The Revisited Genome of Bacillus subtilis Bacteriophage SPP1

 and
and
Abstract
1. Introduction
2. Materials and Methods
3. Results and Discussion
3.1. Properties of the SPP1 DNA Molecule
3.2. Transcription and Translation of the SPP1 Genome
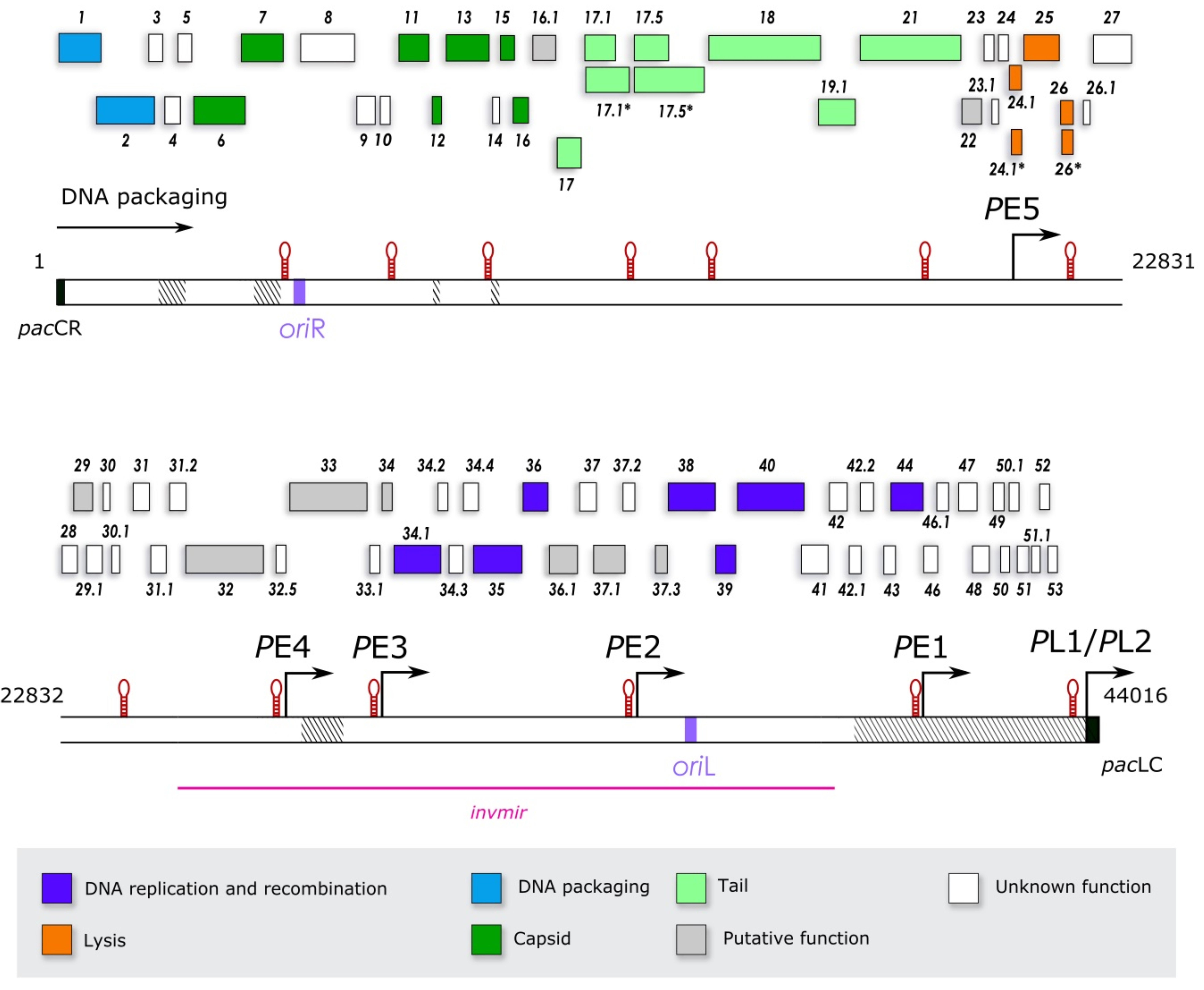
3.3. Organization and Function of the SPP1 Genes
3.4. The SPP1 Genes Set. I. Uncharacterized Early Genes
3.5. The SPP1 Genes Set. II. DNA Replication Early Genes
3.6. The SPP1 Genes Set. III. Lysis Early and Late Genes
3.7. The SPP1 Genes Set. IV. Uncharacterized Late Genes
3.8. The SPP1 Genes Set. V. Viral Particle Assembly Late Genes
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Riva, S.; Polsinelli, M.; Falaschi, A. A new phage of Bacillus subtilis with infectious DNA having separable strands. J. Mol. Biol. 1968, 35, 347–356. [Google Scholar] [CrossRef]
- Riva, S.; Polsinelli, M. Relationship between competence for transfection and for transformation. J. Virol. 1968, 2, 587–593. [Google Scholar] [PubMed]
- Spatz, H.C.; Trautner, T.A. One way to do experiments on gene conversion? Mol. Gen. Genet. 1970, 109, 84–106. [Google Scholar] [CrossRef] [PubMed]
- Spatz, H.C.; Trautner, T.A. The role of recombination in transfection of B. subtilis. Mol. Gen. Genet. 1971, 113, 174–190. [Google Scholar] [CrossRef] [PubMed]
- Deichelbohrer, I.; Alonso, J.C.; Lüder, G.; Trautner, T.A. Plasmid transduction by Bacillus subtilis bacteriophage SPP1: Effects of DNA homology between plasmid and bacteriophage. J. Bacteriol. 1985, 162, 1238–1243. [Google Scholar]
- Alonso, J.C.; Lüder, G.; Trautner, T.A. Intramolecular homologous recombination in Bacillus subtilis 168. Mol. Gen. Genet. 1992, 236, 60–64. [Google Scholar] [CrossRef]
- Behrens, B.; Lüder, G.; Behncke, M.; Trautner, T.A.; Ganesan, A.T. The genome of B. subtilis phage SPP1: Physical arrangement of phage genes. Mol. Gen. Genet. 1979, 175, 351–357. [Google Scholar] [CrossRef]
- Ratcliff, S.W.; Luh, J.; Ganesan, A.T.; Behrens, B.; Thompson, R.; Montenegro, M.A.; Morelli, G.; Trautner, T.A. The genome of Bacillus subtilis phage SPP1: The arrangement of restriction endonuclease generated fragments. Mol. Gen. Genet. 1979, 168, 165–172. [Google Scholar] [CrossRef]
- Santos, M.A.; Almeida, J.; de Lencastre, H.; Morelli, G.; Kamke, M.; Trautner, T.A. Genomic organization of the related Bacillus subtilis bacteriophages SPP1, 41c, rho 15, and SF6. J. Virol. 1986, 60, 702–707. [Google Scholar]
- Yasbin, R.E.; Frank, E. Young Transduction in Bacillus subtilis by bacteriophage SPP1. J. Virol. 1974, 14, 1343–1348. [Google Scholar]
- Ferrari, E.; Canosi, U.; Galizzi, A.; Mazza, G. Studies on Transduction Process by SPP1 Phage. J. Gen. Virol. 1978, 41, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Canosi, U.; Lüder, G.; Trautner, T.A. SPP1-mediated plasmid transduction. J. Virol. 1982, 44, 431–436. [Google Scholar] [PubMed]
- Valero-Rello, A.; López-Sanz, M.; Quevedo-Olmos, A.; Sorokin, A.; Ayora, S. Molecular mechanisms that contribute to horizontal transfer of plasmids by the bacteriophage SPP1. Front. Microbiol. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Alonso, J.C.; Lüder, G.; Trautner, T.A. Requirements for the formation of plasmid-transducing particles of Bacillus subtilis bacteriophage SPP1. EMBO J. 1986, 5, 3723–3728. [Google Scholar] [CrossRef] [PubMed]
- White, H.E.; Sherman, M.B.; Brasilès, S.; Jacquet, E.; Seavers, P.; Tavares, P.; Orlova, E.V. Capsid structure and its stability at the late stages of bacteriophage SPP1 assembly. J. Virol. 2012, 86, 6768–6777. [Google Scholar] [CrossRef] [PubMed]
- Plisson, C.; White, H.E.; Auzat, I.; Zafarani, A.; São-José, C.; Lhuillier, S.; Tavares, P.; Orlova, E.V. Structure of bacteriophage SPP1 tail reveals trigger for DNA ejection. EMBO J. 2007, 26, 3720–3728. [Google Scholar] [CrossRef] [PubMed]
- Tavares, P.; Lurz, R.; Stiege, A.; Rückert, B.; Trautner, T.A. Sequential headful packaging and fate of the cleaved DNA ends in bacteriophage SPP1. J. Mol. Biol. 1996, 264, 954–967. [Google Scholar] [CrossRef] [PubMed]
- Humphreys, G.O.; Trautner, T.A. Maturation of bacteriophage SPP1 DNA: Limited precision in the sizing of mature bacteriophage genomes. J. Virol. 1981, 37, 832–835. [Google Scholar]
- Morelli, G.; Fisseau, C.; Behrens, B.; Trautner, T.A.; Luh, J.; Ratcliff, S.W.; Allison, D.P.; Ganesan, A.T. The genome of B. subtilis phage SPP1: The topology of DNA molecules. Mol. Gen. Genet. 1979, 168, 153–164. [Google Scholar] [CrossRef]
- Tavares, P.; Santos, M.A.; Lurz, R.; Morelli, G.; De Lencastre, H.; Trautner, T.A. Identification of a gene in Bacillus subtilis bacteriophage SPP1 determining the amount of packaged DNA. J. Mol. Biol. 1992, 225, 81–92. [Google Scholar] [CrossRef]
- Baptista, C.; Santos, M.A.; São-José, C. Phage SPP1 reversible adsorption to Bacillus subtilis cell wall teichoic acids accelerates virus recognition of membrane receptor YueB. J. Bacteriol. 2008, 190, 4989–4996. [Google Scholar] [CrossRef] [PubMed]
- Baptista, C.; Barreto, H.C.; São-José, C. High levels of DegU-P activate an Esat-6-Like secretion system in Bacillus subtilis. PLoS ONE 2013, 8, e67840. [Google Scholar] [CrossRef] [PubMed]
- São-José, C.; Lhuillier, S.; Lurz, R.; Melki, R.; Lepault, J.; Santos, M.A.; Tavares, P. The ectodomain of the viral receptor YueB forms a fiber that triggers ejection of bacteriophage SPP1 DNA. J. Biol. Chem. 2006, 281, 11464–11470. [Google Scholar] [CrossRef] [PubMed]
- Jakutyte, L.; Baptista, C.; São-José, C.; Daugelavičius, R.; Carballido-López, R.; Tavares, P. Bacteriophage infection in rod-shaped gram-positive bacteria: Evidence for a preferential polar route for phage SPP1 entry in Bacillus subtilis. J. Bacteriol. 2011, 193, 4893–4903. [Google Scholar] [CrossRef] [PubMed]
- Jakutyte, L.; Lurz, R.; Baptista, C.; Carballido-López, R.; São-José, C.; Tavares, P.; Daugelavičius, R. First steps of bacteriophage SPP1 entry into Bacillus subtilis. Virology 2012, 422, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Alonso, J.C.; Lüder, G.; Stiege, A.C.; Chai, S.; Weise, F.; Trautner, T.A. The complete nucleotide sequence and functional organization of Bacillus subtilis bacteriophage SPP1. Gene 1997, 204, 201–212. [Google Scholar] [CrossRef]
- Burger, K. Biochemische und genetische Untersuchungen zur DNA-Replikation des B. subtilis-Bakteriophagen SPP1. Ph.D. Thesis, Freien Universität, Berlin, Germany, 1978. [Google Scholar]
- Burger, K.J.; Trautner, T.A. Specific labelling of replicating SPP1 DNA: Analysis of viral DNA synthesis and identification of phage DNA-genes. Mol. Gen. Genet. 1978, 166, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Mastromei, G.; Riva, S.; Fietta, A.; Pagani, L. SPP1 DNA replicative forms: Growth of phage SPP1 in Bacillus subtilis mutants temperature-sensitive in DNA synthesis. Mol. Gen. Genet. 1978, 167, 157–164. [Google Scholar] [CrossRef]
- Lo Piano, A.; Martínez-Jiménez, M.I.; Zecchi, L.; Ayora, S. Recombination-dependent concatemeric viral DNA replication. Virus Res. 2011, 160, 1–14. [Google Scholar] [CrossRef]
- Zecchi, L.; Lo Piano, A.; Suzuki, Y.; Cañas, C.; Takeyasu, K.; Ayora, S. Characterization of the Holliday Junction resolving enzyme encoded by the Bacillus subtilis bacteriophage SPP1. PLoS ONE 2012, 7, e48440. [Google Scholar] [CrossRef]
- Auzat, I.; Petitpas, I.; Lurz, R.; Weise, F.; Tavares, P. A touch of glue to complete bacteriophage assembly: The tail-to-head joining protein (THJP) family. Mol. Microbiol. 2014, 91, 1164–1178. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, S.; São-José, C. Probing the function of the two holin-like proteins of bacteriophage SPP1. Virology 2017, 500, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, E.L.; Auger, H.; Jaszczyszyn, Y.; Thermes, C. Ten years of next-generation sequencing technology. Trends Genet. 2014, 30, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Djacem, K.; Tavares, P.; Oliveira, L. Bacteriophage SPP1 pac cleavage: A precise cut without sequence specificity requirement. J. Mol. Biol. 2017, 429, 1381–1395. [Google Scholar] [CrossRef] [PubMed]
- Solovyev, V.; Salamov, A. Automatic annotation of microbial genomes and metagenomic sequences. In Metagenomics and Its Applications in Agriculture, Biomedicine, and Environmental Studies; Li, R.W., Ed.; Nova Science Publisher′s: Hauppauge, NY, USA, 2011; pp. 61–78. ISBN 978-1-61668-682-6. [Google Scholar]
- Moran, C.P.; Lang, N.; LeGrice, S.F.J.; Lee, G.; Stephens, M.; Sonenshein, A.L.; Pero, J.; Losick, R. Nucleotide sequences that signal the initiation of transcription and translation in Bacillus subtilis. Mol. Gen. Genet. 1982, 186, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Madden, T. The BLAST Sequence Analysis Tool. Available online: https://www.ncbi.nlm.nih.gov/books/NBK153387/ (accessed on 22 August 2018).
- Zimmermann, L.; Stephens, A.; Nam, S.-Z.; Rau, D.; Kübler, J.; Lozajic, M.; Gabler, F.; Söding, J.; Lupas, A.N.; Alva, V. A completely reimplemented MPI Bioinformatics toolkit with a new HHpred server at its core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef] [PubMed]
- Stüber, D.; Morelli, G.; Bujard, H.; Montenegro, M.A.; Trautner, T.A. Promoter sites in the genome of B. subtilis phage SPP1. Mol. Gen. Genet. 1981, 181, 518–521. [Google Scholar] [CrossRef] [PubMed]
- Tailor, R.; Bensi, G.; Morelli, G.; Canosi, U.; Trautner, T.A. The genome of Bacillus subtilis phage SPP1: structure of an early promoter. J. Gen. Microbiol. 1985, 131, 1259–1262. [Google Scholar] [CrossRef]
- Gautheret, D.; Lambert, A. Direct RNA motif definition and identification from multiple sequence alignments using secondary structure profiles. J. Mol. Biol. 2001, 313, 1003–1011. [Google Scholar] [CrossRef] [PubMed]
- Hofacker, I.L.; Fontana, W.; Stadler, P.F.; Bonhoeffer, L.S.; Tacker, M.; Schuster, P. Fast folding and comparison of RNA secondary structures. Monatshefte für Chemie 1994, 125, 167–188. [Google Scholar] [CrossRef]
- Lesnik, E.A.; Sampath, R.; Levene, H.B.; Henderson, T.J.; McNeil, J.A.; Ecker, D.J. Prediction of rho-independent transcriptional terminators in Escherichia coli. Nucleic Acids Res. 2001, 29, 3583–3594. [Google Scholar] [CrossRef] [PubMed]
- Macke, T.J.; Ecker, D.J.; Gutell, R.R.; Gautheret, D.; Case, D.A.; Sampath, R. RNAMotif, an RNA secondary structure definition and search algorithm. Nucleic Acids Res. 2001, 29, 4724–4735. [Google Scholar] [CrossRef] [PubMed]
- Moszer, I.; Rocha, E.P.C.; Danchin, A. Codon usage and lateral gene transfer in Bacillus subtilis. Curr. Opin. Microbiol. 1999, 2, 524–528. [Google Scholar] [CrossRef]
- Gupta, S.K.; Ghosh, T.C. CUCG: A non-redundant codon usage database from complete genomes. Curr. Sci. 2000, 78, 28–29. [Google Scholar]
- Kunst, F.; Ogasawara, N.; Moszer, I.; Albertini, A.M.; Alloni, G.; Azevedo, V.; Bertero, M.G.; Bessières, P.; Bolotin, A.; Borchert, S.; et al. The complete genome sequence of the gram-positive bacterium Bacillus subtilis. Nature 1997, 390, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Riva, S.C. Asymetric transcription of B. subtilis phage SPP1 DNA in vitro. Biochem. Biophys. Res. Commun. 1969, 34, 824–830. [Google Scholar] [CrossRef]
- Chenciner, N.; Milanesi, G. Restriction fragment analysis of bacteriophage SPP1 in vitro transcription by host RNA polymerase. J. Virol. 1978, 28, 95–105. [Google Scholar] [PubMed]
- Montenegro, M.A.; Trautner, T.A. In vivo transcription of Bacillus subtilis bacteriophage SPP1. Mol. Gen. Genet. 1981, 181, 512–517. [Google Scholar] [CrossRef]
- Desmyter, A.; Reeve, J.N.; Morelli, G.; Trautner, T.A. Inversion and deletion mutants in Bacillus subtilis bacteriophage SPP1 as a consequence of cloning. Mol. Gen. Genet. 1985, 198, 537–539. [Google Scholar] [CrossRef] [PubMed]
- Milanesi, G.; Cassani, G. Transcription after bacteriophage SPP1 infection in Bacillus subtilis. J. Virol. 1972, 10, 187–192. [Google Scholar] [PubMed]
- Chai, S.; Szepan, U.; Liider, G.; Trautner, T.A.; Alonso, J.C. Sequence analysis of the left end of the Bacillus subtilis bacteriophage SPP1 genome. Gene 1993, 129, 41–49. [Google Scholar] [CrossRef]
- Pedré, X.; Weise, F.; Chai, S.; Lüder, G.; Alonso, J.C. Analysis of cis and trans acting elements required for the initiation of DNA replication in the Bacillus subtilis bacteriophage SPP1. J. Mol. Biol. 1994, 236, 1324–1340. [Google Scholar] [CrossRef]
- Chai, S.; Bravo, A.; Lüder, G.; Nedlin, A.; Trautner, T.A.; Alonso, J.C. Molecular analysis of the Bacillus subtilis bacteriophage SPP1 region encompassing genes 1 to 6. The products of gene 1 and gene 2 are required for pac cleavage. J. Mol. Biol. 1992, 224, 87–102. [Google Scholar] [CrossRef]
- Pope, W.H.; Jacobs-Sera, D.; Russell, D.A.; Rubin, D.H.F.; Kajee, A.; Msibi, Z.N.P.; Larsen, M.H.; Jacobs, W.R.; Lawrence, J.G.; Hendrix, R.W.; et al. Genomics and proteomics of mycobacteriophage patience, an accidental tourist in the Mycobacterium neighborhood. MBio 2014, 5, e02145. [Google Scholar] [CrossRef] [PubMed]
- Erez, Z.; Steinberger-Levy, I.; Shamir, M.; Doron, S.; Stokar-Avihail, A.; Peleg, Y.; Melamed, S.; Leavitt, A.; Savidor, A.; Albeck, S.; et al. Communication between viruses guides lysis-lysogeny decisions. Nature 2017, 541, 488–493. [Google Scholar] [CrossRef] [PubMed]
- Lurz, R.; Orlova, E.V.; Günther, D.; Dube, P.; Dröge, A.; Weise, F.; van Heel, M.; Tavares, P. Structural organisation of the head-to-tail interface of a bacterial virus. J. Mol. Biol. 2001, 310, 1027–1037. [Google Scholar] [CrossRef] [PubMed]
- Sprengel, R.; Reiss, B.; Schaller, H. Translationally coupled initiation of protein synthesis in Bacillus subtilis. Nucleic Acids Res. 1985, 13, 893–909. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Campbell, A.; Karlin, S. Correlations between Shine-Dalgarno sequences and gene features such as predicted expression levels and operon structures. J. Bacteriol. 2002, 184, 5733–5745. [Google Scholar] [CrossRef]
- Chai, S.; Lurz, R.; Alonso, J.C. The small subunit of the terminase enzyme of Bacillus subtilis bacteriophage SPP1 forms a specialized nucleoprotein complex with the packaging initiation region. J. Mol. Biol. 1995, 252, 386–398. [Google Scholar] [CrossRef]
- Gual, A.; Alonso, J.C. Characterization of the small subunit of the terminase enzyme of the Bacillus subtilis bacteriophage SPP1. Virology 1998, 242, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Büttner, C.R.; Chechik, M.; Ortiz-Lombardía, M.; Smits, C.; Ebong, I.-O.; Chechik, V.; Jeschke, G.; Dykeman, E.; Benini, S.; Robinson, C.V.; et al. Structural basis for DNA recognition and loading into a viral packaging motor. Proc. Natl. Acad. Sci. USA 2012, 109, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Greive, S.J.; Fung, H.K.H.; Chechik, M.; Jenkins, H.T.; Weitzel, S.E.; Aguiar, P.M.; Brentnall, A.S.; Glousieau, M.; Gladyshev, G.V.; Potts, J.R.; et al. DNA recognition for virus assembly through multiple sequence-independent interactions with a helix-turn-helix motif. Nucleic Acids Res. 2016, 44, 776–789. [Google Scholar] [CrossRef] [PubMed]
- Camacho, A.G.; Gual, A.; Lurz, R.; Tavares, P.; Alonso, J.C. Bacillus subtilis bacteriophage SPP1 DNA packaging motor requires terminase and portal proteins. J. Biol. Chem. 2003, 278, 23251–23259. [Google Scholar] [CrossRef]
- Chai, S.; Alonso, J.C. Distamycin-induced inhibition of formation of a nucleoprotein complex between the terminase small subunit G1P and the non-encapsidated end (pacL site) of Bacillus subtilis bacteriophage SPP1. Nucleic Acids Res. 1996, 24, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Chai, S.; Szepan, U.; Alonso, J.C. Bacillus subtilis bacteriophage SPP1 terminase has a dual activity: It is required for the packaging initiation and represses its own synthesis. Gene 1997, 184, 251–256. [Google Scholar] [CrossRef]
- Gual, A.; Camacho, A.G.; Alonso, J.C. Functional analysis of the terminase large subunit, G2P, of Bacillus subtilis Bacteriophage SPP1. J. Biol. Chem. 2000, 275, 35311–35319. [Google Scholar] [CrossRef]
- Smits, C.; Chechik, M.; Kovalevskiy, O.V.; Shevtsov, M.B.; Foster, A.W.; Alonso, J.C.; Antson, A.A. Structural basis for the nuclease activity of a bacteriophage large terminase. EMBO Rep. 2009, 10, 592–598. [Google Scholar] [CrossRef]
- Cornilleau, C.; Atmane, N.; Jacquet, E.; Smits, C.; Alonso, J.C.; Tavares, P.; Oliveira, L. The nuclease domain of the SPP1 packaging motor coordinates DNA cleavage and encapsidation. Nucleic Acids Res. 2013, 41, 340–354. [Google Scholar] [CrossRef]
- Oliveira, L.; Henriques, A.O.; Tavares, P. Modulation of the viral ATPase activity by the portal protein correlates with DNA packaging efficiency. J. Biol. Chem. 2006, 281, 21914–21923. [Google Scholar] [CrossRef]
- Oliveira, L.; Cuervo, A.; Tavares, P. Direct interaction of the bacteriophage SPP1 packaging ATPase with the portal protein. J. Biol. Chem. 2010, 285, 7366–7373. [Google Scholar] [CrossRef] [PubMed]
- Dube, P.; Tavares, P.; Lurz, R.; Van Heel, M. The portal protein of bacteriophage SPP1: a DNA pump with 13-fold symmetry. EMBO J. 1993, 1, 1303–1309. [Google Scholar] [CrossRef]
- Jekow, P.; Behlke, J.; Tichelaar, W.; Lurz, R.; Regalla, M.; Hinrichs, W.; Tavares, P. Effect of the ionic environment on the molecular structure of bacteriophage SPP1 portal protein. Eur. J. Biochem. 1999, 264, 724–735. [Google Scholar] [CrossRef] [PubMed]
- Orlova, E.V.; Dube, P.; Beckmann, E.; Zemlin, F.; Lurz, R.; Trautner, T.A.; Tavares, P.; Van Heel, M. Structure of the 13-fold symmetric portal protein of bacteriophage SPP1. Nat. Struct. Biol. 1999, 6, 842–846. [Google Scholar] [CrossRef] [PubMed]
- Orlova, E.V.; Gowen, B.; Dröge, A.; Stiege, A.; Weise, F.; Lurz, R.; van Heel, M.; Tavares, P. Structure of a viral DNA gatekeeper at 10 Å resolution by cryo-electron microscopy. EMBO J. 2003, 22, 1255–1262. [Google Scholar] [CrossRef] [PubMed]
- Isidro, A.; Henriques, A.O.; Tavares, P. The portal protein plays essential roles at different steps of the SPP1 DNA packaging process. Virology 2004, 322, 253–263. [Google Scholar] [CrossRef]
- Isidro, A.; Santos, M.A.; Henriques, A.O.; Tavares, P. The high-resolution functional map of bacteriophage SPP1 portal protein. Mol. Microbiol. 2004, 51, 949–962. [Google Scholar] [CrossRef]
- Chaban, Y.; Lurz, R.; Brasilès, S.; Cornilleau, C.; Karreman, M.; Zinn-Justin, S.; Tavares, P.; Orlova, E.V. Structural rearrangements in the phage head-to-tail interface during assembly and infection. Proc. Natl. Acad. Sci. USA 2015, 112, 7009–7014. [Google Scholar] [CrossRef]
- Cuervo, A.; Vaney, M.-C.; Antson, A.A.; Tavares, P.; Oliveira, L. Structural rearrangements between portal protein subunits are essential for viral DNA translocation. J. Biol. Chem. 2007, 282, 18907–18913. [Google Scholar] [CrossRef]
- Vinga, I.; Dröge, A.; Stiege, A.C.; Lurz, R.; Santos, M.A.; Daugelavičius, R.; Tavares, P. The minor capsid protein gp7 of bacteriophage SPP1 is required for efficient infection of Bacillus subtilis. Mol. Microbiol. 2006, 61, 1609–1621. [Google Scholar] [CrossRef]
- Stiege, A.C.; Isidro, A.; Dröge, A.; Tavares, P. Specific targeting of a DNA-binding protein to the SPP1 procapsid by interaction with the portal oligomer. Mol. Microbiol. 2003, 49, 1201–1212. [Google Scholar] [CrossRef] [PubMed]
- Becker, B.; de la Fuente, N.; Gassel, M.; Günther, D.; Tavares, P.; Lurz, R.; Trautner, T.A.; Alonso, J.C. Head morphogenesis genes of the Bacillus subtilis bacteriophage SPP1. J. Mol. Biol. 1997, 268, 822–839. [Google Scholar] [CrossRef] [PubMed]
- Dröge, A.; Santos, M.A.; Stiege, A.C.; Alonso, J.C.; Lurz, R.; Trautner, T.A.; Tavares, P. Shape and DNA packaging activity of bacteriophage SPP1 procapsid: protein components and interactions during assembly. J. Mol. Biol. 2000, 296, 117–132. [Google Scholar] [CrossRef] [PubMed]
- Poh, S.L.; el Khadali, F.; Berrier, C.; Lurz, R.; Melki, R.; Tavares, P. Oligomerization of the SPP1 scaffolding Protein. J. Mol. Biol. 2008, 378, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Zairi, M.; Stiege, A.C.; Nhiri, N.; Jacquet, E.; Tavares, P. The collagen-like protein gp12 is a temperature-dependent reversible binder of SPP1 viral capsids. J. Biol. Chem. 2014, 289, 27169–27181. [Google Scholar] [CrossRef] [PubMed]
- Lhuillier, S.; Gallopin, M.; Gilquin, B.; Brasilès, S.; Lancelot, N.; Letellier, G.; Gilles, M.; Dethan, G.; Orlova, E.V.; Couprie, J.; et al. Structure of bacteriophage SPP1 head-to-tail connection reveals mechanism for viral DNA gating. Proc. Natl. Acad. Sci. USA 2009, 106, 8507–8512. [Google Scholar] [CrossRef] [PubMed]
- Chagot, B.; Auzat, I.; Gallopin, M.; Petitpas, I.; Gilquin, B.; Tavares, P.; Zinn-Justin, S. Solution structure of gp17 from the Siphoviridae bacteriophage SPP1: Insights into its role in virion assembly. Proteins Struct. Funct. Bioinform. 2012, 80, 319–326. [Google Scholar] [CrossRef]
- Auzat, I.; Dröge, A.; Weise, F.; Lurz, R.; Tavares, P. Origin and function of the two major tail proteins of bacteriophage SPP1. Mol. Microbiol. 2008, 70, 557–569. [Google Scholar] [CrossRef]
- Langlois, C.; Ramboarina, S.; Cukkemane, A.; Auzat, I.; Chagot, B.; Gilquin, B.; Ignatiou, A.; Petitpas, I.; Kasotakis, E.; Paternostre, M.; et al. Bacteriophage SPP1 tail tube protein self-assembles into β-structure-rich tubes. J. Biol. Chem. 2015, 290, 3836–3849. [Google Scholar] [CrossRef]
- Goulet, A.; Lai-Kee-Him, J.; Veesler, D.; Auzat, I.; Robin, G.; Shepherd, D.A.; Ashcroft, A.E.; Richard, E.; Lichière, J.; Tavares, P.; et al. The opening of the SPP1 bacteriophage tail, a prevalent mechanism in Gram-positive-infecting siphophages. J. Biol. Chem. 2011, 286, 25397–25405. [Google Scholar] [CrossRef]
- Veesler, D.; Robin, G.; Lichière, J.; Auzat, I.; Tavares, P.; Bron, P.; Campanacci, V.; Cambillau, C. Crystal structure of bacteriophage SPP1 distal tail protein (gp19.1): A baseplate hub paradigm in gram-positive infecting phages. J. Biol. Chem. 2010, 285, 36666–36673. [Google Scholar] [CrossRef] [PubMed]
- Vinga, I.; Baptista, C.; Auzat, I.; Petipas, I.; Lurz, R.; Tavares, P.; Santos, M.A.; São-José, C. Role of bacteriophage SPP1 tail spike protein gp21 on host cell receptor binding and trigger of phage DNA ejection. Mol. Microbiol. 2012, 83, 289–303. [Google Scholar] [CrossRef] [PubMed]
- Veesler, D.; Blangy, S.; Spinelli, S.; Tavares, P.; Campanacci, V.; Cambillau, C. Crystal structure of Bacillus subtilis SPP1 phage gp22 shares fold similarity with a domain of lactococcal phage p2 RBP. Protein Sci. 2010, 19, 1439–1443. [Google Scholar] [CrossRef] [PubMed]
- Veesler, D.; Blangy, S.; Lichière, J.; Ortiz-Lombardía, M.; Tavares, P.; Campanacci, V.; Cambillau, C. Crystal structure of Bacillus subtilis SPP1 phage gp23.1, a putative chaperone. Protein Sci. 2010, 19, 1812–1816. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, S.; São-José, C. More than a hole: The holin lethal function may be required to fully sensitize bacteria to the lytic action of canonical endolysins. Mol. Microbiol. 2016, 102, 92–106. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Jiménez, M.I.; Alonso, J.C.; Ayora, S. Bacillus subtilis bacteriophage SPP1-encoded gene 34.1 product is a recombination-dependent DNA replication protein. J. Mol. Biol. 2005, 351, 1007–1019. [Google Scholar] [CrossRef]
- Ayora, S.; Missich, R.; Mesa, P.; Lurz, R.; Yang, S.; Egelman, E.H.; Alonso, J.C. Homologous-pairing activity of the Bacillus subtilis bacteriophage SPP1 replication protein G35P. J. Biol. Chem. 2002, 277, 35969–35979. [Google Scholar] [CrossRef]
- Seco, E.M.; Zinder, J.C.; Manhart, C.M.; Lo Piano, A.; McHenry, C.S.; Ayora, S. Bacteriophage SPP1 DNA replication strategies promote viral and disable host replication in vitro. Nucleic Acids Res. 2013, 41, 1711–1721. [Google Scholar] [CrossRef]
- Missich, R.; Weise, F.; Chai, S.; Lurz, R.; Pedré, X.; Alonso, J.C. The replisome organizer (G38P) of Bacillus subtilis bacteriophage SPP1 forms specialized nucleoprotein complexes with two discrete distant regions of the SPP1 genome. J. Mol. Biol. 1997, 270, 50–64. [Google Scholar] [CrossRef]
- Ayora, S.; Stasiak, A.; Alonso, J.C. The Bacillus subtilis bacteriophage SPP1 G39P delivers and activates the G40P DNA helicase upon interacting with the G38P-bound replication origin. J. Mol. Biol. 1999, 288, 71–85. [Google Scholar] [CrossRef]
- Bailey, S.; Sedelnikova, S.E.; Mesa, P.; Ayora, S.; Waltho, J.P.; Ashcroft, A.E.; Baron, A.J.; Alonso, J.C.; Rafferty, J.B. Structural analysis of Bacillus subtilis SPP1 phage helicase loader protein G39P. J. Biol. Chem. 2003, 278, 15304–15312. [Google Scholar] [CrossRef] [PubMed]
- Ayora, S.; Weise, F.; Mesa, P.; Stasiak, A.; Alonso, J.C. Bacillus subtilis bacteriophage SPP1 hexameric DNA helicase, G40P, interacts with forked DNA. Nucleic Acids Res. 2002, 30, 2280–2289. [Google Scholar] [CrossRef] [PubMed]
- Mesa, P.; Alonso, J.C.; Ayora, S. Bacillus subtilis Bacteriophage SPP1 G40P helicase lacking the N-terminal domain unwinds DNA bidirectionally. J. Mol. Biol. 2006, 357, 1077–1088. [Google Scholar] [CrossRef] [PubMed]
- Ayora, S.; Langer, U.; Alonso, J.C. Bacillus subtilis DnaG primase stabilises the bacteriophage SPP1 G40P helicase-ssDNA complex. FEBS Lett. 1998, 439, 59–62. [Google Scholar] [CrossRef]
- Wang, G.; Klein, M.G.; Tokonzaba, E.; Zhang, Y.; Holden, L.G.; Chen, X.S. The structure of a DnaB-family replicative helicase and its interactions with primase. Nat. Struct. Mol. Biol. 2008, 15, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Jiménez, M.I.; Mesa, P.; Alonso, J.C. Bacillus subtilis tau subunit of DNA polymerase III interacts with bacteriophage SPP1 replicative DNA helicase G40P. Nucleic Acids Res. 2002, 30, 5056–5064. [Google Scholar] [CrossRef] [PubMed]
- Rocha, E.P.C.; Danchin, A.; Viari, A. Translation in Bacillus subtilis: Roles and trends of initiation and termination, insights from a genome analysis. Nucleic Acids Res. 1999, 27, 3567–3576. [Google Scholar] [CrossRef] [PubMed]
- Lovett, P.S.; Ambulos, N.P.; Mulbry, W.; Noguchi, N.; Rogers, E.J. UGA can be decoded as tryptophan at low efficiency in Bacillus subtilis. J. Bacteriol. 1991, 173, 1810–1812. [Google Scholar] [CrossRef]
- Xu, J.; Hendrix, R.W.; Duda, R.L. Conserved translational frameshift in dsDNA bacteriophage tail assembly genes. Mol. Cell 2004, 16, 11–21. [Google Scholar] [CrossRef]
- Xu, J.; Hendrix, R.W.; Duda, R.L. A balanced ratio of proteins from gene G and frameshift-extended gene GT is required for phage Lambda tail assembly. J. Mol. Biol. 2013, 425, 3476–3487. [Google Scholar] [CrossRef]
- Xu, J.; Hendrix, R.W.; Duda, R.L. Chaperone–protein interactions that mediate assembly of the bacteriophage lambda tail to the correct length. J. Mol. Biol. 2014, 426, 1004–1018. [Google Scholar] [CrossRef] [PubMed]
- Juhala, R.J.; Ford, M.E.; Duda, R.L.; Youlton, A.; Hatfull, G.F.; Hendrix, R.W. Genomic sequences of bacteriophages HK97 and HK022: Pervasive genetic mosaicism in the lambdoid bacteriophages. J. Mol. Biol. 2000, 299, 27–51. [Google Scholar] [CrossRef] [PubMed]
- Hendrix, R.W.; Lawrence, J.G.; Hatfull, G.F.; Casjens, S. The origins and ongoing evolution of viruses. Trends Microbiol. 2000, 8, 504–508. [Google Scholar] [CrossRef]
- Cumby, N.; Davidson, A.R.; Maxwell, K.L. The moron comes of age. Bacteriophage 2012, 2, e23146. [Google Scholar] [CrossRef]
- Chai, S.; Kruft, V.; Alonso, J.C. Analysis of the Bacillus subtilis bacteriophages SPP1 and SF6 gene 1 product: a protein involved in the initiation of headful packaging. Virology 1994, 202, 930–939. [Google Scholar] [CrossRef]
- Umene, K.; Shiraishi, A. Complete nucleotide sequence of Bacillus subtilis (natto) bacteriophage PM1, a phage associated with disruption of food production. Virus Genes 2013, 46, 524–534. [Google Scholar] [CrossRef] [PubMed]
- Marks, T.J.; Hamilton, P.T. Characterization of a thermophilic bacteriophage of Geobacillus kaustophilus. Arch. Virol. 2014, 159, 2771–2775. [Google Scholar] [CrossRef] [PubMed]
- Botstein, D. A theory of modular evolution for bacteriophages. Ann. N. Y. Acad. Sci. 1980, 354, 484–491. [Google Scholar] [CrossRef]
- Hendrix, R.W.; Smith, M.C.; Burns, R.N.; Ford, M.E.; Hatfull, G.F. Evolutionary relationships among diverse bacteriophages and prophages: All the world′s a phage. Proc. Natl. Acad. Sci. USA 1999, 96, 2192–2197. [Google Scholar] [CrossRef]
- Alonso, J.C.; Tavares, P.; Lurz, R.; Trautner, T.A. Bacteriophage SPP1. In The Bacteriophages; Calendar, R., Ed.; Oxford University Press: New York, NY, USA, 2006; Volume 54, p. 746. ISBN 9780195148503. [Google Scholar]
- Tavares, P.; Zinn-Justin, S.; Orlova, E.V. Genome gating in tailed bacteriophage capsids. In Viral Molecular Machines; Rossmann, M.G., Rao, V.B., Eds.; Advances in Experimental Medicine and Biology; Springer: Boston, MA, USA, 2012; Volume 726, pp. 585–600. ISBN 978-1-4614-0979-3. [Google Scholar]
- Oliveira, L.; Tavares, P.; Alonso, J.C. Headful DNA packaging: bacteriophage SPP1 as a model system. Virus Res. 2013, 173, 247–259. [Google Scholar] [CrossRef]
- Tavares, P. The bacteriophage head-to-tail interface. In Subcellular Biochemistry; Harris, J.R., Bhella, D., Eds.; Springer: Singapore, 2018; Volume 88, pp. 305–328. ISBN 978-981-10-8456-0. [Google Scholar]
- Hendrix, R.W.; Duda, R.L. Bacteriophage lambda PaPa: Not the mother of all lambda phages. Science 1992, 258, 1145–1148. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.W.; Landthaler, M.; Shub, D.A.; Stoddard, B.L. DNA binding and bleavage by the HNH Homing Endonuclease I-HmuI. J. Mol. Biol. 2004, 342, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Edgell, D.R.; Gibb, E.A.; Belfort, M. Mobile DNA elements in T4 and related phages. Virol. J. 2010, 7, 290. [Google Scholar] [CrossRef]
- Landthaler, M.; Lau, N.C.; Shub, D.A. Group I intron homing in Bacillus phages SPO1 and SP82: A gene conversion event initiated by a nicking homing endonuclease. J. Bacteriol. 2004, 186, 4307–4314. [Google Scholar] [CrossRef] [PubMed]
- Quiles-Puchalt, N.; Carpena, N.; Alonso, J.C.; Novick, R.P.; Marina, A.; Penadés, J.R. Staphylococcal pathogenicity island DNA packaging system involving cos-site packaging and phage-encoded HNH endonucleases. Proc. Natl. Acad. Sci. USA 2014, 111, 6016–6021. [Google Scholar] [CrossRef] [PubMed]
- Kala, S.; Cumby, N.; Sadowski, P.D.; Hyder, B.Z.; Kanelis, V.; Davidson, A.R.; Maxwell, K.L. HNH proteins are a widespread component of phage DNA packaging machines. Proc. Natl. Acad. Sci. USA 2014, 111, 6022–6027. [Google Scholar] [CrossRef]
- Neamah, M.M.; Mir-Sanchis, I.; López-Sanz, M.; Acosta, S.; Baquedano, I.; Haag, A.F.; Marina, A.; Ayora, S.; Penadés, J.R. Sak and Sak4 recombinases are required for bacteriophage replication in Staphylococcus aureus. Nucleic Acids Res. 2017, 45, 6507–6519. [Google Scholar] [CrossRef]
- Ogunleye, A.; Irorere, V.U.; Williams, C.; Hill, D.; Bhat, A.; Radecka, I. Poly-γ-glutamic acid: production, properties and applications. Microbiology 2015, 161, 1–17. [Google Scholar] [CrossRef]
- Mamberti, S.; Prati, P.; Cremaschi, P.; Seppi, C.; Morelli, C.F.; Galizzi, A.; Fabbi, M.; Calvio, C. γ-PGA hydrolases of phage origin in Bacillus subtilis and other microbial genomes. PLoS ONE 2015, 10, 1–17. [Google Scholar] [CrossRef]
- Kimura, K.; Itoh, Y. Characterization of poly-γ-glutamate hydrolase encoded by a bacteriophage genome: possible role in phage infection of Bacillus subtilis encapsulated with poly-γ-glutamate. Appl. Environ. Microbiol. 2003, 69, 2491–2497. [Google Scholar] [CrossRef]
- Ghosh, K.; Senevirathne, A.; Kang, H.; Hyun, W.; Kim, J.; Kim, K.-P.; Ghosh, K.; Senevirathne, A.; Kang, H.S.; Hyun, W.; et al. Complete nucleotide sequence analysis of a novel Bacillus subtilis-infecting bacteriophage BSP10 and its effect on poly-gamma-glutamic acid degradation. Viruses 2018, 10, 240. [Google Scholar] [CrossRef] [PubMed]
- Molshanski-Mor, S.; Yosef, I.; Kiro, R.; Edgar, R.; Manor, M.; Gershovits, M.; Laserson, M.; Pupko, T.; Qimron, U. Revealing bacterial targets of growth inhibitors encoded by bacteriophage T7. Proc. Natl. Acad. Sci. USA 2014, 111, 18715–18720. [Google Scholar] [CrossRef] [PubMed]
- Bondy-Denomy, J.; Qian, J.; Westra, E.R.; Buckling, A.; Guttman, D.S.; Davidson, A.R.; Maxwell, K.L. Prophages mediate defense against phage infection through diverse mechanisms. ISME J. 2016, 10, 2854–2866. [Google Scholar] [CrossRef] [PubMed]
- Dedrick, R.M.; Jacobs-Sera, D.; Bustamante, C.A.G.; Garlena, R.A.; Mavrich, T.N.; Pope, W.H.; Reyes, J.C.C.; Russell, D.A.; Adair, T.; Alvey, R.; et al. Prophage-mediated defence against viral attack and viral counter-defence. Nat. Microbiol. 2017, 2, 16251. [Google Scholar] [CrossRef] [PubMed]
- Seco, E.M.; Ayora, S. Bacillus subtilis DNA polymerases, PolC and DnaE, are required for both leading and lagging strand synthesis in SPP1 origin-dependent DNA replication. Nucleic Acids Res. 2017, 45, 8302–8313. [Google Scholar] [CrossRef]
- Alonso, J.C.; Ayora, S.; Canosa, I.; Weise, F.; Rojo, F. Site-specific recombination in Gram-positive theta-replicating plasmids. FEMS Microbiol. Lett. 1996, 142, 1–10. [Google Scholar] [CrossRef]
- Stahl, F.W.; Murray, N.E. The evolution of gene clusters and genetic circularity in microorganisms. Genetics 1966, 53, 569–576. [Google Scholar]
- Young, R. Phage lysis: Do we have the hole story yet? Curr. Opin. Microbiol. 2013, 16, 790–797. [Google Scholar] [CrossRef]
- Krogh, S.; Jørgensen, S.T.; Devine, K.M. Lysis genes of the Bacillus subtilis defective prophage PBSX. J. Bacteriol. 1998, 180, 2110–2117. [Google Scholar]
- Bläsi, U.; Young, R. Two beginnings for a single purpose: The dual-start holins in the regulation of phage lysis. Mol. Microbiol. 1996, 21, 675–682. [Google Scholar] [CrossRef]
- Abrescia, N.G.A.A.; Bamford, D.H.; Grimes, J.M.; Stuart, D.I. Structure unifies the viral universe. Annu. Rev. Biochem. 2012, 81, 795–822. [Google Scholar] [CrossRef] [PubMed]
- Veesler, D.; Cambillau, C. A common evolutionary origin for tailed-bacteriophage functional modules and bacterial machineries. Microbiol. Mol. Biol. Rev. 2011, 75, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Jamet, A.; Touchon, M.; Ribeiro-Gonçalves, B.; Carriço, J.A.; Charbit, A.; Nassif, X.; Ramirez, M.; Rocha, E.P.C. A widespread family of polymorphic toxins encoded by temperate phages. BMC Biol. 2017, 15, 75. [Google Scholar] [CrossRef] [PubMed]
- Jamet, A.; Charbit, A.; Nassif, X. Antibacterial toxins: Gram-positive bacteria strike back! Trends Microbiol. 2018, 26, 89–91. [Google Scholar] [CrossRef] [PubMed]
- Deichelbohrer, I.; Messer, W.; Trautner, T.A. Genome of Bacillus subtilis bacteriophage SPP1: structure and nucleotide sequence of pac, the origin of DNA packaging. J. Virol. 1982, 42, 83–90. [Google Scholar] [PubMed]
- Bravo, A.; Alonso, J.C.; Trautner, T.A. Functional analysis of the Bacillus subtilis bacteriophage SPP1 pac site. Nucleic Acids Res. 1990, 18, 2881. [Google Scholar] [CrossRef] [PubMed]
- Dröge, A.; Tavares, P. In vitro packaging of DNA of the Bacillus subtilis bacteriophage SPP1. J. Mol. Biol. 2000, 296, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.; Alonso, J.C.; Tavares, P. A defined in vitro system for DNA packaging by the bacteriophage SPP1: Insights into the headful packaging mechanism. J. Mol. Biol. 2005, 353, 529–539. [Google Scholar] [CrossRef]
- Tavares, P.; Dröge, A.; Lurz, R.; Graeber, I.; Orlova, E.; Dube, P.; Van Heel, M. The SPP1 connection. FEMS Microbiol. Rev. 1995, 17, 47–56. [Google Scholar] [CrossRef]
- Bönemann, G.; Pietrosiuk, A.; Mogk, A. Tubules and donuts: a type VI secretion story: MicroReview. Mol. Microbiol. 2010, 76, 815–821. [Google Scholar] [CrossRef]
- Scholl, D. Phage Tail–Like Bacteriocins. Annu. Rev. Virol. 2017, 4, 453–467. [Google Scholar] [CrossRef] [PubMed]
- São-José, C.; Baptista, C.; Santos, M.A. Bacillus subtilis operon encoding a membrane receptor for bacteriophage SPP1. J. Bacteriol. 2004, 186, 8337–8346. [Google Scholar] [CrossRef] [PubMed]
- King, J. Assembly of the tail of bacteriophage T4. J. Mol. Biol. 1968, 32, 231–262. [Google Scholar] [CrossRef]
- Katsura, I.; Kühl, P.W. Morphogenesis of the tail of bacteriophage lambda: III. Morphogenetic pathway. J. Mol. Biol. 1975, 91, 257–273. [Google Scholar] [CrossRef]
- Ghosh, N.; McKillop, T.J.; Jowitt, T.A.; Howard, M.; Davies, H.; Holmes, D.F.; Roberts, I.S.; Bella, J. Collagen-like proteins in pathogenic E. coli strains. PLoS ONE 2012, 7, e37872. [Google Scholar] [CrossRef] [PubMed]
- Brüssow, H.; Desiere, F. Comparative phage genomics and the evolution of Siphoviridae: Insights from dairy phages. Mol. Microbiol. 2001, 39, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Casjens, S.R. Comparative genomics and evolution of the tailed-bacteriophages. Curr. Opin. Microbiol. 2005, 8, 451–458. [Google Scholar] [CrossRef]
- Stockdale, S.R.; Collins, B.; Spinelli, S.; Douillard, F.P.; Mahony, J.; Cambillau, C.; van Sinderen, D. Structure and assembly of TP901-1 virion unveiled by mutagenesis. PLoS ONE 2015, 10, e0131676. [Google Scholar] [CrossRef]
- Murphy, J.; Bottacini, F.; Mahony, J.; Kelleher, P.; Neve, H.; Zomer, A.; Nauta, A.; van Sinderen, D. Comparative genomics and functional analysis of the 936 group of lactococcal Siphoviridae phages. Sci. Rep. 2016, 6, 21345. [Google Scholar] [CrossRef]
- Lopes, A.; Tavares, P.; Petit, M.-A.; Guérois, R.; Zinn-Justin, S. Automated classification of tailed bacteriophages according to their neck organization. BMC Genom. 2014, 15, 1027. [Google Scholar] [CrossRef]
- Zivanovic, Y.; Confalonieri, F.; Ponchon, L.; Lurz, R.; Chami, M.; Flayhan, A.; Renouard, M.; Huet, A.; Decottignies, P.; Davidson, A.R.; et al. Insights into bacteriophage T5 structure from analysis of its morphogenesis genes and protein components. J. Virol. 2014, 88, 1162–1174. [Google Scholar] [CrossRef] [PubMed]
- Fraser, J.S.; Yu, Z.; Maxwell, K.L.; Davidson, A.R. Ig-Like Domains on bacteriophages: a tale of promiscuity and deceit. J. Mol. Biol. 2006, 359, 496–507. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, S.; Veesler, D.; Bebeacua, C.; Cambillau, C. Structures and host-adhesion mechanisms of lactococcal siphophages. Front. Microbiol. 2014, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Dowah, A.S.A.; Clokie, M.R.J. Review of the nature, diversity and structure of bacteriophage receptor binding proteins that target Gram-positive bacteria. Biophys. Rev. 2018, 10, 535–542. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Position (nt) | ORF | ORF Sequence Change | ORF Product Amino Acid Change |
|---|---|---|---|
| 230 | 1 | insertion of a G; frameshift | gp1 Cter sequence changed and shortened |
| 766 | 2 | T→C | none (silent mutation) |
| 1997 | 4 | insertion of an A; frameshift | gp4 Cter changed and lengthened |
| 5001 | intergenic region ORFs 7 and 8 | insertion of a T | none |
| 23,794 | 30 | deletion of a C; frameshift | gp30 Cter changed and lengthened |
| 34,450 | 37.1 | T→G | gp37.1 C18→G18 |
| 37,304 and 37,306 | 40 | GGC→CGG | gp40 R194R195→P194G195 |
| 38,371 to 38,374 | 41 | GTGT→TGTG | gp41 K101C102→N101V102 |
| 40,240 | 44 and former 45 | insertion of a G; frameshift; ORFs fusion | gp44 Cter lengthened by former gp45 sequence |
| 41,787 | 48 | insertion of an A; frameshift | gp48 changed and shortened |
| 42,750 | 51 | insertion of an A; frameshift | gp51 changed and lengthened |
| 42,778 to 42,781 | 51 | CGCG→GCGC | gp51 R56E57→A56Q57 |
| 42,819 | 51 | insertion of a G; frameshift | gp51 Cter lengthened |
| Terminator Sequence (Loop,Stem) | Start (nt) | End (nt) | ΔG° (kcal.mol−1) |
| AAAAGCGAGTTAACCGACGTAAAAATGCGTCGGTTTTTTTCGTGCTTC | 4718 | 4765 | −12.20 |
| GTGTCAGTGCGCGGGTTCAATTCCCGGAGTCCGTTTTTACCGCCC | 6780 | 6824 | −5.50 |
| TAAAAAGAGAAGAGGGGCTAACGCCTCTCTTTTTTTGAAAG | 8775 | 8815 | −14.00 |
| ATTGCCAGCAGAGAGCACGGGTTAATTCCCGCGCTCTTTTTTTGTATTCA | 11,255 | 11,304 | −9.20 |
| TGAATTAGACAGGGCCGCGCAAGTGGCTCTTTTTAATAGGT | 12,009 | 12,049 | −15.20 |
| TGAAAAGACGCGGCGCCGCTAACGGCGTCCGTTTTGAACATGA | 16,711 | 16,753 | −12.10 |
| GATTGACGAAGTAAAGGGCCATGTGCCCTTTATTTTTTTGCAAA | 21,897 | 21,940 | −10.70 |
| ACCTTCGCTTGCCGCCCGGCTGATGGGCGGTTTTTTATTTTT | 24,095 | 24,136 | −13.20 |
| TGTCCTAAAATCGGCCCGTTCCCAGTCGGGCCACTTTTTTTATTTTA | 27,934 | 27,980 | −13.60 |
| GACTTTGAAAGGAACCGTTCTCTAACGGTTCTTTTTTTATTTC | 30,205 | 30,247 | −9.90 |
| TATTTTGATTGAGTCGGGGAAACCCGGCTTTTTTATTTTGGG | 35,232 | 35,273 | −14.90 |
| ATCAAAGTTATGGTGGGAGTAATCCCGCCTTTTTCTATTTT | 40,466 | 40,506 | −14.40 |
| AACACAGAGAGGCACCCTATTTGGGTGCTTTTTTGTTGTA | 43,774 | 43,813 | −11.90 |
| ORF | Protein | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ORF | RBS (mRNA) | Spacing | Start | nt | Stop | nt | Essential | Length | MM | Predicted TMM | 3D Structure | Protein Function | References | |
| Gene | (aa) | (kDa) | Segments | (PDB or EMD) | ||||||||||
| 1 | AAGGAGGU | 10 | AUG | 43,884 | UGA | 311 | yes | 147 | 16.3 | no | 3ZQQ a (Xtal b) | small terminase subunit (TerS) | [57,63,64,65,66,67,68,69] | |
| 2 | n.d. c | AUG d | 308 d | UAG | 1576 | yes | 422 d | 48.8 d | 1 e | 2WBN f; 2WC9 f (Xtal b) | large terminase subunit (TerL) | [57,67,70,71,72,73,74] | ||
| 3 | AAAGGAGG | 11 | AUG | 1567 | UAA | 1782 | no | 71 | 8.5 | no | n.d. | unknown | ||
| 4 | GGgGGU | 10 | AUG | 1782 | UAA | 2072 | no | 96 | 11.4 | no | n.d. | unknown | ||
| 5 | AAGGAGG | 11 | AUG | 2065 | UGA | 2334 | no | 89 | 10.3 | no | n.d. | unknown | ||
| 6 | AGGAGGU | 11 | AUG | 2336 | UGA | 3847 | yes | 503 | 57.3 | no | 2JES (Xtal b); | st.; portal protein | [20,60,67,73,74,75,76,77,78,79,80,81,82] | |
| 5A20, 5A21 (cryoEM g) | ||||||||||||||
| 7 | AGGAGG | 12 | AUG | 3804 | UAA | 4730 | no | 308 | 35.1 | no | n.d. | st.; initiation of infection; binds to portal | [83,84] | |
| 8 | AAAGGAG | 12 | AUG | 5067 | UGA | 6215 | n.d. | 382 | 43.7 | no | n.d. | unknown | ||
| 9 | AAcGGAGG | 9 | AUG | 6217 | UAA | 6555 | n.d. | 112 | 12.6 | no | n.d. | unknown | ||
| 10 | GGuGGUG h | 12 h | AUG | 6583 | UAG | 6750 | n.d. | 55 | 6.2 | no | n.d. | unknown | ||
| 11 | AGGAG | 9 | AUG | 6917 | UAA | 7561 | yes | 214 | 23.4 | no | n.d. | procapsid scaffolding protein | [85,86,87] | |
| 12 | AAGGgGG | 11 | AUG | 7576 | UAA | 7770 | no | 64 | 6.6 | no | n.d. | st.; capsid accessory protein with collagen-like fold | [15,88] | |
| 13 | AAAGGAG | 9 | AUG | 7803 | UAA | 8777 | yes | 324 | 35.3 | no | 4AN5 (cryoEM g) | st.; major capsid protein (MCP) | [15,85,86] | |
| 14 | AAAGGAG | 10 | AUG | 8828 | UGA | 9004 | no | 58 | 6.7 | no | n.d. | unknown | ||
| 15 | AAuGAGG | 10 | AUG | 9015 | UAA | 9323 | yes | 102 | 11.6 | no | 2KBZ (NMR i); | st.; connector adaptor protein | [60,78,81,89] | |
| 5A20, 5A21 (cryoEM g) | ||||||||||||||
| 16 | n.d. c | AUG j | 9325 j | UAG | 9654 | yes | 109 j | 12.5 j | no | 2KCA (NMR i) | st.; connector stopper protein | [60,78,81,89] | ||
| 5A20, 5A21 (cryoEM g) | ||||||||||||||
| 16.1 | AAaGAGG | 11 | AUG | 9644 | UGA | 10,069 | n.d. | 141 | 15.9 | no | n.d. | putative tail protein | ||
| 17 | AGGAGGU | 10 | AUG | 10,066 | UGA | 10,470 | yes | 134 | 15 | no | 2LFP (NMR i) | st.; tail-to-head joining protein (THJP) | [32,90] | |
| 17.1 | AGGAGG | 10 | AUG | 10,484 k | UAA | 11,017 | yes | 177 | 19.2 | no | n.d. | st.; tail tube protein (TTP) | [16,91,92] | |
| 17.1* | AGGAGG | 10 | AUG | 10,484 k | UAA | 11,279 | no | 264 | 28.2 | no | n.d. | st.; tail tube protein; Cter FN3 motif | [91] | |
| 17.5 | GAGG | 12 | AUG | 11,363 l | UAA | 11,884 | n.d. m | 173 | 20.2 | no | n.d. | tail chaperone protein | [91] | |
| 17.5* | GAGG | 12 | AUG | 11,363 l | UAG | 12,255 | n.d. m | 297 | 34 | no | n.d. | tail chaperone protein | [91] | |
| 18 | AGGAGG | 9 | AUG | 12,267 | UGA | 15,365 | n.d. m | 1032 | 110.9 | 4 n | n.d. | st.; tape measure protein (TMP) | [16] | |
| 19.1 | GAGG | 10 | AUG | 15,362 | UAA | 16,123 | n.d. m | 253 | 28.6 | no | 2X8K (Xtal b) | st.; distal tail protein (Dit) | [93,94] | |
| 21 | AAGaAGGUGA | 10 | UUG | 16,137 | UAA | 19,463 | n.d. m | 1108 | 123.6 | no | n.d. | st.; tail tip protein; Tal; anti-receptor protein | [93,95] | |
| 22 | AAGGAGG | 9 | AUG | 19,476 | UAA | 19,916 | n.d. | 146 | 16.7 | no | 2XC8 (Xtal b) | putative tail protein | [96] | |
| 23 | AGGAGGU | 10 | AUG | 19,932 | UGA | 20,096 | n.d. | 54 | 6.1 | no | n.d. | n.d. | ||
| 23.1 | GGAG | 9 | AUG | 20,089 | UAA | 20,244 | n.d. | 51 | 5.8 | no | 2XF7 (Xtal b) | n.d. | [97] | |
| 24 | GgGGUG h | 10 h | AUG | 20,237 | UAG | 20,467 | n.d. | 76 | 8.4 | no | n.d. | n.d. | ||
| 24.1 | AAAGGgGG | 11 | AUG | 20,547 | UAA | 20,825 | n.d. | 92 | 10.6 | 1 | n.d. | component of holin; cell lysis | [33,98] | |
| 24.1* | AGGAGGU | 10 | AUG | 20,574 | UAA | 20,825 | n.d. | 83 | 9.5 | 1 | n.d. | component of holin; cell lysis | [33] | |
| 25 | AAGGAG | 12 | AUG | 20,845 | UAA | 21,660 | n.d. | 271 | 29.9 | no | n.d. | endolysin; cell lysis | [33,98] | |
| 26 | AAAGGAG | 8 | AUG | 21,662 | UAA | 21,910 | n.d. | 82 | 9.4 | 2 | n.d. | component of holin; cell lysis | [33,98] | |
| 26* | AAAGGAG o | 14 o | AUG | 21,668 | UAA | 21,910 | n.d. | 80 | 9.1 | 2 | n.d. | component of holin; cell lysis | [33] | |
| 26.1 | AAGGgGG o | 10 o | AUG | 22,009 | UAG | 22,152 | n.d. | 47 | 5.8 | no | n.d. | unknown | ||
| 27 | AAGGAGG | 12 | UUG | 22,277 | UAA | 22,831 | n.d. | 184 | 20.8 | no | n.d. | unknown | ||
| 28 | GGAGG | 9 | AUG | 22,834 | UGA | 23,121 | n.d. | 95 | 10.8 | no | n.d. | unknown | ||
| 29 | AGGAGG | 13 | AUG | 23,069 | UGA | 23,371 | n.d. | 100 | 12 | no | n.d. | putative DNA binding protein | ||
| 29.1 | AGGgGG | 9 | GUG | 23,358 | UGA | 23,675 | n.d. | 105 | 12.3 | no | n.d. | unknown | ||
| 30 | AGGgGG | 10 | AUG | 23,675 | UAA | 23,854 | n.d. | 59 | 7.2 | 1 | n.d. | unknown | ||
| 30.1 | AGGgGG | 9 | AUG | 23,859 | UGA | 24,029 | n.d. | 56 | 6.4 | no | n.d. | unknown | ||
| 31 | AAcGGAGGU | 12 | AUG | 24,209 | UAA | 24,493 | n.d. | 94 | 11 | no | n.d. | unknown | ||
| 31.1 | GAGG | 12 | AUG | 24,589 | UAA | 24,951 | n.d. | 120 | 12.9 | 3 | n.d. | unknown | ||
| 31.2 | GgGGUG h | 10 h | AUG | 24,964 | UGA | 25,281 | no p | 105 | 11.5 | 2 | n.d. | unknown | ||
| 32 | GGAGGUG | 8 | AUG | 25,278 | UAA | 27,788 | no p | 836 | 96.3 | no | n.d. | putative ATP-binding protein | ||
| 32.5 | GGAGGUG | 11 | UUG | 28,039 | UAA | 28,209 | n.d. | 56 | 6.7 | no | n.d. | unknown | ||
| 33 | AAAaGgGGU | 11 | AUG | 28,226 | UAA | 29,995 | no | 589 | 64.9 | no | n.d. | putative bacteria surface binding protein | ||
| 33.1 | AAcGGAGG | 9 | AUG | 30,011 | UAA | 30,229 | n.d. | 72 | 8.4 | no | n.d. | unknown | ||
| 34 | AGGgGG | 9 | AUG | 30,364 | UAG | 30,522 | n.d. | 52 | 6.3 | no | n.d. | putative transcriptional repressor | ||
| 34.1 | AGGAGG | 9 | AUG | 30,534 | UGA | 31,469 | no | 311 | 35.9 | no | n.d. | 5′-3′ exonuclease | [13,99] | |
| 34.2 | GGAGG | 12 | AUG | 31,466 | UAA | 31,639 | n.d. | 57 | 6.7 | no | n.d. | unknown | ||
| 34.3 | AAGGAGG | 11 | AUG | 31,641 | UAA | 31,895 | n.d. | 84 | 9.8 | no | n.d. | unknown | ||
| 34.4 | GGAGG | 11 | AUG | 31,897 | UAA | 32,187 | n.d. | 96 | 11.1 | no | n.d. | unknown | ||
| 35 | GGAGGU | 11 | AUG | 32,177 | UAG | 33,040 | yes | 287 | 32 | no | n.d. | recT-like recombinase | [13,100] | |
| 36 | AGGAGGgGA | 10 | AUG | 33,033 | UAA | 33,512 | no | 159 | 17.1 | no | n.d. | SSB | [101] | |
| 36.1 | AAaGgGGUGA h | 10 h | AUG | 33,537 | UGA | 34,028 | n.d. | 163 | 18.9 | no | n.d. | putative HNH endonuclease | ||
| 37 | GGAGG | 10 | AUG | 34,032 | UGA | 34,406 | n.d. | 124 | 14.3 | no | n.d. | unknown | ||
| 37.1 | GGAGG | 11 | AUG | 34,399 | UGA | 34,992 | n.d. | 197 | 22.3 | no | n.d. | putative poly-gamma-glutamate hydrolase | ||
| 37.2 | GaAGG | 14 | AUG | 34,989 | UGA | 35,243 | n.d. | 84 | 9.7 | no | n.d. | unknown | ||
| 37.3 | AGGAGG | 11 | AUG | 35,400 | UAG | 35,573 | n.d. | 57 | 6.7 | no | n.d. | putative DNA binding protein | ||
| 38 | AAGGAGG | 13 | AUG | 35,580 | UGA | 36,350 | yes | 256 | 30 | no | n.d. | SPP1 origin binding protein | ||
| and replication re-start (PriA-like) | [56,102] | |||||||||||||
| 39 | AGGAGG | 9 | AUG | 36,347 | UGA | 36,727 | yes | 126 | 14.6 | no | 1NO1 (Xtal b) | gp40 helicase loader | [56,103,104] | |
| 40 | GAGG | 11 | AUG | 36,724 | UAA | 38,052 | yes | 442 | 49.7 | no | 3BGW (Xtal b) | replicative DNA helicase; | ||
| binds host DnaG and DnaX | [56,103,105,106,107,108,109] | |||||||||||||
| 41 | AAAGGgGG | 10 | AUG | 38,069 | UGA | 38,569 | n.d. | 166 | 19.1 | no | n.d. | unknown | ||
| 42 | AAAGGAG | 11 | AUG | 38,566 | UAA | 38,961 | no | 131 | 16 | no | n.d. | unknown | ||
| 42.1 | GGAGG | 9 | AUG | 38,964 | UGA | 39,134 | no | 56 | 6.5 | no | n.d. | unknown | ||
| 42.2 | GgGG c, q | 12 c, q | AUG | 39,131 q | UAA | 39,427 | no | 98 q | 10.7 q | no | n.d. | unknown | ||
| 43 | GGAGG | 10 | GUG | 39,431 | UGA | 39,784 | no | 117 | 14.2 | no | n.d. | unknown | ||
| 44 | AAGGAG | 11 | AUG | 39,777 | UAA | 40,487 | no | 236 | 27.5 | no | n.d. | Holliday junction resolvase | [13,31] | |
| 46 | GAGG | 12 | AUG | 40,596 | UAA | 40,898 | no | 100 | 11.5 | no | n.d. | unknown | ||
| 46.1 | AGGAGG | 9 | AUG | 40,898 | UAA | 41,209 | no | 103 | 11.7 | 3 | n.d. | unknown | ||
| 47 | AGGgGG | 9 | AUG | 41,304 | UGA | 41,663 | no | 119 | 13.7 | no | n.d. | unknown | ||
| 48 | GGAG | 13 | AUG | 41,645 | UGA | 41,995 | no | 116 | 13.2 | no | n.d. | unknown | ||
| 49 | GGAGG | 9 | GUG | 42,075 | UGA | 42,248 | no | 57 | 6.5 | no | n.d. | unknown | ||
| 50 | AAGGAGG | 9 | GUG | 42,245 | UAA | 42,418 | no | 57 | 6.8 | no | n.d. | unknown | ||
| 50.1 | AAAGGAGG | 9 | GUG | 42,434 | UGA | 42,616 | no | 60 | 6.7 | 2 | n.d. | unknown | ||
| 51 | AAGGAGG | 9 | AUG | 42,613 | UAA | 43,014 | no | 133 | 14.7 | 1 | n.d. | unknown | ||
| 51.1 | AAGGAG | 9 | AUG | 43,027 | UGA | 43,182 | no | 51 | 6.1 | 1 | n.d. | unknown | ||
| 52 | AAAGGAG | 10 | AUG | 43,179 | UGA | 43,421 | no | 80 | 9.5 | no | n.d. | unknown | ||
| 53 | AAAGGAG | 10 | AUG | 43,405 | UGA | 43,611 | no | 68 | 7.4 | 1 | n.d. | unknown | ||
| Amino Acid | Codon | Fraction | No | Amino Acid | Codon | Fraction | No | Amino Acid | Codon | Fraction | No | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ala | GCG | SPP1 | 0.26 | 268 | Gly | GGA | SPP1 | 0.31 | 318 | Pro | CCU | SPP1 | 0.22 | 107 |
| B.s | 0.26 | 24,574 | B.s | 0.31 | 26,381 | B.s | 0.29 | 12,824 | ||||||
| Ala | GCA | SPP1 | 0.27 | 272 | Gly | GGU | SPP1 | 0.24 | 245 | Pro | CCC | SPP1 | 0.08 | 42 |
| B.s | 0.28 | 26,416 | B.s | 0.18 | 15,457 | B.s | 0.09 | 4001 | ||||||
| Ala | GCU | SPP1 | 0.27 | 279 | Gly | GGC | SPP1 | 0.25 | 262 | Ser | AGU | SPP1 | 0.16 | 123 |
| B.s | 0.25 | 23,062 | B.s | 0.34 | 28,493 | B.s | 0.11 | 8096 | ||||||
| Ala | GCC | SPP1 | 0.20 | 201 | His | CAU | SPP1 | 0.53 | 131 | Ser | AGC | SPP1 | 0.22 | 170 |
| B.s | 0.21 | 19,342 | B.s | 0.67 | 18,610 | B.s | 0.23 | 17,226 | ||||||
| Arg | AGG | SPP1 | 0.18 | 130 | His | CAC | SPP1 | 0.47 | 119 | Ser | UCG | SPP1 | 0.10 | 79 |
| B.s | 0.10 | 4788 | B.s | 0.33 | 9019 | B.s | 0.10 | 7717 | ||||||
| Arg | AGA | SPP1 | 0.28 | 198 | Ile | AUA | SPP1 | 0.27 | 261 | Ser | UCA | SPP1 | 0.25 | 193 |
| B.s | 0.26 | 13,077 | B.s | 0.13 | 11,517 | B.s | 0.24 | 18,053 | ||||||
| Arg | CGG | SPP1 | 0.12 | 83 | Ile | AUU | SPP1 | 0.35 | 335 | Ser | UCU | SPP1 | 0.17 | 132 |
| B. s | 0.15 | 7329 | B.s | 0.50 | 45,181 | B.s | 0.20 | 15,615 | ||||||
| Arg | CGA | SPP1 | 0.10 | 74 | Ile | AUC | SPP1 | 0.38 | 365 | Ser | UCC | SPP1 | 0.11 | 83 |
| B. s | 0.11 | 5115 | B.s | 0.37 | 32,872 | B.s | 0.13 | 9757 | ||||||
| Arg | CGU | SPP1 | 0.18 | 125 | Leu (s) | UUG | SPP1 | 0.24 | 246 | Thr | ACG | SPP1 | 0.26 | 211 |
| B. s | 0.18 | 8755 | B.s | 0.16 | 18,745 | B.s | 0.27 | 17,693 | ||||||
| Arg | CGC | SPP1 | 0.14 | 99 | Leu | UUA | SPP1 | 0.24 | 251 | Thr | ACA | SPP1 | 0.47 | 379 |
| B. s | 0.20 | 9444 | B.s | 0.20 | 23,338 | B.s | 0.41 | 27,117 | ||||||
| Asn | AAU | SPP1 | 0.45 | 344 | Leu | CUG | SPP1 | 0.10 | 107 | Thr | ACU | SPP1 | 0.15 | 120 |
| B.s | 0.57 | 27,137 | B.s | 0.24 | 28,295 | B.s | 0.16 | 10,620 | ||||||
| Asn | AAC | SPP1 | 0.55 | 425 | Leu | CUA | SPP1 | 0.13 | 139 | Thr | ACC | SPP1 | 0.12 | 94 |
| B.s | 0.43 | 20,861 | B.s | 0.05 | 6030 | B.s | 0.16 | 10,497 | ||||||
| Asp | GAU | SPP1 | 0.52 | 442 | Leu | CUU | SPP1 | 0.20 | 203 | Trp | UGG | SPP1 | 1.00 | 190 |
| B.s | 0.64 | 40,291 | B.s | 0.24 | 28,226 | B.s | 1.00 | 12,571 | ||||||
| Asp | GAC | SPP1 | 0.48 | 415 | Leu | CUC | SPP1 | 0.08 | 87 | Tyr | UAU | SPP1 | 0.50 | 276 |
| B.s | 0.36 | 22,699 | B.s | 0.11 | 13,232 | B.s | 0.65 | 27,650 | ||||||
| Cys | UGU | SPP1 | 0.54 | 50 | Lys | AAG | SPP1 | 0.38 | 460 | Tyr | UAC | SPP1 | 0.50 | 278 |
| B.s | 0.45 | 4429 | B.s | 0.30 | 25,647 | B.s | 0.35 | 14,673 | ||||||
| Cys | UGC | SPP1 | 0.46 | 42 | Lys | AAA | SPP1 | 0.62 | 760 | Val (s) | GUG | SPP1 | 0.23 | 211 |
| B.s | 0.55 | 5322 | B.s | 0.70 | 60,072 | B.s | 0.26 | 21,585 | ||||||
| Gln | CAG | SPP1 | 0.38 | 192 | Met (s) | AUG | SPP1 | 1.00 | 412 | Val | GUA | SPP1 | 0.26 | 240 |
| B.s | 0.46 | 22,750 | B.s | 1.00 | 32,918 | B.s | 0.20 | 16,296 | ||||||
| Gln | CAA | SPP1 | 0.62 | 319 | Phe | UUU | SPP1 | 0.48 | 270 | Val | GUU | SPP1 | 0.32 | 292 |
| B.s | 0.54 | 23,889 | B.s | 0.68 | 37,445 | B.s | 0.28 | 23,440 | ||||||
| Glu | GAG | SPP1 | 0.39 | 452 | Phe | UUC | SPP1 | 0.52 | 294 | Val | GUC | SPP1 | 0.18 | 168 |
| B.s | 0.32 | 28,211 | B.s | 0.32 | 17,253 | B.s | 0.26 | 21,143 | ||||||
| Glu | GAA | SPP1 | 0.61 | 704 | Pro | CCG | SPP1 | 0.44 | 219 | End | UGA | SPP1 | 0.39 | 31 |
| B.s | 0.68 | 59,808 | B.s | 0.43 | 19,421 | B.s | 0.24 | 965 | ||||||
| Gly | GGG | SPP1 | 0.21 | 215 | Pro | CCA | SPP1 | 0.26 | 127 | End | UAG | SPP1 | 0.11 | 9 |
| B.s | 0.16 | 13,670 | B.s | 0.19 | 8541 | B.s | 0.14 | 591 | ||||||
| End | UAA | SPP1 | 0.50 | 40 | ||||||||||
| B.s | 0.62 | 2542 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Godinho, L.M.; El Sadek Fadel, M.; Monniot, C.; Jakutyte, L.; Auzat, I.; Labarde, A.; Djacem, K.; Oliveira, L.; Carballido-Lopez, R.; Ayora, S.; et al. The Revisited Genome of Bacillus subtilis Bacteriophage SPP1. Viruses 2018, 10, 705. https://doi.org/10.3390/v10120705
Godinho LM, El Sadek Fadel M, Monniot C, Jakutyte L, Auzat I, Labarde A, Djacem K, Oliveira L, Carballido-Lopez R, Ayora S, et al. The Revisited Genome of Bacillus subtilis Bacteriophage SPP1. Viruses. 2018; 10(12):705. https://doi.org/10.3390/v10120705
Chicago/Turabian StyleGodinho, Lia M., Mehdi El Sadek Fadel, Céline Monniot, Lina Jakutyte, Isabelle Auzat, Audrey Labarde, Karima Djacem, Leonor Oliveira, Rut Carballido-Lopez, Silvia Ayora, and et al. 2018. "The Revisited Genome of Bacillus subtilis Bacteriophage SPP1" Viruses 10, no. 12: 705. https://doi.org/10.3390/v10120705
APA StyleGodinho, L. M., El Sadek Fadel, M., Monniot, C., Jakutyte, L., Auzat, I., Labarde, A., Djacem, K., Oliveira, L., Carballido-Lopez, R., Ayora, S., & Tavares, P. (2018). The Revisited Genome of Bacillus subtilis Bacteriophage SPP1. Viruses, 10(12), 705. https://doi.org/10.3390/v10120705