A Novel Hepadnavirus is Associated with Chronic Hepatitis and Hepatocellular Carcinoma in Cats

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Case Selection and Tissues
2.2. DNA Extraction
2.3. Conventional PCR Assays
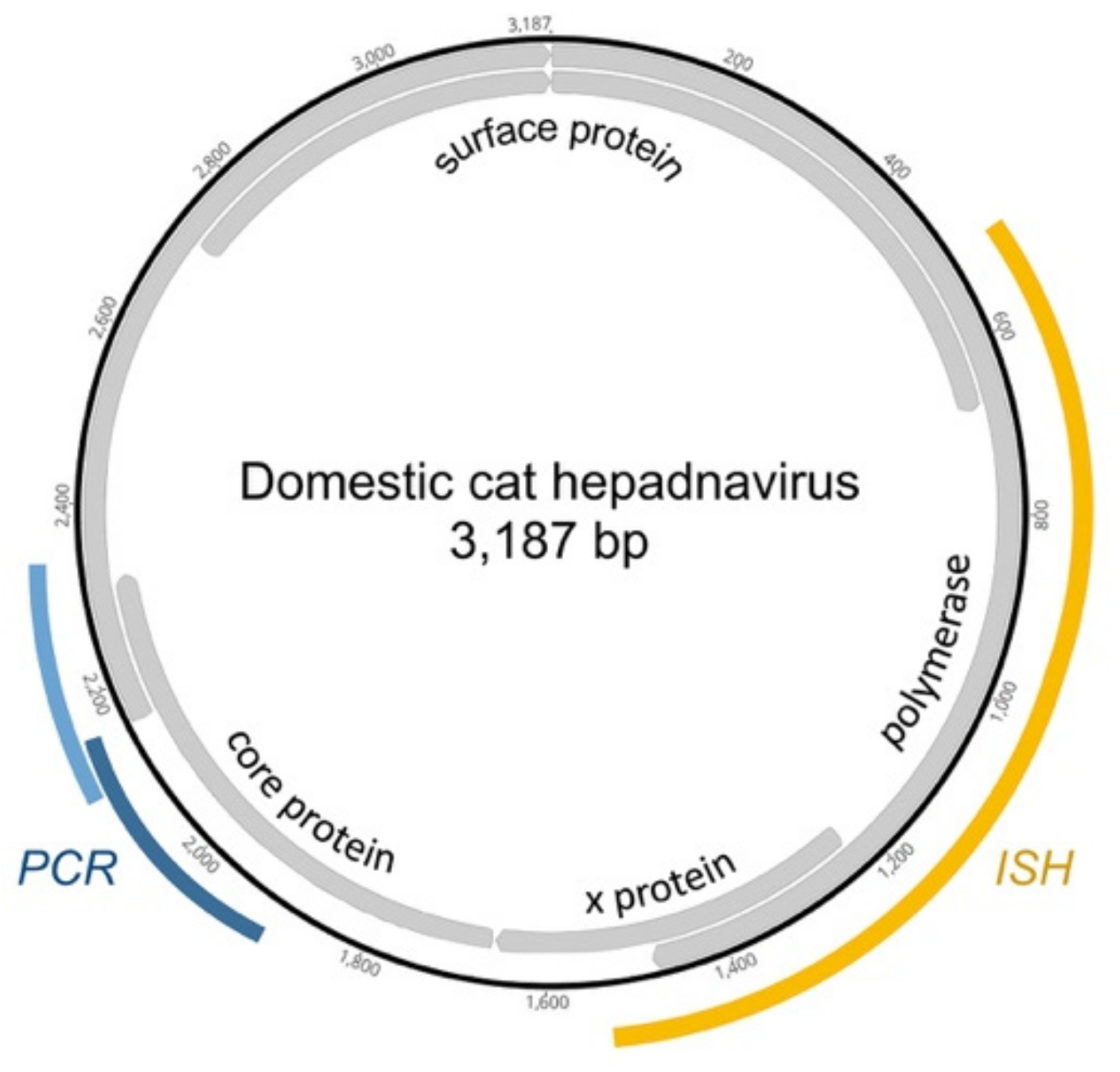
2.4. In Situ Hybridization
2.5. Immunohistochemistry
3. Results
3.1. Cases and Controls
3.2. Detection of DCH in Feline Liver Biopsies
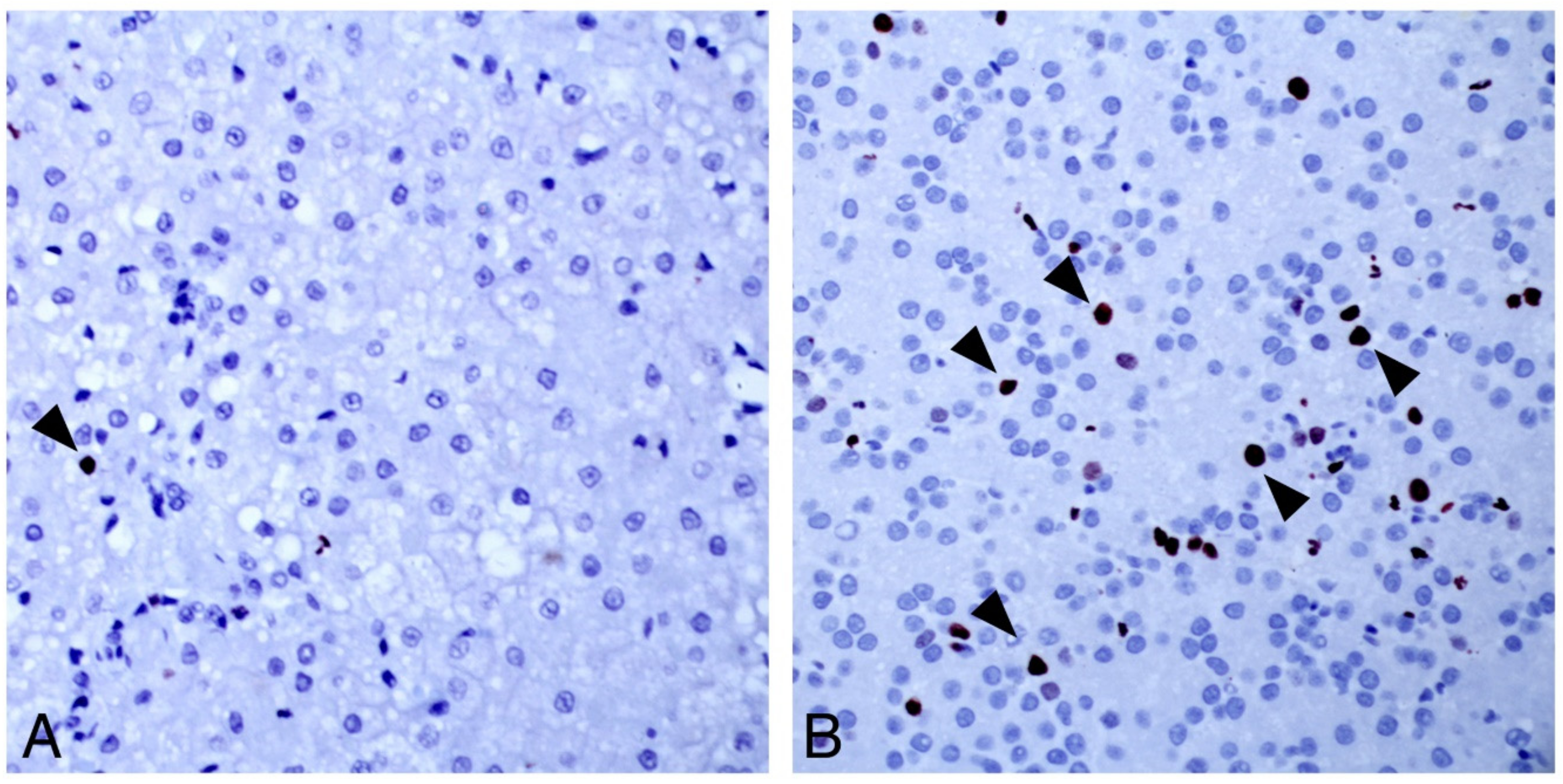
3.3. Character of DCH-Associated Lesions by Traditional Pathology and ISH
3.3.1. Chronic Hepatitis
3.3.2. Hepatocellular Carcinoma
3.4. DCH-Negative Lesions
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organisation. Global Hepatitis Report 2017; World Health Organisation: Geneva, Switzerland, 2017; ISBN 978-92-4-156545-5. [Google Scholar]
- Guidotti, L.G.; Isogawa, M.; Chisari, F.V. Host-virus interactions in hepatitis B virus infection. Curr. Opin. Immunol. 2015, 36, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Fattovich, G.; Bortolotti, F.; Donato, F. Natural history of chronic hepatitis B: Special emphasis on disease progression and prognostic factors. J. Hepatol. 2008, 48, 335–352. [Google Scholar] [CrossRef] [PubMed]
- Aghazadeh, M.; Shi, M.; Barrs, V.R.; McLuckie, A.J.; Lindsay, S.A.; Jameson, B.; Hampson, B.; Holmes, E.C.; Beatty, J.A. A Novel Hepadnavirus Identified in an Immunocompromised Domestic Cat in Australia. Viruses 2018, 10, 269. [Google Scholar] [CrossRef] [PubMed]
- Lanave, G.; Capozza, P.; Diakoudi, G.; Catella, C.; Catucci, L.; Ghergo, P.; Stasi, F.; Barrs, V.; Beatty, J.; DeCaro, N.; et al. Identification of hepadnavirus in the sera of cats. Sci. Rep. 2019, 9, 10668. [Google Scholar] [CrossRef] [PubMed]
- Bayton, W.A.; Westgarth, C.; Scase, T.; Price, D.J.; Bexfield, N.H. Histopathological frequency of feline hepatobiliary disease in the UK. J. Small Anim. Pract. 2018, 59, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Van den Ingh, T.S.G.; Van Winkle, T.; Cullen, J.M.; Charles, J.A.; Desmet, V.J. Morphological Classification of Parenchymal Disorders of the Canine and Feline Liver: 2. Hepatocellular Death, Hepatitis and Cirrhosis. In WSAVA Standards for Clinical and Histological Diagnosis of Canine and Feline Liver Diseases; Rothuizen, J., Bunch, S.E., Charles, J.A., Cullen, J.M., Desmet, V.J., Szatmári, V., Twedt, D.C., van den Ingh, T.S., Van Winkle, T., Washabau, R.J., Eds.; Saunders: Edinburgh, UK, 2006; pp. 85–101. [Google Scholar]
- Glebe, D.; Urban, S. Viral and cellular determinants involved in hepadnaviral entry. World J. Gastroenterol. 2007, 13, 22–38. [Google Scholar] [CrossRef] [PubMed]
- Drexler, J.F.; Geipel, A.; König, A.; Corman, V.M.; Van Riel, D.; Leijten, L.M.; Bremer, C.M.; Rasche, A.; Cottontail, V.M.; Maganga, G.D.; et al. Bats carry pathogenic hepadnaviruses antigenically related to hepatitis B virus and capable of infecting human hepatocytes. Proc. Natl. Acad. Sci. USA 2013, 110, 16151–16156. [Google Scholar] [CrossRef] [PubMed]
- Desmet, V.J.; Gerber, M.; Hoofnagle, J.H.; Manns, M.; Scheuer, P.J. Classification of chronic hepatitis: Diagnosis, grading and staging. Hepatology 1994, 19, 1513–1520. [Google Scholar] [CrossRef] [PubMed]
- Dandri, M.; Petersen, J. Mechanism of Hepatitis B Virus Persistence in Hepatocytes and Its Carcinogenic Potential. Clin. Infect. Dis. 2016, 62, S281–S288. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.D.; Kim, W.R.; Coelho, R.; Mettler, T.A.; Benson, J.T.; Sanderson, S.O.; Therneau, T.M.; Kim, B.; Roberts, L.R. Cirrhosis Is Present in Most Patients With Hepatitis B and Hepatocellular Carcinoma. Clin. Gastroenterol. Hepatol. 2011, 9, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.S.Y.; Covert, E.; Wilson, E.; Kottilil, S. Chronic Hepatitis B Infection A Review. J. Am. Med. Assoc. 2018, 319, 1802–1813. [Google Scholar] [CrossRef] [PubMed]
- American Veterinary Medical Association. AVMA Pet Ownership and Demographics Sourcebook (2017–2018); American Veterinary Medical Association: Schaumburg, IL, USA, 2018. [Google Scholar]
- Hammer, A.S.; Sikkema, D.A. Hepatic Neoplasia in the Dog and Cat. Veter. Clin. N. Am. Small Anim. Pract. 1995, 25, 419–435. [Google Scholar] [CrossRef]
- Cullen, J.M. Tumors of the Liver and Gallbladder. In Tumors in Domestic Animals; Wiley: Hoboken, NJ, USA, 2016; pp. 602–631. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Histopathological Diagnosis | Number of Cases | DCH Positive | |
|---|---|---|---|
| cPCR | ISH | ||
| Hepatocellular carcinoma | 29 | 8/29 | 8/8 |
| Biliary carcinoma | 18 | 0/18 | 0/11 |
| Hepatitis | 14 | 6/14 | 2/6 |
| Cholangitis | 6 | 0/6 | 0/5 |
| Other 1 | 13 | 0/4 | 0/3 |
| Normal liver 2 | 15 | 0/15 | 0/3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pesavento, P.A.; Jackson, K.; Scase, T.; Tse, T.; Hampson, B.; Munday, J.S.; Barrs, V.R.; Beatty, J.A. A Novel Hepadnavirus is Associated with Chronic Hepatitis and Hepatocellular Carcinoma in Cats. Viruses 2019, 11, 969. https://doi.org/10.3390/v11100969
Pesavento PA, Jackson K, Scase T, Tse T, Hampson B, Munday JS, Barrs VR, Beatty JA. A Novel Hepadnavirus is Associated with Chronic Hepatitis and Hepatocellular Carcinoma in Cats. Viruses. 2019; 11(10):969. https://doi.org/10.3390/v11100969
Chicago/Turabian StylePesavento, Patricia A., Kenneth Jackson, Timothy Scase, Tiffany Tse, Bronte Hampson, John S. Munday, Vanessa R. Barrs, and Julia A. Beatty. 2019. "A Novel Hepadnavirus is Associated with Chronic Hepatitis and Hepatocellular Carcinoma in Cats" Viruses 11, no. 10: 969. https://doi.org/10.3390/v11100969
APA StylePesavento, P. A., Jackson, K., Scase, T., Tse, T., Hampson, B., Munday, J. S., Barrs, V. R., & Beatty, J. A. (2019). A Novel Hepadnavirus is Associated with Chronic Hepatitis and Hepatocellular Carcinoma in Cats. Viruses, 11(10), 969. https://doi.org/10.3390/v11100969