Shell-Less Egg Syndrome (SES) Widespread in Western Canadian Layer Operations Is Linked to a Massachusetts (Mass) Type Infectious Bronchitis Virus (IBV) Isolate

Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Eggs and Chickens
2.3. Collection of Samples
2.4. Screening of Samples and Sequencing of IBV Partial S1 Gene
2.4.1. Tissue Homogenization
2.4.2. RNA Extraction and Complementary (c)DNA Synthesis
2.4.3. Real-Time PCR Assay
2.4.4. IBV Propagation in SPF Eggs
2.4.5. Conventional PCR Assay Targeting Partial IBV S1 Gene
2.4.6. Molecular Characterization of IBV Isolates
2.5. Reproduction of SES Using Selected Mass Type IBV Isolates
2.5.1. Pilot Experiment
2.5.2. Experimental Reproduction of SES Using 15AB-01 Mass Type IBV Isolate
2.6. Histology
2.7. IBV Antibody Titer Determination
2.8. Data Analyses
2.8.1. IBV Genome Load Quantification
2.8.2. Statistical Analysis
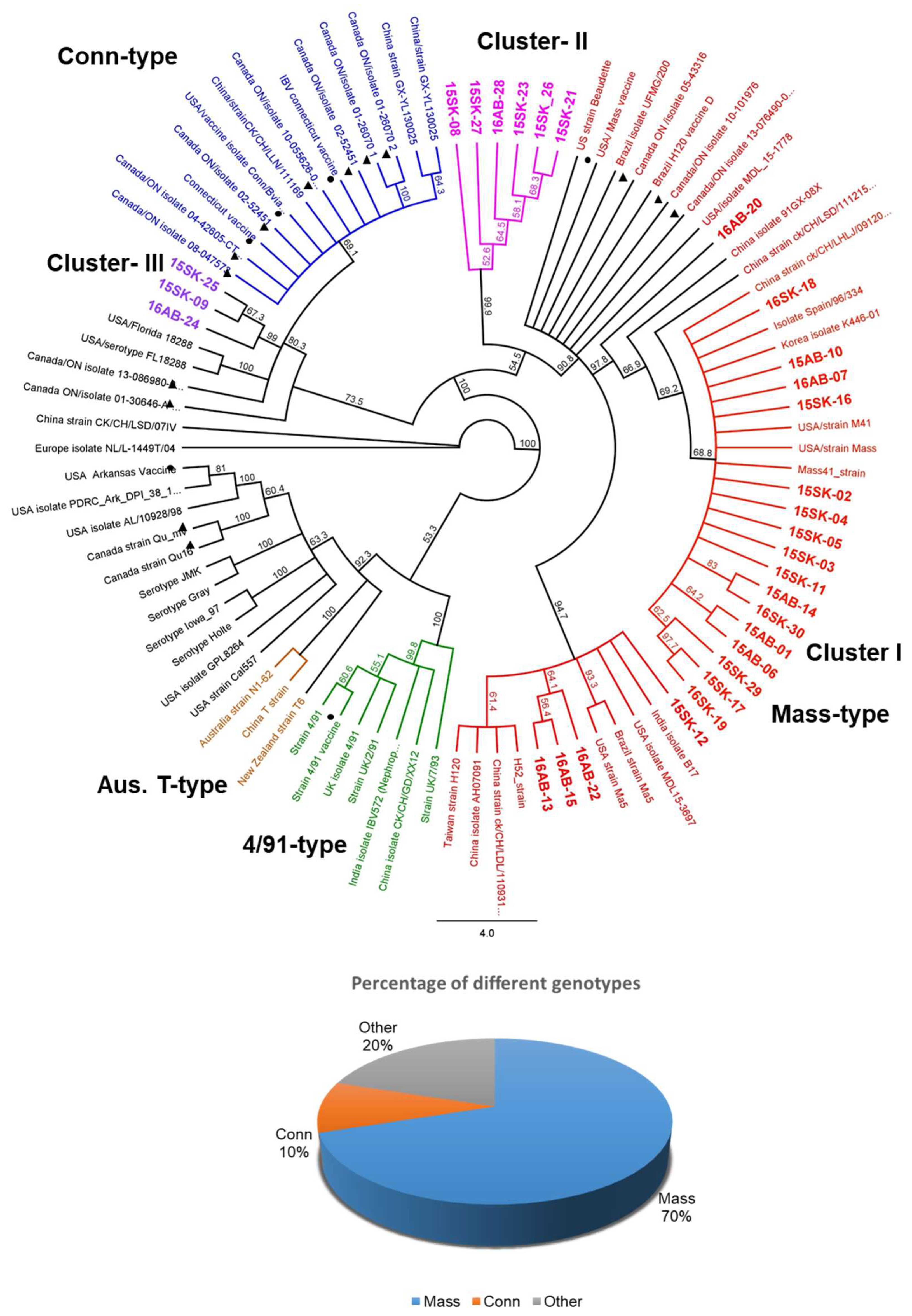
3. Results
3.1. Background of the Screened Poultry Flocks with and without a History of SES
3.2. Sample Screening for the Detection of IBV Genome
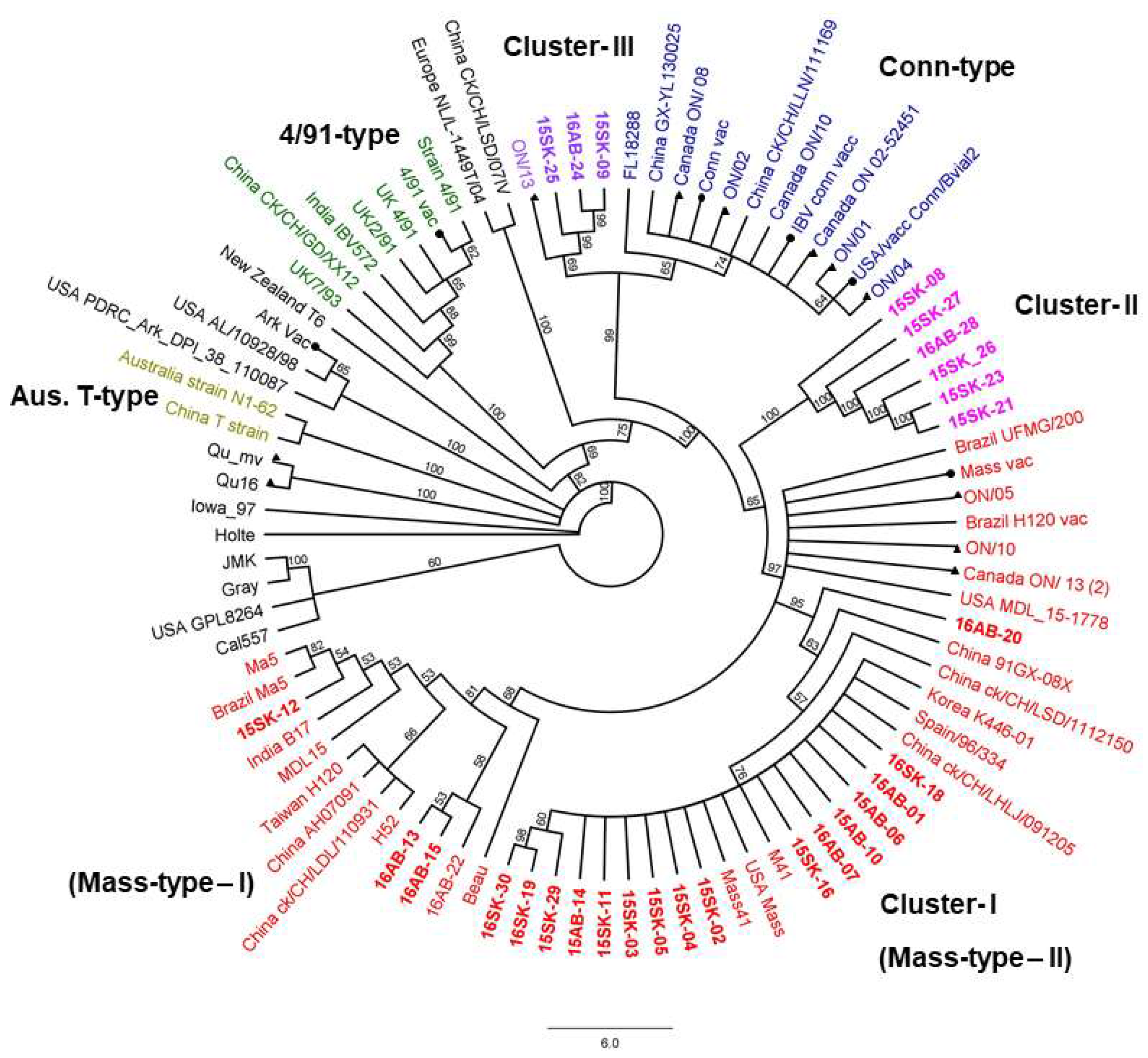
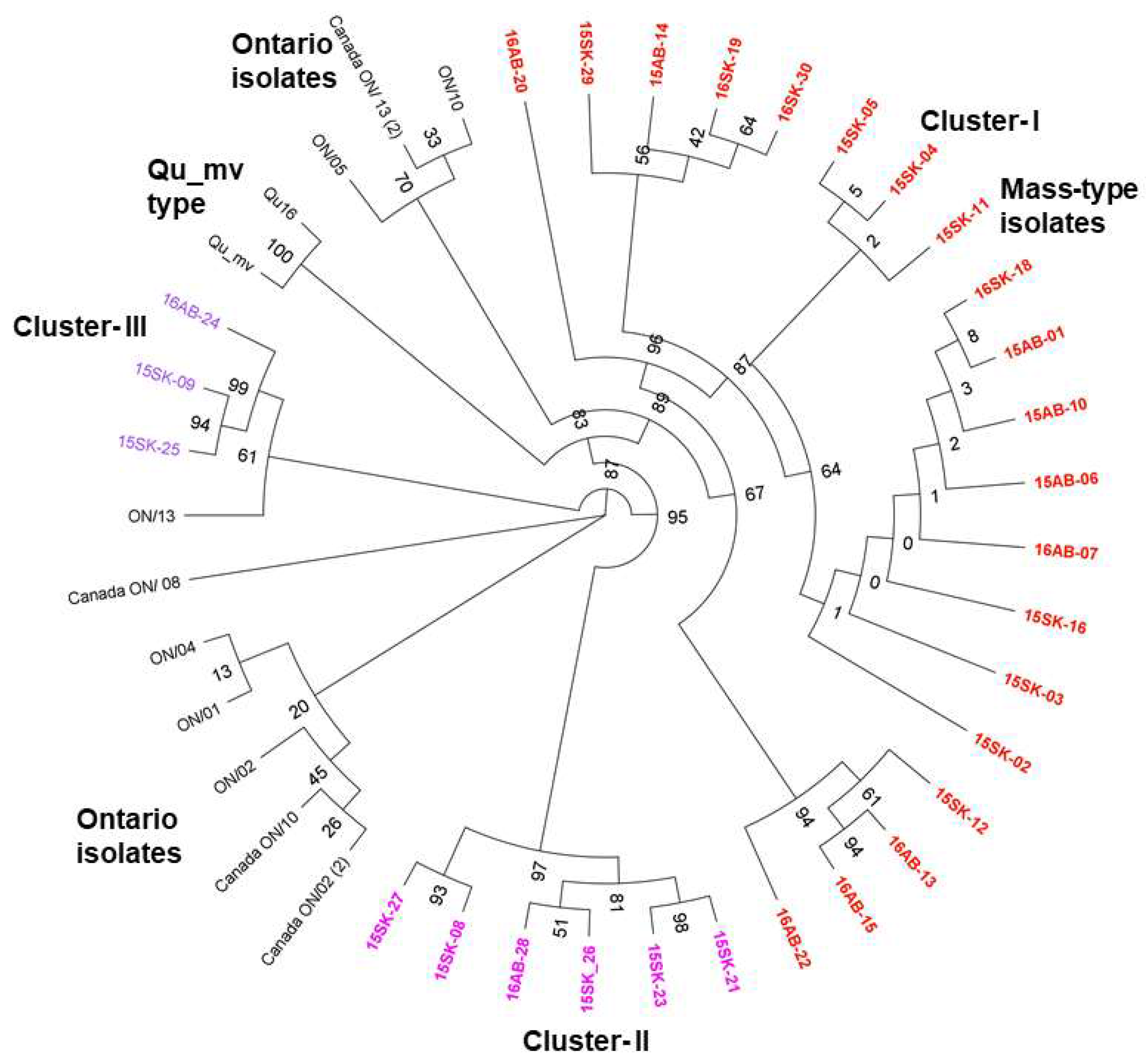
3.3. Egg Propagation, IBV S1 Gene Amplification, and Sequencing
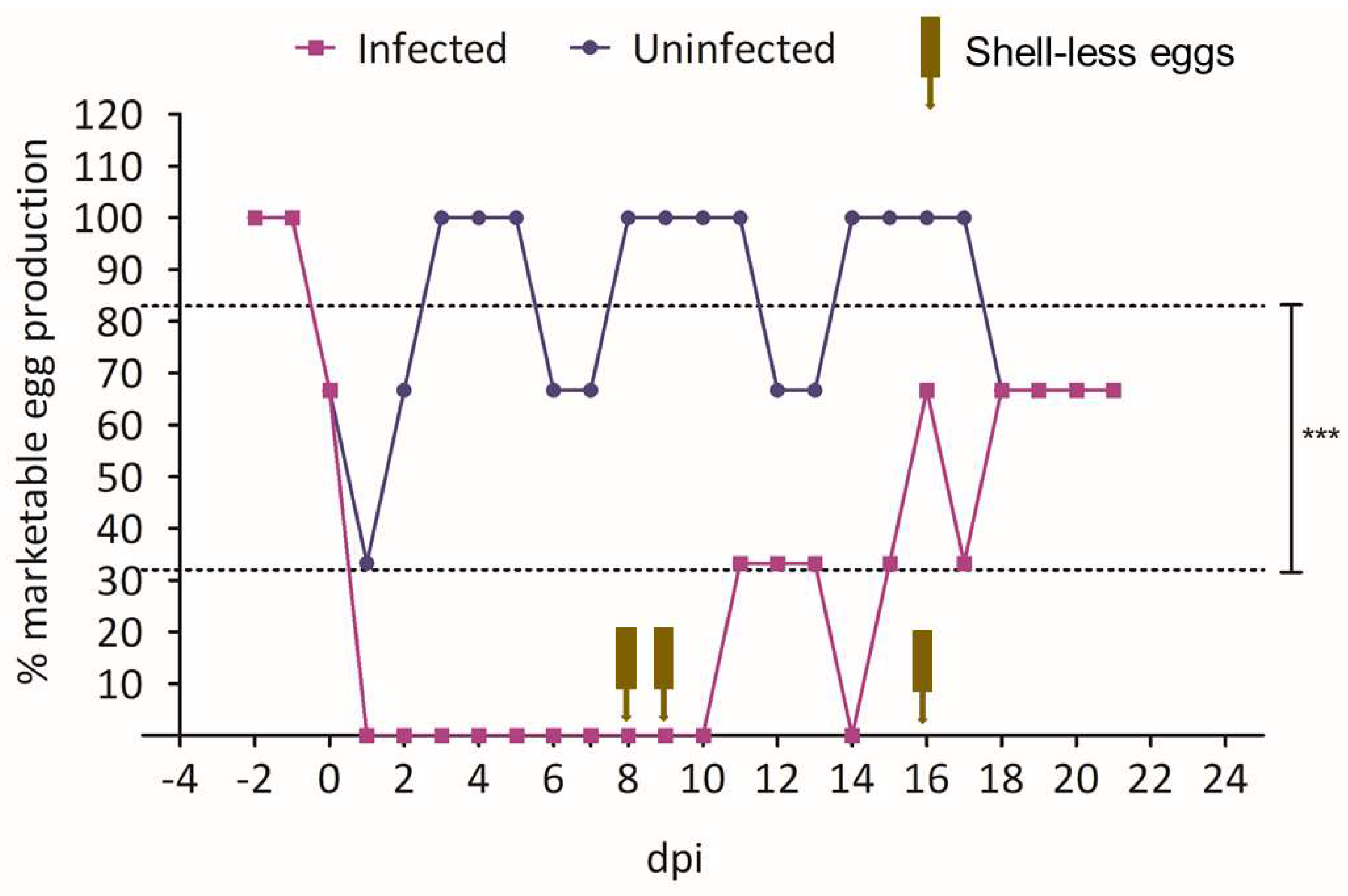
3.4. Experimental Reproduction of SES
3.4.1. Pilot Experiment
3.4.2. Egg Production Following Infection with 15AB-01 Mass Type IBV Isolate
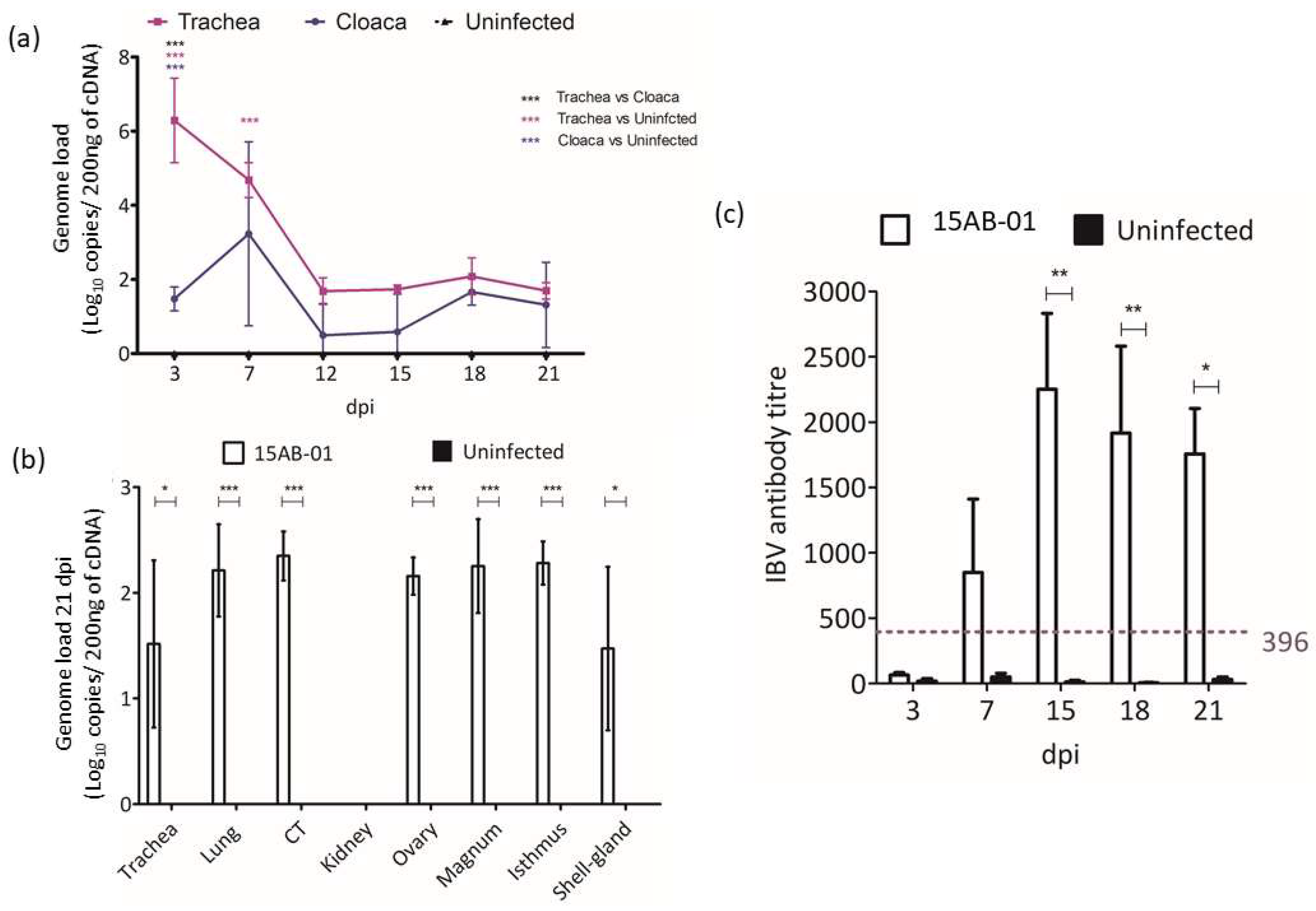
3.4.3. IBV Genome Loads Following Infection with 15AB-01 Mass Type IBV Isolate
3.4.4. Antibody Response Following Infection with 15AB-01 Mass Type IBV Isolate
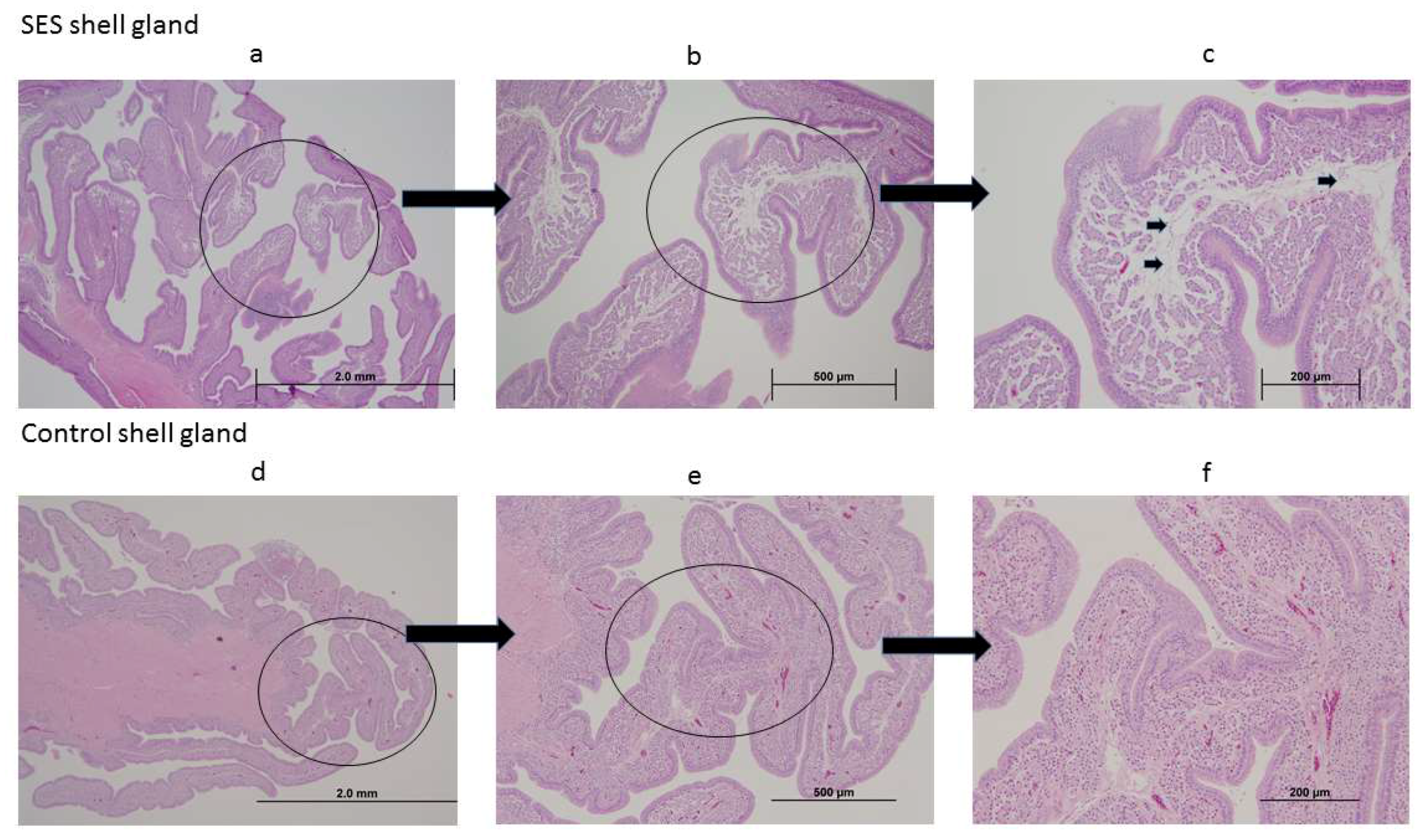
3.4.5. Histological Changes in the Shell Gland Following Infection with 15AB-01 Mass Type IBV Isolate
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sevoian, M.; Levine, P. Effects of infectious bronchitis on the reproductive tracts, egg production, and egg quality of laying chickens. Avian Dis. 1957, 1, 136–164. [Google Scholar] [CrossRef]
- McMartin, D. The pathogenicity of an infectious bronchitis virus for laying hens, with an observation on pathogenesis. Br. Vet. J. 1968, 124, 576–581. [Google Scholar] [CrossRef]
- Cook, J.; Huggins, M. Newly isolated serotypes of infectious bronchitis virus: Their role in disease. Avian Pathol. 1986, 15, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Zhou, E.M.; Dong, S.W.; Qiu, H.K.; Zhang, L.; Hu, S.B.; Zhao, F.F.; Jiang, S.J.; Sun, Y.N. Analysis of avian hepatitis E virus from chickens, China. Emerg. Infect. Dis. 2010, 16, 1469–1472. [Google Scholar] [CrossRef] [PubMed]
- Igwe, A.O.; Ihedioha, J.I.; Okoye, J.O.A. Changes in serum calcium and phosphorus levels and their relationship to egg production in laying hens infected with velogenic Newcastle disease virus. J. Appl. Anim. Res. 2018, 46, 523–528. [Google Scholar] [CrossRef]
- Qi, X.; Tan, D.; Wu, C.; Tang, C.; Li, T.; Han, X.; Wang, J.; Liu, C.; Li, R.; Wang, J. Deterioration of eggshell quality in laying hens experimentally infected with H9N2 avian influenza virus. Vet. Res. 2016, 47, 35. [Google Scholar] [CrossRef] [PubMed]
- Chousalkar, K.K.; Roberts, J.R. Ultrastructural study of infectious bronchitis virus infection in infundibulum and magnum of commercial laying hens. Vet. Microbiol. 2007, 122, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Chousalkar, K.K.; Roberts, J.R.; Reece, R. Comparative histopathology of two serotypes of infectious bronchitis virus (T and N1/88) in laying hens and cockerels. Poult. Sci. 2007, 86, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Bisgaard, M. The influence of infectious bronchitis virus on egg production, fertility, hatchability, and mortality rate in chickens (author’s transl). Nord. Vet. Med. 1976, 28, 368–376. [Google Scholar] [PubMed]
- Higashihara, M.; Hiruma, M.; Houdatsu, T.; Takai, S.; Matumoto, M. Experimental infection of laying chickens with egg-drop syndrome 1976 virus. Avian Dis. 1987, 31, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.; Imada, T.; Kawamura, H.; Taniguchi, T.; Kawakami, M. Pathogenicity and distribution of egg-drop syndrome-1976 virus (jpa-1) in inoculated laying hens. Avian Dis. 1981, 25, 642–649. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.; Imada, T.; Kawamura, H.; Taniguchi, S.; Saio, H.; Shimamatsu, K. Outbreaks of egg-drop syndrome-1976 in Japan and its etiological agent. Avian Dis. 1981, 25, 628–641. [Google Scholar] [CrossRef] [PubMed]
- Van Eck, J.H.H.; Davelaar, F.G.; Van Den Heuvel-Plesman, T.A.M.; Van Kol, N.; Kouwenhoven, B.; Guldie, F.H.M. Dropped egg production, soft-shelled and shell-less eggs associated with appearance of precipitins to adenovirus in flocks of laying fowls. Avian Pathol. 1976, 5, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Boltz, D.A.; Nakai, M.; Bahra, J.M. Avian infectious bronchitis virus: A possible cause of reduced fertility in the rooster. Avian Dis. 2004, 48, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.; Chesher, J.; Baxendale, W.; Greenwood, N.; Huggins, M.; Orbell, S. Protection of chickens against renal damage caused by a nephropathogenic infectious bronchitis virus. Avian Pathol. 2001, 30, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Muneer, M.A.; Newman, J.A.; Halvorson, D.A.; Sivanandan, V.; Coon, C.N. Effects of avian infectious bronchitis virus (Arkansas strain) on vaccinated laying chickens. Avian Dis. 1987, 31, 820–828. [Google Scholar] [CrossRef] [PubMed]
- Montassier, H.J. Molecular epidemiology and evolution of avian infectious bronchitis virus. Revista Brasileira de Ciência Avícola 2010, 12, 87–96. [Google Scholar] [CrossRef]
- Kant, A.; Koch, G.; van Roozelaar, D.J.; Kusters, J.G.; Poelwijk, F.A.J.; van der Zeijst, B.A.M. Location of antigenic sites defined by neutralizing monoclonal antibodies on the s1 avian infectious bronchitis virus glycopolypeptide. J. Gen. Virol. 1992, 73, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D.; Davis, P.J.; Cook, J.K.A. Infectious bronchitis virus: Evidence for recombination within the Massachusetts serotype. Avian Pathol. 1992, 21, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, M.W.; Lee, D.H. Different evolutionary trajectories of vaccine-controlled and non-controlled avian infectious bronchitis viruses in commercial poultry. PLoS ONE 2017, 12, e0176709. [Google Scholar] [CrossRef] [PubMed]
- Martin, E.A.; Brash, M.L.; Hoyland, S.K.; Coventry, J.M.; Sandrock, C.; Guerin, M.T.; Ojkic, D. Genotyping of infectious bronchitis viruses identified in Canada between 2000 and 2013. Avian Pathol. 2014, 43, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Ojkic, D.; Binnington, B. Phylogenetic Analysis of Ontario Infectious Bronchitis Virus Isolates. In Proceedings of the 51st Western Poultry Disease Conference, Puerto Vallarta, Mexico, 1–5 May 2002; p. 73. [Google Scholar]
- Mo, M.L.; Li, M.; Huang, B.C.; Fan, W.S.; Wei, P.; Wei, T.C.; Cheng, Q.Y.; Wei, Z.J.; Lang, Y.H. Molecular characterization of major structural protein genes of avian coronavirus infectious bronchitis virus isolates in southern China. Viruses 2013, 5, 3007–3020. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.H.; Sung, H.W.; Song, C.S.; Kwon, H.M. Sequence analysis of the s1 glycoprotein gene of infectious bronchitis viruses: Identification of a novel phylogenetic group in Korea. J. Vet. Sci. 2007, 8, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Grgic, H.; Hunter, D.B.; Hunton, P.; Nagy, E. Pathogenicity of infectious bronchitis virus isolates from Ontario chickens. Can. J. Vet. Res. 2008, 72, 403–410. [Google Scholar] [PubMed]
- Stachowiak, B.; Key, D.W.; Hunton, P.; Gillingham, S.; Nagy, É. Infectious bronchitis virus surveillance in Ontario commercial layer flocks. J. Appl. Poult. Res. 2005, 14, 141–146. [Google Scholar] [CrossRef]
- Smati, R.; Silim, A.; Guertin, C.; Henrichon, M.; Marandi, M.; Arella, M.; Merzouki, A. Molecular characterization of three new avian infectious bronchitis virus (ibv) strains isolated in Quebec. Virus Genes 2002, 25, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Kameka, A.M.; Haddadi, S.; Kim, D.S.; Cork, S.C.; Abdul-Careem, M.F. Induction of innate immune response following infectious bronchitis coronavirus infection in the respiratory tract of chickens. Virology 2014, 450–451, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. Raxml version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo-Carrasco, F.; Smith, S.A.; Stamatakis, A. Algorithms, data structures, and numerics for likelihood-based phylogenetic inference of huge trees. BMC Bioinform. 2011, 12, 470. [Google Scholar] [CrossRef] [PubMed]
- Nghia Bui, V.; Ogawa, H.; Karibe, K.; Matsuo, K.; Nguyen, T.H.; Awad, S.S.A.; Minoungou, G.L.; Saito, K.; Watanabe, Y.; et al. Surveillance of avian influenza virus in migratory water birds in eastern Hokkaido, Japan. J. Vet. Med. Sci. 2011, 73, 209–215. [Google Scholar]
- Günes, A.; Marek, A.; Grafl, B.; Berger, E.; Hess, M. Real-time PCR assay for universal detection and quantitation of all five species of fowl adenoviruses (fadv-a to fadv-e). J. Virol. Methods 2012, 183, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Wajid, A.; Wasim, M.; Yaqub, T.; Rehmani, S.F.; Bibi, T.; Mukhtar, N.; Muhammad, J.; Bacha, U.; Afridi, S.Q.; Zahid, M.N.; et al. Rapid detection/pathotyping of Newcastle disease virus isolates in clinical samples using real-time polymerase chain reaction assay. Protoc. Exch. 2014. [Google Scholar] [CrossRef]
- Marsolais, G.; Marois, P. Types of avian infectious bronchitis strains isolated in Quebec. Can. J. Comp. Med. 1982, 46, 150–153. [Google Scholar] [PubMed]
- Gelb, J.; Wolff, J.B.; Moran, C.A. Variant serotypes of infectious bronchitis virus isolated from commercial layer and broiler chickens. Avian Dis. 1991, 35, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Ojkić, D.; Martin, E.; Brash, M.; Stalker, M. Infectious Bronchitis Virus Infection in Ontario: An Update on the Changing Strains over Time. In Proceedings of the 18th International Symposium of the World Association of Veterinary Laboratory Diagnosticians, Sorrento, Italy, 7–10 June 2017. [Google Scholar]
- Jones, R.C.; Jordan, F.T.W. The exposure of day-old chicks to infectious bronchitis and the subsequent development of the oviduct. Vet. Rec. 1970, 87, 504–505. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.K.; Jackwood, M.; Jones, R.C. The long view: 40 years of infectious bronchitis research. Avian Pathol. 2012, 41, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Ignjatovic, J.; Sapats, S. Avian infectious bronchitis virus. Rev. Sci. Tech. 2000, 19, 493–508. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, T.; Yamaguchi, S.; Maeda, M.; Kawamura, H.; Horiuchi, T. Pathological changes in laying hens inoculated with the jpa-1 strain of the egg drop syndrome-1976 virus. National Inst. Anim. Health Q. 1981, 21, 83–93. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SES Status | Province of Origin | Farm ID | Real-Time PCR Results | # of Samples from Which S1 Nucleotide Sequence Obtained | Sample ID | GenBank Accession # | |
|---|---|---|---|---|---|---|---|
| # of Samples Screened | # of Samples Positive | ||||||
| History of SES | AB | 1 | 5 | 4 | 4 | 15AB-01 | MH509460 |
| 15AB-06 | MH509461 | ||||||
| 15AB-10 | MH509472 | ||||||
| 15AB-14 | MH509473 | ||||||
| 3 | 12 | 3 | 1 | 16AB-24 | MH509449 | ||
| 4 | 9 | 0 | 0 | ||||
| 7 | 12 | 1 | 1 | 16AB-07 | MH509466 | ||
| SK | 13 | 74 | 5 | 0 | |||
| 14 | 32 | 6 | 0 | ||||
| 16 | 24 | 0 | 0 | ||||
| 18 | 12 | 1 | 0 | ||||
| 20 | 27 | 3 | 1 | 15SK-12 | MH509477 | ||
| 23 | 15 | 7 | 5 | 15SK-08 | MH509448 | ||
| 15SK21 | MH509453 | ||||||
| 15SK-23 | MH509452 | ||||||
| 15SK-26 | MH509451 | ||||||
| 15SK-27 | MH509450 | ||||||
| 26 | 15 | 2 | 0 | ||||
| 27 | 30 | 3 | 0 | ||||
| Without a history of SES) | AB | 2 | 12 | 9 | 5 | 16AB-13 | MH509476 |
| 16AB-15 | MH509475 | ||||||
| 16AB-20 | MH509454 | ||||||
| 16AB-22 | MH509474 | ||||||
| 16AB-28 | MH509458 | ||||||
| 5 | 12 | 0 | 0 | ||||
| 6 | 9 | 0 | 0 | ||||
| SK | 8 | 15 | 4 | 4 | 15SK-02 | MH509465 | |
| 15SK-04 | MH509462 | ||||||
| 15SK-11 | MH509470 | ||||||
| 15SK-16 | MH509471 | ||||||
| 9 | 36 | 1 | 0 | ||||
| 10 | 48 | 3 | 0 | ||||
| 11 | 12 | 1 | 0 | ||||
| 12 | 48 | 4 | 1 | 15SK-03 | MH509464 | ||
| 15 | 22 | 2 | 0 | ||||
| 17 | 12 | 1 | 0 | ||||
| 19 | 27 | 5 | 3 | 15SK-05 | MH509463 | ||
| 15SK-18 | MH509469 | ||||||
| 15SK-19 | MH509459 | ||||||
| 21 | 12 | 2 | 1 | 15SK-29 | MH509467 | ||
| 22 | 27 | 9 | 3 | 15SK-09 | MH509457 | ||
| 15SK-17 | MH509455 | ||||||
| 15SK-25 | MH509456 | ||||||
| 24 | 15 | 1 | 0 | ||||
| 25 | 28 | 2 | 1 | 15SK-30 | MH509468 | ||
| Total | 602 | 79 | 30 | ||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amarasinghe, A.; Popowich, S.; De Silva Senapathi, U.; Abdul-Cader, M.S.; Marshall, F.; Van der Meer, F.; Cork, S.C.; Gomis, S.; Abdul-Careem, M.F. Shell-Less Egg Syndrome (SES) Widespread in Western Canadian Layer Operations Is Linked to a Massachusetts (Mass) Type Infectious Bronchitis Virus (IBV) Isolate. Viruses 2018, 10, 437. https://doi.org/10.3390/v10080437
Amarasinghe A, Popowich S, De Silva Senapathi U, Abdul-Cader MS, Marshall F, Van der Meer F, Cork SC, Gomis S, Abdul-Careem MF. Shell-Less Egg Syndrome (SES) Widespread in Western Canadian Layer Operations Is Linked to a Massachusetts (Mass) Type Infectious Bronchitis Virus (IBV) Isolate. Viruses. 2018; 10(8):437. https://doi.org/10.3390/v10080437
Chicago/Turabian StyleAmarasinghe, Aruna, Shelly Popowich, Upasama De Silva Senapathi, Mohamed Sarjoon Abdul-Cader, Frank Marshall, Frank Van der Meer, Susan C. Cork, Susantha Gomis, and Mohamed Faizal Abdul-Careem. 2018. "Shell-Less Egg Syndrome (SES) Widespread in Western Canadian Layer Operations Is Linked to a Massachusetts (Mass) Type Infectious Bronchitis Virus (IBV) Isolate" Viruses 10, no. 8: 437. https://doi.org/10.3390/v10080437
APA StyleAmarasinghe, A., Popowich, S., De Silva Senapathi, U., Abdul-Cader, M. S., Marshall, F., Van der Meer, F., Cork, S. C., Gomis, S., & Abdul-Careem, M. F. (2018). Shell-Less Egg Syndrome (SES) Widespread in Western Canadian Layer Operations Is Linked to a Massachusetts (Mass) Type Infectious Bronchitis Virus (IBV) Isolate. Viruses, 10(8), 437. https://doi.org/10.3390/v10080437