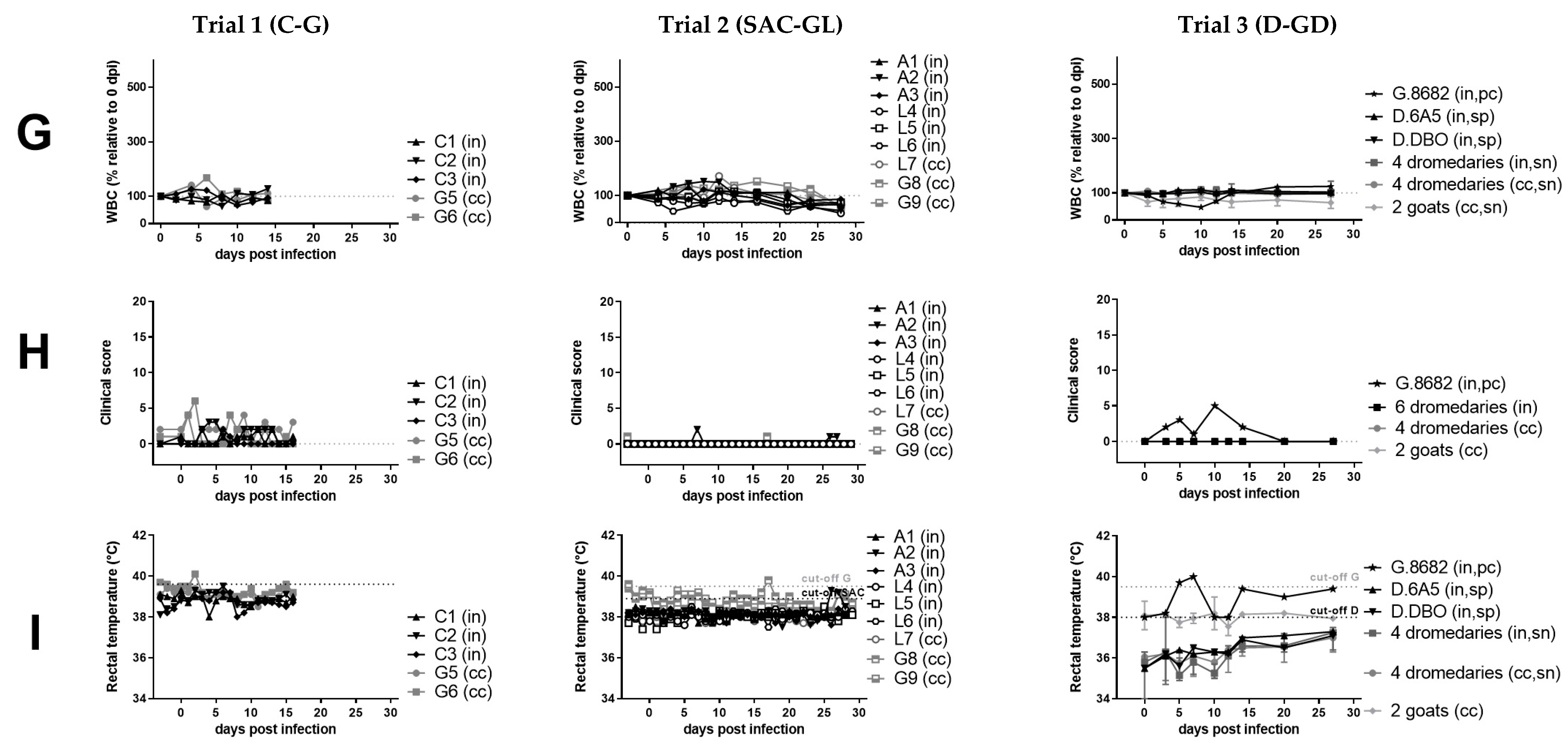
3. Results
3.1. Clinical and Hematological Results Suggest Resistance of Camelids and Cattle to PPR-Induced Disease
3.2. Humoral Response to PPRV Is Weak in Camelids but Pronounced in Cattle
3.3. Molecular Detection of PPRV-RNA in Swab Samples from Camelids and Cattle
3.4. PPRV-Antigen or RNA Detection in Lymphoreticular Tissue from SAC and Cattle
3.4.1. Gross Pathological Results
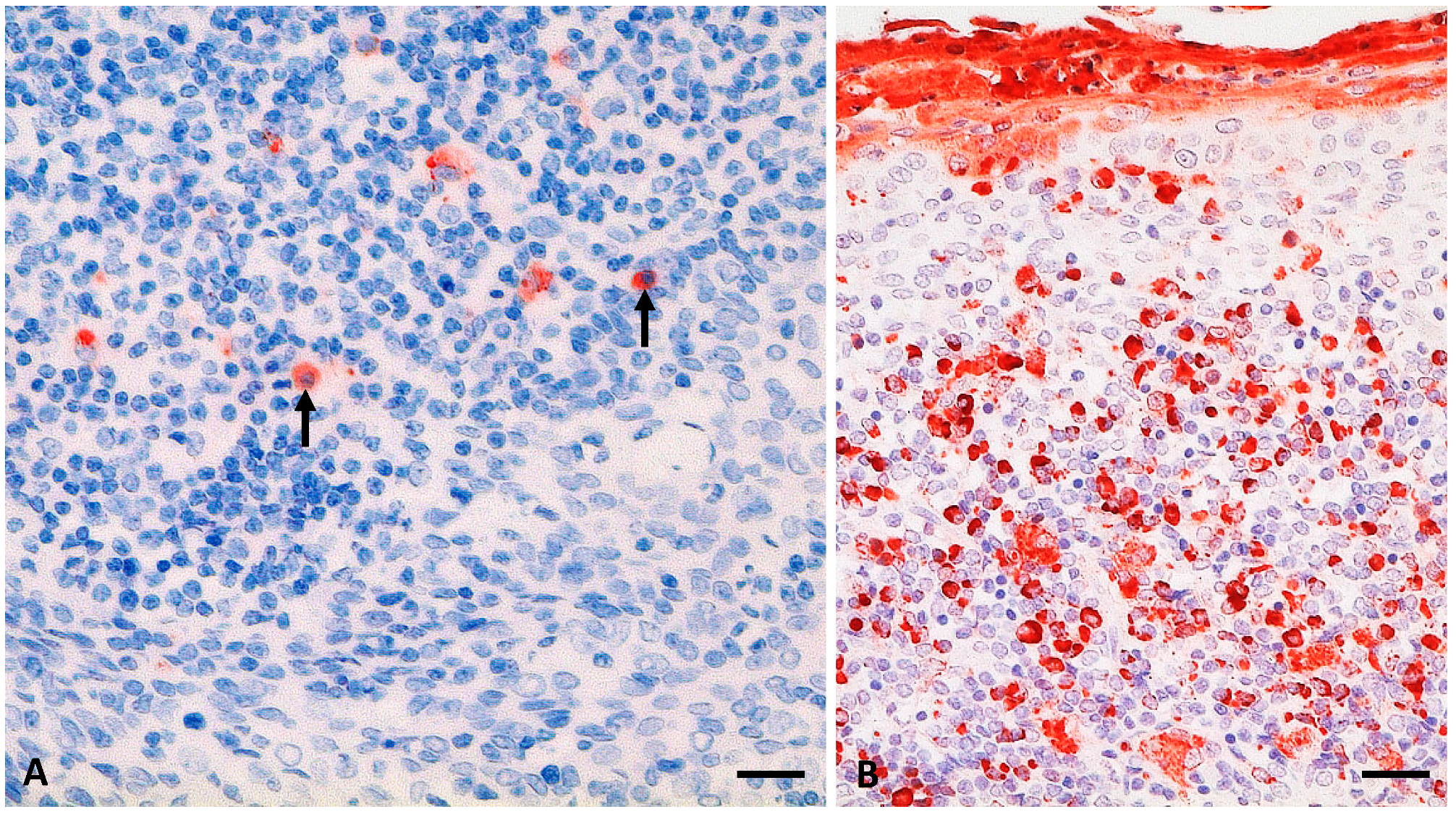
3.4.2. Molecular, Histopathological, and Immunohistochemical Results
South American Camelids
Cattle
3.5. RT-qPCR Is the Most Sensitive Virological Method for Diagnosis of PPRV-Infection in SAC and Cattle
4. Discussion
4.1. Clinical Signs Associated with PPRV-Infection
4.2. PPRV Detection in Camels
4.3. PPRV-Transmission
4.4. PPRV Antibody Response and Performance of Diagnostic Methods
4.5. Pathogenesis, Virus, and Host Factors
4.6. Possible Impact of Ecology on PPRV Infection and Spread
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Amarasinghe, G.K.; Ayllon, M.A.; Bao, Y.; Basler, C.F.; Bavari, S.; Blasdell, K.R.; Briese, T.; Brown, P.A.; Bukreyev, A.; Balkema-Buschmann, A.; et al. Taxonomy of the order Mononegavirales: Update 2019. Arch. Virol. 2019, 164, 1967–1980. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Scientific Opinion on peste des petits ruminants. EFSA Panel on Animal Health and Welfare (AHAW). EFSA J. 2015, 13, 3985. [Google Scholar] [CrossRef]
- Wernike, K.; Eschbaumer, M.; Breithaupt, A.; Maltzan, J.; Wiesner, H.; Beer, M.; Hoffmann, B. Experimental infection of sheep and goats with a recent isolate of peste des petits ruminants virus from Kurdistan. Vet. Microbiol. 2014, 172, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Pope, R.A.; Parida, S.; Bailey, D.; Brownlie, J.; Barrett, T.; Banyard, A.C. Early events following experimental infection with Peste-Des-Petits ruminants virus suggest immune cell targeting. PLoS ONE 2013, 8, e55830. [Google Scholar] [CrossRef]
- Munir, M. Role of wild small ruminants in the epidemiology of peste des petits ruminants. Transbound. Emerg. Dis. 2013. [Google Scholar] [CrossRef] [PubMed]
- Libeau, G.; Diallo, A.; Parida, S. Evolutionary genetics underlying the spread of peste des petits ruminants virus. Anim. Front. 2014, 4, 14–20. [Google Scholar] [CrossRef]
- Kock, R.A.; Wambua, J.M.; Mwanzia, J.; Wamwayi, H.; Ndungu, E.K.; Barrett, T.; Kock, N.D.; Rossiter, P.B. Rinderpest epidemic in wild ruminants in Kenya 1993-97. Vet. Rec. 1999, 145, 275–283. [Google Scholar] [CrossRef]
- World Organisation for Animal Health (OIE). Rinderpest. OIE Technical Disease Cards. 2013. Available online: https://www.oie.int/fileadmin/Home/eng/Animal_Health_in_the_World/docs/pdf/Disease_cards/RINDERPEST.pdf (accessed on 6 December 2019).
- Schulz, C.; Fast, C.; Schlottau, K.; Hoffmann, B.; Beer, M. Neglected hosts of small ruminant morbillivirus. Emerg. Infect. Dis. 2018, 24, 2334–2337. [Google Scholar] [CrossRef]
- Kinne, J.; Kreutzer, R.; Kreutzer, M.; Wernery, U.; Wohlsein, P. Peste des petits ruminants in Arabian wildlife. Epidemiol. Infect. 2010, 138, 1211–1214. [Google Scholar] [CrossRef]
- Intisar, K.S.; Ali, Y.H.; Haj, M.A.; Sahar, M.A.; Shaza, M.M.; Baraa, A.M.; Ishag, O.M.; Nouri, Y.M.; Taha, K.M.; Nada, E.M.; et al. Peste des petits ruminants infection in domestic ruminants in Sudan. Trop. Anim. Health Prod. 2017, 49, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Kwiatek, O.; Ali, Y.H.; Saeed, I.K.; Khalafalla, A.I.; Mohamed, O.I.; Obeida, A.A.; Abdelrahman, M.B.; Osman, H.M.; Taha, K.M.; Abbas, Z.; et al. Asian lineage of peste des petits ruminants virus, Africa. Emerg. Infect. Dis. 2011, 17, 1223–1231. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, R.; Koteeswaran, A.; Venugopalan, A.T.; Shyam, G.; Shaouna, S.; Shaila, M.S.; Ramachandran, S. Isolation of pestes des petits ruminants virus from an outbreak in Indian buffalo (Bubalus bubalis). Vet. Rec. 1997, 141, 573–574. [Google Scholar] [CrossRef] [PubMed]
- Sen, A.; Saravanan, P.; Balamurugan, V.; Bhanuprakash, V.; Venkatesan, G.; Sarkar, J.; Rajak, K.K.; Ahuja, A.; Yadav, V.; Sudhakar, S.B.; et al. Detection of subclinical peste des petits ruminants virus infection in experimental cattle. Virusdisease 2014, 25, 408–411. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Roger, F.; Yigezu, L.M.; Hurard, C.; Libeau, G.; Mebratu, G.Y.; Diallo, A.; Faye, B. Investigations on a new pathological condition of camels in Ethiopia. J. Camel Pract. Res. 2000, 7, 163–165. [Google Scholar]
- Khalafalla, A.I.; Saeed, I.K.; Ali, Y.H.; Abdurrahman, M.B.; Kwiatek, O.; Libeau, G.; Obeida, A.A.; Abbas, Z. An outbreak of peste des petits ruminants (PPR) in camels in the Sudan. Acta Trop. 2010, 116, 161–165. [Google Scholar] [CrossRef]
- Zakian, A.; Nouri, M.; Kahroba, H.; Mohammadian, B.; Mokhber-Dezfouli, M.R. The first report of peste des petits ruminants (PPR) in camels (Camelus dromedarius) in Iran. Trop. Anim. Health Prod. 2016, 48, 1215–1219. [Google Scholar] [CrossRef]
- World Organisation for Animal Health (OIE). Peste des Petits Ruminants. OIE Technical Disease Cards. 2013. Available online: https://www.oie.int/fileadmin/Home/eng/Animal_Health_in_the_World/docs/pdf/Disease_cards/PESTE_DES_PETITS_RUMINANTS.pdf (accessed on 6 December 2019).
- El-Hakim, U.A. The role of camels in dessimination of peste des petits ruminants virus among sheep and goats in Saudi Arabia. Assiut Vet. Med. J. 2006, 52, 132–145. [Google Scholar]
- Nawathe, D.R.; Taylor, W.P. Experimental infection of domestic pigs with the virus of peste des petits ruminants. Trop. Anim. Health Prod. 1979, 11, 120–122. [Google Scholar] [CrossRef]
- Taylor, W.P. The susceptibility of the one-humped camel (Camelus dromedarius) to infection with rinderpest virus. Bull. Epizoot. Dis. Afr. 1968, 16, 405–410. [Google Scholar] [PubMed]
- Nugent, G. Maintenance, spillover and spillback transmission of bovine tuberculosis in multi-host wildlife complexes: A New Zealand case study. Vet. Microbiol. 2011, 151, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.V.; Thacker, T.C.; Waters, W.R.; Gortazar, C.; Corner, L.A. Mycobacterium bovis: A Model Pathogen at the Interface of Livestock, Wildlife, and Humans. Vet. Med. Int. 2012, 2012, 236205. [Google Scholar] [CrossRef] [PubMed]
- Fritzsche McKay, A.; Hoye, B.J. Are Migratory Animals Superspreaders of Infection? Integr. Comp. Biol. 2016, 56, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Schulz, C.; Becker, S.C. Mosquitoes as arbovirus vectors: From species identification to vector competence. In Mosquito-Borne Diseases: Implications for Public Health; Benelli, G., Mehlhorn, H., Eds.; Series: Parasitology Research Monographs 10; Springer: Cham, Switzerland, 2018; pp. 163–212. [Google Scholar]
- Plowright, R.K.; Parrish, C.R.; McCallum, H.; Hudson, P.J.; Ko, A.I.; Graham, A.L.; Lloyd-Smith, J.O. Pathways to zoonotic spillover. Nat. Rev. Microbiol. 2017, 15, 502–510. [Google Scholar] [CrossRef]
- Bowden, S.E.; Drake, J.M. Ecology of multi-host pathogens of animals. Nat. Educ. Knowl. 2013, 4, 5. [Google Scholar]
- Truong, T.; Boshra, H.; Embury-Hyatt, C.; Nfon, C.; Gerdts, V.; Tikoo, S.; Babiuk, L.A.; Kara, P.; Chetty, T.; Mather, A.; et al. Peste des petits ruminants virus tissue tropism and pathogenesis in sheep and goats following experimental infection. PLoS ONE 2014, 9, e87145. [Google Scholar] [CrossRef]
- Bataille, A.; Kwiatek, O.; Belfkhi, S.; Mounier, L.; Parida, S.; Mahapatra, M.; Caron, A.; Chubwa, C.C.; Keyyu, J.; Kock, R.; et al. Optimization and evaluation of a non-invasive tool for peste des petits ruminants surveillance and control. Sci. Rep. 2019, 9, 4742. [Google Scholar] [CrossRef]
- Abdullah, N.; Kelly, J.T.; Graham, S.C.; Birch, J.; Goncalves-Carneiro, D.; Mitchell, T.; Thompson, R.N.; Lythgoe, K.A.; Logan, N.; Hosie, M.J.; et al. Structure-Guided Identification of a Nonhuman Morbillivirus with Zoonotic Potential. J. Virol. 2018, 92. [Google Scholar] [CrossRef]
- Matrosovich, M.; Herrler, G.; Klenk, H.D. Sialic Acid Receptors of Viruses. Top. Curr. Chem. 2015, 367, 1–28. [Google Scholar] [CrossRef]
- Parida, S.; Muniraju, M.; Mahapatra, M.; Muthuchelvan, D.; Buczkowski, H.; Banyard, A.C. Peste des petits ruminants. Vet. Microbiol. 2015, 181, 90–106. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Maherchandani, S.; Kashyap, S.K.; Singh, S.V.; Sharma, S.; Chaubey, K.K.; Ly, H. Peste des petits ruminants virus infection of small ruminants: A comprehensive review. Viruses 2014, 6, 2287–2327. [Google Scholar] [CrossRef] [PubMed]
- Jo, W.K.; Kruppa, J.; Habierski, A.; van de Bildt, M.; Mazzariol, S.; Di Guardo, G.; Siebert, U.; Kuiken, T.; Jung, K.; Osterhaus, A.; et al. Evolutionary evidence for multi-host transmission of cetacean morbillivirus. Emerg. Microbes Infect. 2018, 7, 201. [Google Scholar] [CrossRef] [PubMed]
- Jo, W.K.; Osterhaus, A.D.; Ludlow, M. Transmission of morbilliviruses within and among marine mammal species. Curr. Opin. Virol. 2018, 28, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Ludlow, M.; Rennick, L.J.; Nambulli, S.; de Swart, R.L.; Duprex, W.P. Using the ferret model to study morbillivirus entry, spread, transmission and cross-species infection. Curr. Opin. Virol. 2014, 4, 15–23. [Google Scholar] [CrossRef]
- Sawatsky, B.; Cattaneo, R.; von Messling, V. Canine Distemper Virus Spread and Transmission to Naive Ferrets: Selective Pressure on Signaling Lymphocyte Activation Molecule-Dependent Entry. J. Virol. 2018, 92. [Google Scholar] [CrossRef]
- De Vries, R.D.; Duprex, W.P.; de Swart, R.L. Morbillivirus infections: An introduction. Viruses 2015, 7, 699–706. [Google Scholar] [CrossRef]
- Beineke, A.; Puff, C.; Seehusen, F.; Baumgartner, W. Pathogenesis and immunopathology of systemic and nervous canine distemper. Vet. Immunol. Immunopathol. 2009, 127, 1–18. [Google Scholar] [CrossRef]
- Birch, J.; Juleff, N.; Heaton, M.P.; Kalbfleisch, T.; Kijas, J.; Bailey, D. Characterization of ovine Nectin-4, a novel peste des petits ruminants virus receptor. J. Virol. 2013, 87, 4756–4761. [Google Scholar] [CrossRef]
- Hoffmann, B.; Wiesner, H.; Maltzan, J.; Mustefa, R.; Eschbaumer, M.; Arif, F.A.; Beer, M. Fatalities in wild goats in Kurdistan associated with Peste des Petits Ruminants virus. Transbound. Emerg. Dis. 2012, 59, 173–176. [Google Scholar] [CrossRef]
- Adombi, C.M.; Lelenta, M.; Lamien, C.E.; Shamaki, D.; Koffi, Y.M.; Traore, A.; Silber, R.; Couacy-Hymann, E.; Bodjo, S.C.; Djaman, J.A.; et al. Monkey CV1 cell line expressing the sheep-goat SLAM protein: A highly sensitive cell line for the isolation of peste des petits ruminants virus from pathological specimens. J. Virol. Methods 2011, 173, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Von Messling, V.; Springfeld, C.; Devaux, P.; Cattaneo, R. A ferret model of canine distemper virus virulence and immunosuppression. J. Virol. 2003, 77, 12579–12591. [Google Scholar] [CrossRef] [PubMed]
- Jenckel, M.; Béard, E.; Schulz, C.; Sailleau, C.; Viarouge, C.; Hoffmann, B.; Höper, D.; Beer, M.; Zientara, S. Complete coding genome sequence of putative novel bluetongue virus serotype 27. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Schulz, C.; Bréard, E.; Sailleau, C.; Jenckel, M.; Viarouge, C.; Vitour, D.; Palmarini, M.; Gallois, M.; Höper, D.; Hoffmann, B.; et al. Bluetongue virus serotype 27: Detection and characterization of two novel variants in Corsica, France. J. Gen. Virol. 2016, 97, 2073–2083. [Google Scholar] [CrossRef]
- Stober, M. Kennzeichen, Anamnese, Grundregeln der Untersuchungstechnik, Allgemeine Untersuchung. In Die Klinische Untersuchung des Rindes, 3rd ed.; Rosenberger, G., Ed.; Parey: Berlin/Hamburg, Germany, 1990; pp. 75–141. [Google Scholar]
- Fowler, M.E. Clinical Diagnosis: Examination and Procedures. In Medicine and Surgery of Camelids; Fowler, M.E., Ed.; Blackwell Publishing: Ames, IA, USA, 2010; pp. 89–109. [Google Scholar]
- Libeau, G.; Prehaud, C.; Lancelot, R.; Colas, F.; Guerre, L.; Bishop, D.H.; Diallo, A. Development of a competitive ELISA for detecting antibodies to the peste des petits ruminants virus using a recombinant nucleoprotein. Res. Vet. Sci. 1995, 58, 50–55. [Google Scholar] [CrossRef]
- Mayr, A.; Bachmann, P.A.; Bibrack, B.; Wittmann, G. Quantitative Bestimmung der Virusinfektiosität (Virustitration). In Virologische Arbeitsmethoden; Band, I., Mayr, A., Bachmann, P.A., Bibrack, B., Wittmann, G., Eds.; VEB Gustav Fischer Verlag: Jena, Germany, 1974; pp. 35–39. [Google Scholar]
- Batten, C.A.; Banyard, A.C.; King, D.P.; Henstock, M.R.; Edwards, L.; Sanders, A.; Buczkowski, H.; Oura, C.C.; Barrett, T. A real time RT-PCR assay for the specific detection of Peste des petits ruminants virus. J. Virol. Methods 2011, 171, 401–404. [Google Scholar] [CrossRef]
- Hoffmann, B.; Depner, K.; Schirrmeier, H.; Beer, M. A universal heterologous internal control system for duplex real-time RT-PCR assays used in a detection system for pestiviruses. J. Virol. Methods 2006, 136, 200–209. [Google Scholar] [CrossRef]
- Couacy-Hymann, E.; Koffi, M.Y.; Kouadio, V.K.; Mossoum, A.; Kouadio, L.; Kouassi, A.; Assemian, K.; Godji, P.H.; Nana, P. Experimental infection of cattle with wild type peste-des-petits-ruminants virus—Their role in its maintenance and spread. Res. Vet. Sci. 2019, 124, 118–122. [Google Scholar] [CrossRef]
- Saeed, I.K.; Ali, Y.H.; AbdulRahman, M.B.; Mohammed, Z.A.; Osman, H.M.; Taha, K.M.; Musa, M.Z.; Khalafalla, A.I. Mixed infection of peste des petits ruminants virus (PPRV) and other respiratory viruses in dromedary camels in Sudan, an abattoir study. Trop. Anim. Health Prod. 2015, 47, 995–998. [Google Scholar] [CrossRef]
- Fakri, F.Z.; Bamouh, Z.; Jazouli, M.; Omari Tadlaoui, K.; Elharrak, M. Experimental infection of dromedary camels with virulent virus of Peste des Petits Ruminants. Vet. Microbiol. 2019, 235, 195–198. [Google Scholar] [CrossRef]
- Mornet, P.; Gilbert, Y.; Orue, J.; Thiéry, G. La peste des petits ruminants en Afrique occidentale française. Ses rapports avec la peste bovine. Revue D’élevage et de Médecine Vétérinaire des Pays Tropicaux 1956, 9, 313–342. [Google Scholar] [CrossRef]
- Wernery, U. Peste des petits ruminants (PPR) in camelids with own investigations. J. Camel Pract. Res. 2011, 18, 219–223. [Google Scholar]
- Singh, K.V.; Ata, F. Experimental rinderpest in camels. A preliminary report. Bull. Epizoot. Dis. Afr. 1967, 15, 19–23. [Google Scholar] [PubMed]
- Hammouchi, M.; Loutfi, C.; Sebbar, G.; Touil, N.; Chaffai, N.; Batten, C.; Harif, B.; Oura, C.; El Harrak, M. Experimental infection of alpine goats with a Moroccan strain of peste des petits ruminants virus (PPRV). Vet. Microbiol. 2012, 160, 240–244. [Google Scholar] [CrossRef]
- Wernery, U.; Kinne, J.; Schuster, R.K. Peste des petits ruminants. In Camelid Infectious Disorders; Wernery, U., Kinne, J., Schuster, R.K., Eds.; OIE, World Organisation for Animal Health: Paris, France, 2014; pp. 257–261. [Google Scholar]
- Haroun, M.; Hajer, I.; Mukhtar, M.; Ali, B.E. Detection of antibodies against peste des petits ruminants virus in sera of cattle, camels, sheep and goats in Sudan. Vet. Res. Commun. 2002, 26, 537–541. [Google Scholar] [CrossRef]
- Omani, R.N.; Gitao, G.C.; Gachohi, J.; Gathumbi, P.K.; Bwihangane, B.A.; Abbey, K.; Chemweno, V.J. Peste Des Petits Ruminants (PPR) in Dromedary Camels and Small Ruminants in Mandera and Wajir Counties of Kenya. Adv. Virol. 2019, 2019, 4028720. [Google Scholar] [CrossRef]
- Megersa, B.; Biffa, D.; Abunna, F.; Regassa, A.; Bohlin, J.; Skjerve, E. Epidemic characterization and modeling within herd transmission dynamics of an “emerging trans-boundary” camel disease epidemic in Ethiopia. Trop. Anim. Health Prod. 2012, 44, 1643–1651. [Google Scholar] [CrossRef]
- Scott, G.R.; MacDonald, J. Kenya camels and rinderpest. Bull. Epizoot. Dis. Afr. 1962, 10, 495–497. [Google Scholar]
- Woma, T.Y.; Kalla, D.J.; Ekong, P.S.; Ularamu, H.G.; Chollom, S.C.; Lamurde, I.I.; Bajehson, D.B.; Tom, N.D.; Aaron, G.B.; Shamaki, D.; et al. Serological evidence of camel exposure to peste des petits ruminants virus (PPRV) in Nigeria. Trop. Anim. Health Prod. 2015, 47, 603–606. [Google Scholar] [CrossRef]
- Wernery, U.; Kinne, J. Foot and mouth disease and similar virus infections in camelids: A review. Rev. Sci. Tech. 2012, 31, 907–918. [Google Scholar] [CrossRef]
- Schulz, C.; Eschbaumer, M.; Rudolf, M.; König, P.; Keller, M.; Bauer, C.; Gauly, M.; Grevelding, C.G.; Beer, M.; Hoffmann, B. Experimental infection of South American camelids with bluetongue virus serotype 8. Vet. Microbiol. 2012, 154, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Schulz, C.; Beer, M.; Hoffmann, B. Schmallenberg virus infection in South American camelids: Field and experimental investigations. Vet. Microbiol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Caron, A.; Cappelle, J.; Cumming, G.S.; de Garine-Wichatitsky, M.; Gaidet, N. Bridge hosts, a missing link for disease ecology in multi-host systems. Vet. Res. 2015, 46, 83. [Google Scholar] [CrossRef] [PubMed]
- Hurst, C.J.; Lindquist, H.D.A. Defining the ecology of viruses. In Viral Ecology; Hurst, C.J., Ed.; Academic Press: San Diego, CA, USA, 2000; pp. 3–40. [Google Scholar] [CrossRef]
- Lin, W.H.; Kouyos, R.D.; Adams, R.J.; Grenfell, B.T.; Griffin, D.E. Prolonged persistence of measles virus RNA is characteristic of primary infection dynamics. Proc. Natl. Acad. Sci. USA 2012, 109, 14989–14994. [Google Scholar] [CrossRef] [PubMed]
- De Witte, L.; de Vries, R.D.; van der Vlist, M.; Yuksel, S.; Litjens, M.; de Swart, R.L.; Geijtenbeek, T.B. DC-SIGN and CD150 have distinct roles in transmission of measles virus from dendritic cells to T-lymphocytes. PLoS Pathog. 2008, 4, e1000049. [Google Scholar] [CrossRef] [PubMed]
- Albayrak, H.; Gür, S. A serologic investigation for Peste des petits ruminants infection in sheep, cattle and camels (Camelus dromedarius) in Aydin province, West Anatolia. Trop. Anim. Health Prod. 2010, 42, 151–153. [Google Scholar] [CrossRef]
- Balamurugan, V.; Sen, A.; Venkatesan, G.; Bhanuprakash, V.; Singh, R.K. Protective immune response of live attenuated thermo-adapted peste des petits ruminants vaccine in goats. Virusdisease 2014, 25, 350–357. [Google Scholar] [CrossRef][Green Version]
- Altan, E.; Parida, S.; Mahapatra, M.; Turan, N.; Yilmaz, H. Molecular characterization of Peste des petits ruminants viruses in the Marmara Region of Turkey. Transbound. Emerg. Dis. 2018. [Google Scholar] [CrossRef]
- Albina, E.; Kwiatek, O.; Minet, C.; Lancelot, R.; Servan de Almeida, R.; Libeau, G. Peste des Petits Ruminants, the next eradicated animal disease? Vet. Microbiol. 2013, 165, 38–44. [Google Scholar] [CrossRef]
- Fournié, G.; Waret-Szkuta, A.; Camacho, A.; Yigezu, L.M.; Pfeiffer, D.U.; Roger, F. A dynamic model of transmission and elimination of peste des petits ruminants in Ethiopia. Proc. Natl. Acad. Sci. USA 2018, 115, 8454–8459. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trial No. | Trial ID | Intranasally Inoculated Animals | Contact Control Animals | Outcome of Experiment | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Species * and ID | Sex | Age (Months) | Species * and ID | Sex | Age (Months) | Seroconver-sion (Total No. by Spp.) | Shedding of PPRV-RNA (Total No. by Spp.) | Shedding of Infectious PPRV | Contact Transmission | ||
| 1 | C–G | C1–C3 | 3 f | 3 | G5, G6 | 2 m | 4 | 3 C | 3 C | no | no |
| 2 | SAC-GL | A1–A3 L4–L6 | 3 m 3 m | 6–10 7–8 | L7 | 1 m | 9 | 3 A, 3 L | 3 A, 2 L | no | no |
| G8, G9 | 2 m | 12–13 | 0 G | 0 G | no | no | |||||
| 3 | D-GD (pen 1) | D.54A, D.6A5, D.BO | 3 m | young adult (72) | D.204, D.05E | 2 f | young adult (72) | 2 D | 0 D | no | no |
| G.9499, G.9500 | 2 m | 24 | 0 G | 0 G | no | no | |||||
| 3 | D-GD (pen 2) | D.54A, D.F7B, D.No2 | 1 m, 2 f | old (216) | D.No1, D.No6 | 1 m, 1 f | old (170) | 0 D | 0 D | no | no |
| 3 | D-GD (pen 3) | G.8682 † | 1 m | 24 | none | 1 G | 1 G | yes | NA | ||
| (A) | ||||||||||||
| Animal Trial ID | C-G | C-G | C-G | SAC-GL | SAC-GL | SAC-GL | SAC-GL | SAC-GL | SAC-GL | D-GD | ||
| Animal ID | C1 | C2 | C3 | A1 | A2 | A3 | L4 | L5 | L6 | pc-G | ||
| dpi | 17 | 17 | 17 | 28 | 28 | 28 | 28 | 29 | 29 | 35 | ||
| Organ Location | Organ ID | Tissue (Cq) | ||||||||||
| head | 1 | third eye lid | 33.63 | 34.11 | 33.17 | - | - | - | - | - | - | 39.66 |
| 2 | lacrimal gland | - | - | - | - | - | - | - | - | - | 34.39 | |
| 6 | tongue (apex) | nd | nd | nd | - | - | - | - | - | - | nd | |
| 7 | palatine tonsil | 33.26 | 34.80 | 30.55 | 33.88 | - | - | - | - | - | 32.78 | |
| 8 | retropharyngeal ln. | 32.83 | 32.20 | 31.45 | - | 35.32 | 37.42 | - | - | - | 31.48 | |
| 9 | mandibular ln. | nd | nd | nd | 33.21 | 34.56 | - | - | - | 33.28 | nd | |
| 10 | parotideal ln. | nd | nd | nd | 35.66 | - | - | - | - | - | nd | |
| cervical † | 11 | trachea | - | - | - | - | - | - | - | - | - | nd |
| 12 | esophagus | - | - | - | - | - | - | - | - | - | nd | |
| thoracal | 13 | lung | - | - | - | - | - | - | - | - | - | nd |
| 14 | bronchial ln. | - | 34.84 | 35.41 | 34.94 | - | - | - | - | 32.93 | 32.28 | |
| 15 | mediastinal ln. | 36.07 | - | - | 32.56 | - | - | - | - | 35.08 | 31.07 | |
| abdominal | 19 | jejunal PP | 36.20 | 37.52 | 33.08 | 33.05 | - | - | - | - | - | nd |
| 20 | ileal PP | - | 38.43 | 33.82 | - | - | - | - | - | - | nd | |
| 21 | caecum | - | - | - | - | - | - | - | - | - | nd | |
| 22 | colon | - | 32.75 | - | - | - | - | - | - | - | 34.41 | |
| 23 | mesenteric ln. | - | 34.69 | 32.80 | - | - | - | - | - | - | 32.17 | |
| 24 | rectum | - | 38.43 | 33.82 | - | - | - | - | - | - | nd | |
| 27 | spleen | 37.86 | - | - | - | - | - | - | - | - | 34.28 | |
| cerebral | 30 | different brain tissues ‡ | nd | nd | nd | - | - | - | - | - | - | - |
| (B) | ||||||||||||
| Animal Trial ID | C-G | C-G | C-G | SAC-GL | SAC-GL | SAC-GL | SAC-GL | SAC-GL | SAC-GL | D-GD | ||
| Animal ID | C1 | C2 | C3 | A1 | A2 | A3 | L4 | L5 | L6 | pc-G | ||
| dpi | 17 | 17 | 17 | 28 | 28 | 28 | 28 | 29 | 29 | 35 | ||
| Organ Location | Organ ID | Tissue (IHC) | ||||||||||
| head | 1 | third eye lid * | nd | - | - | - | - | - | - | - | - | nd |
| 2 | lacrimal gland | - | - | - | - | - | - | - | - | - | nd | |
| 6 | tongue (apex) | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | |
| 7 | palatine tonsil * | - | - | - | - | ++ | + | - | - | - | nd | |
| 8 | retropharyngeal ln. | - | - | - | - | - | - | - | - | - | nd | |
| 9 | mandibular ln. | nd | nd | nd | - | nd | - | - | - | - | nd | |
| 10 | parotideal ln. | nd | nd | nd | - | - | - | - | - | - | nd | |
| cervical † | 11 | trachea | - | - | - | - | - | - | - | - | - | nd |
| 12 | esophagus | - | - | - | - | - | - | - | - | - | nd | |
| thoracal | 13 | lung | - | - | - | - | - | - | - | - | - | nd |
| 14 | bronchial ln. | - | - | - | - | - | - | - | - | - | nd | |
| 15 | mediastinal ln. | nd | - | - | - | nd | nd | - | - | - | nd | |
| abdominal | 19 | jejunal PP | - | - | - | - | nd | - | - | - | - | nd |
| 20 | ileal PP | nd | - | - | - | - | - | - | - | - | nd | |
| 21 | caecum * | - | - | - | - | + | - | - | - | - | nd | |
| 22 | colon * | - | - | - | - | - | + | - | - | - | nd | |
| 23 | mesenteric ln. | - | - | - | - | - | - | - | - | - | nd | |
| 24 | rectum * | - | - | - | - | nd | - | - | - | - | nd | |
| 27 | spleen | - | - | - | - | - | - | - | - | - | nd | |
| cerebral | 30 | different brain tissues ‡ | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd |
| Serial No. | Animal Trial ID | Animal Species | Sample Material | Animal ID | dpi | SLAM-Cells (Max. TCID50/mL on VDS or CHS) * | Detection of PPRV-Np by RT-qPCR (Cq Value) | Ag-ELISA (OD NC %) | LFD (pos/neg) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| (Trial No.) | Bao et al. 2008 | Batten et al. 2011 | |||||||||
| 35 | C-G (1) | cattle | oronasal | swab | C3 | 5 | neg | 27.02 | 28.22 | 13 | neg |
| 36 | C-G (1) | cattle | oronasal | swab | C1 | 6 | neg | 32.01 | 30.73 | 46 | neg |
| 37 | C-G (1) | cattle | fecal | swab | C3 | 7 | neg | 40.28 | 35.48 | −12 | neg |
| 38 | C-G (1) | cattle | oronasal | swab | C3 | 7 | neg | 30.16 | 31.43 | 15 | neg |
| 39 | C-G (1) | cattle | conjunctival | swab | C2 | 7 | neg | 32.17 | 32.23 | 1 | neg |
| 40 | C-G (1) | cattle | mediastinal ln. | tissue | C1 | 17 | neg | 44.16 | 36.07 | −12 | neg |
| 41 | C-G (1) | cattle | bronchial ln. | tissue | C2 | 17 | neg | 36.11 | 34.84 | −17 | neg |
| 42 | C-G (1) | cattle | palatine tonsil | tissue | C3 | 17 | neg | 30.06 | 30.55 | 1 | neg |
| 43 | C-G (1) | cattle | retropharyng. ln. | tissue | C3 | 17 | neg | 31.84 | 31.45 | −6 | neg |
| 44 | C-G (1) | cattle | ileal peyer’s patches | tissue | C3 | 17 | neg | 33.36 | 34.11 | −11 | neg |
| 45 | SAC-GL (2) | llama | oronasal | swab | L5 | 4 | neg | 38.37 | 39.00 | −10 | neg |
| 46 | SAC-GL (2) | alpaca | oronasal | swab | A1 | 7 | neg | No Cq | 36.58 | 15 | neg |
| 47 | SAC-GL (2) | alpaca | oronasal | swab | A2 | 8 | neg | 39.21 | 33.85 | −13 | neg |
| 48 | SAC-GL (2) | alpaca | EDTA-blood | blood | A3 | 8 | neg | No Cq | 37.68 | 0 | neg |
| 49 | SAC-GL (2) | llama | fecal | swab | L6 | 14 | neg | No Cq | 38.61 | −24 | neg |
| 50 | SAC-GL (2) | alpaca | palatine tonsil | tissue | A1 | 28 | neg | No Cq | 33.88 | −21 | neg |
| 51 | SAC-GL (2) | alpaca | mediastinal ln. | tissue | A1 | 28 | neg | No Cq | 32.56 | 9 | neg |
| 52 | SAC-GL (2) | alpaca | retropharyng. ln. | tissue | A2 | 28 | neg | No Cq | 35.32 | 9 | neg |
| 53 | SAC-GL (2) | llama | mandibular ln. | tissue | L6 | 29 | neg | No Cq | 33.28 | −20 | neg |
| 54 | SAC-GL (2) | llama | bronchial ln. | tissue | L6 | 29 | neg | No Cq | 32.93 | −20 | neg |
| PPRV cell culture virus, strain Kurdistan/2011 | pos control | 10^5.5 | 18.56 | ND | ND | pos | |||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schulz, C.; Fast, C.; Wernery, U.; Kinne, J.; Joseph, S.; Schlottau, K.; Jenckel, M.; Höper, D.; Patteril, N.A.G.; Syriac, G.; et al. Camelids and Cattle Are Dead-End Hosts for Peste-des-Petits-Ruminants Virus. Viruses 2019, 11, 1133. https://doi.org/10.3390/v11121133
Schulz C, Fast C, Wernery U, Kinne J, Joseph S, Schlottau K, Jenckel M, Höper D, Patteril NAG, Syriac G, et al. Camelids and Cattle Are Dead-End Hosts for Peste-des-Petits-Ruminants Virus. Viruses. 2019; 11(12):1133. https://doi.org/10.3390/v11121133
Chicago/Turabian StyleSchulz, Claudia, Christine Fast, Ulrich Wernery, Jörg Kinne, Sunitha Joseph, Kore Schlottau, Maria Jenckel, Dirk Höper, Nissy Annie Georgy Patteril, Ginu Syriac, and et al. 2019. "Camelids and Cattle Are Dead-End Hosts for Peste-des-Petits-Ruminants Virus" Viruses 11, no. 12: 1133. https://doi.org/10.3390/v11121133
APA StyleSchulz, C., Fast, C., Wernery, U., Kinne, J., Joseph, S., Schlottau, K., Jenckel, M., Höper, D., Patteril, N. A. G., Syriac, G., Hoffmann, B., & Beer, M. (2019). Camelids and Cattle Are Dead-End Hosts for Peste-des-Petits-Ruminants Virus. Viruses, 11(12), 1133. https://doi.org/10.3390/v11121133