Environmental Conditions and Species Identity Drive Metabolite Levels in Green Leaves and Leaf Litter of 14 Temperate Woody Species


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Species and Leaf Sampling
2.2. Quantitative Foliar Chemical Analyses
2.3. Statistical Analyses
3. Results
3.1. Species-Specific Differences between Leaf Metabolites
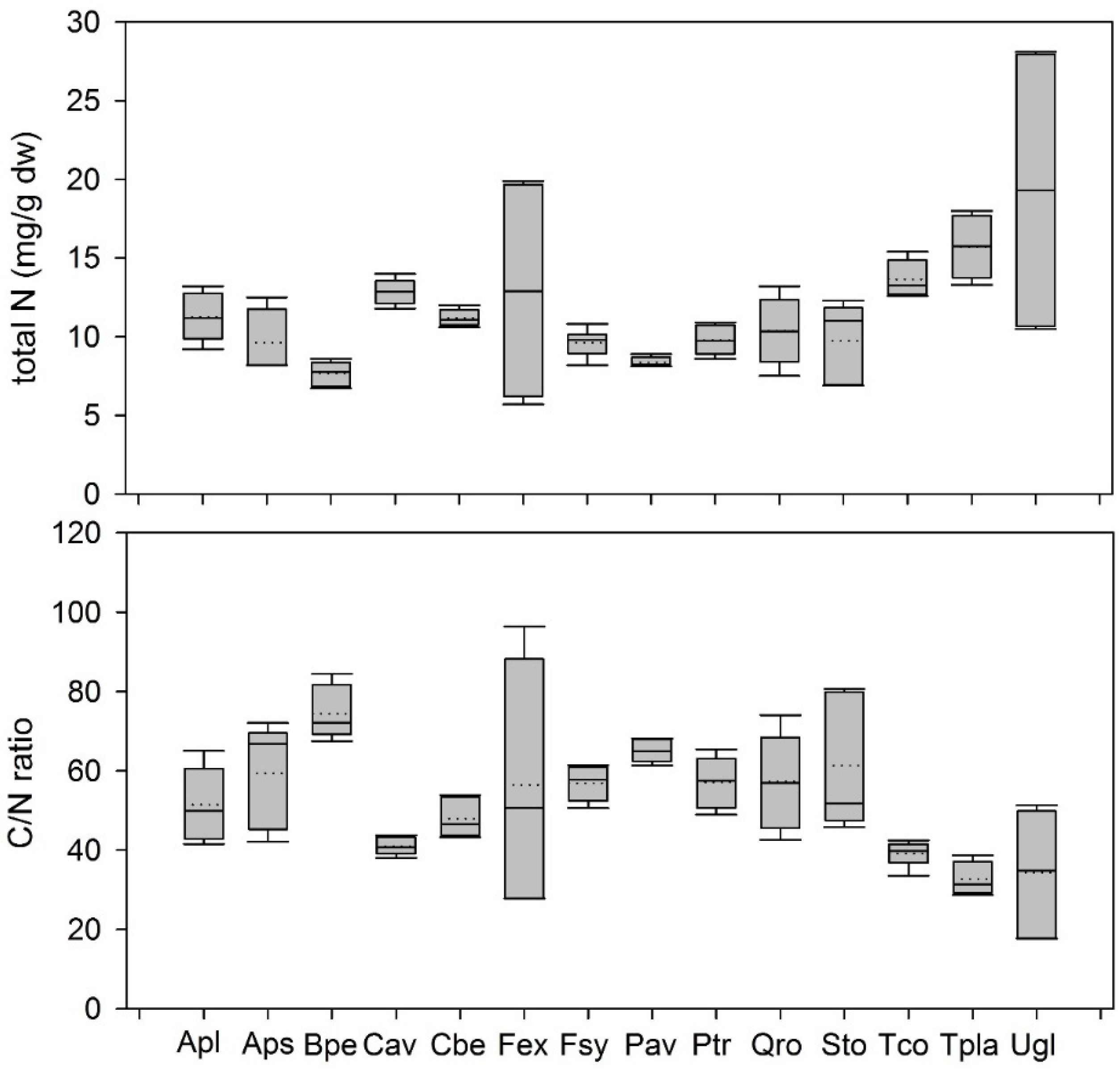
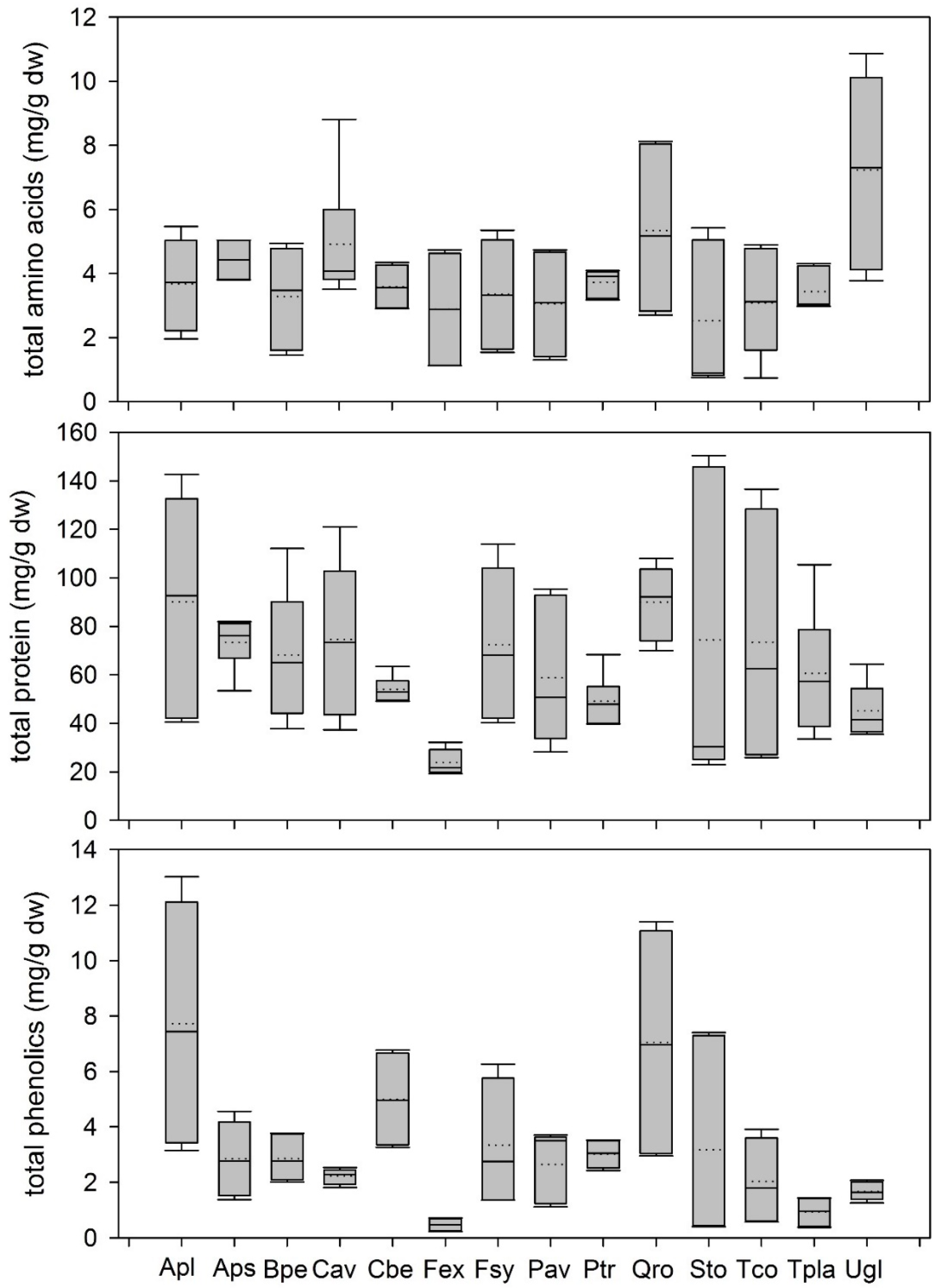
3.1.1. Variation of Leaf Metabolites in Leaf Litter
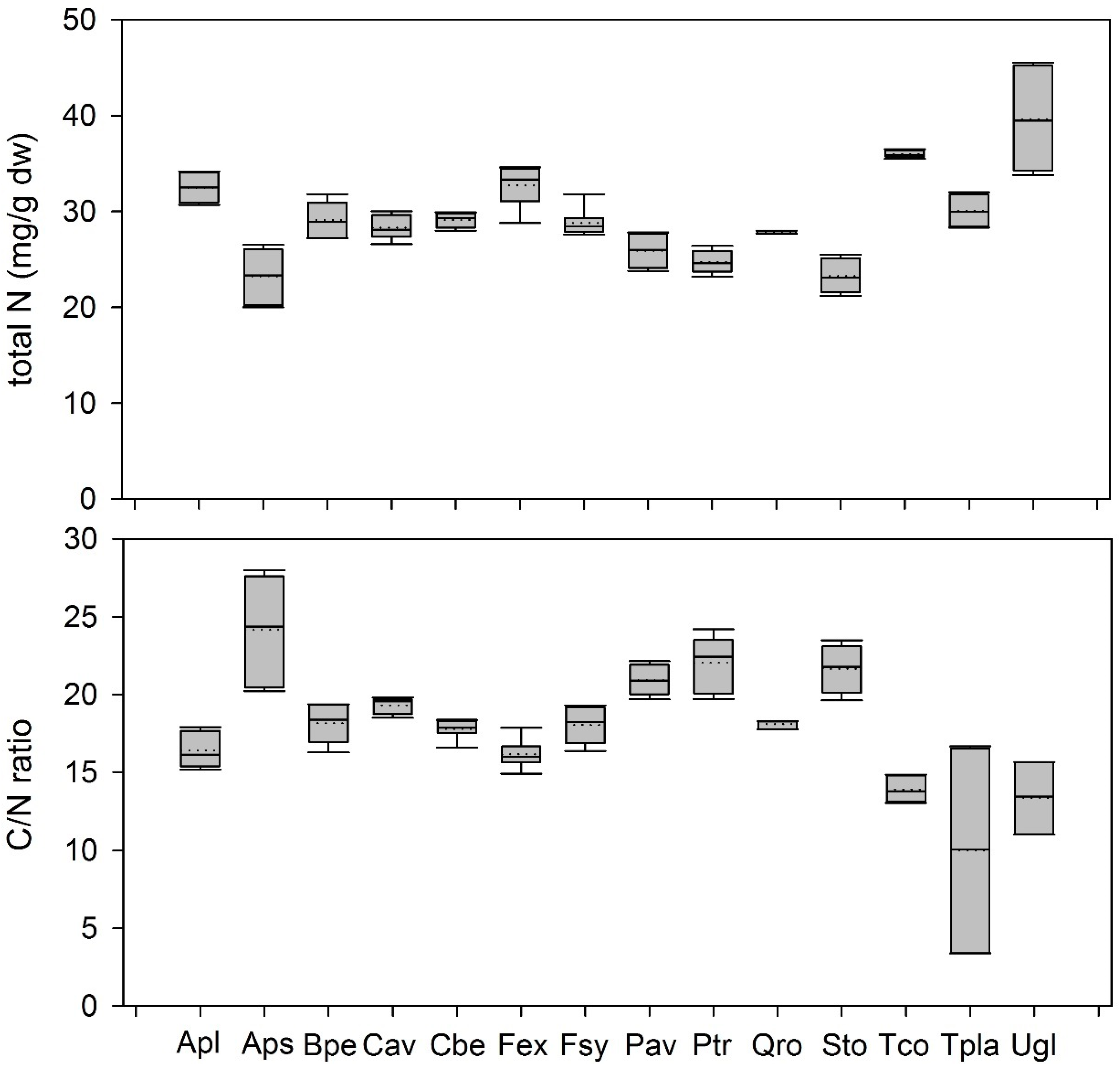
3.1.2. Variation of Leaf Metabolites in Green Leaves
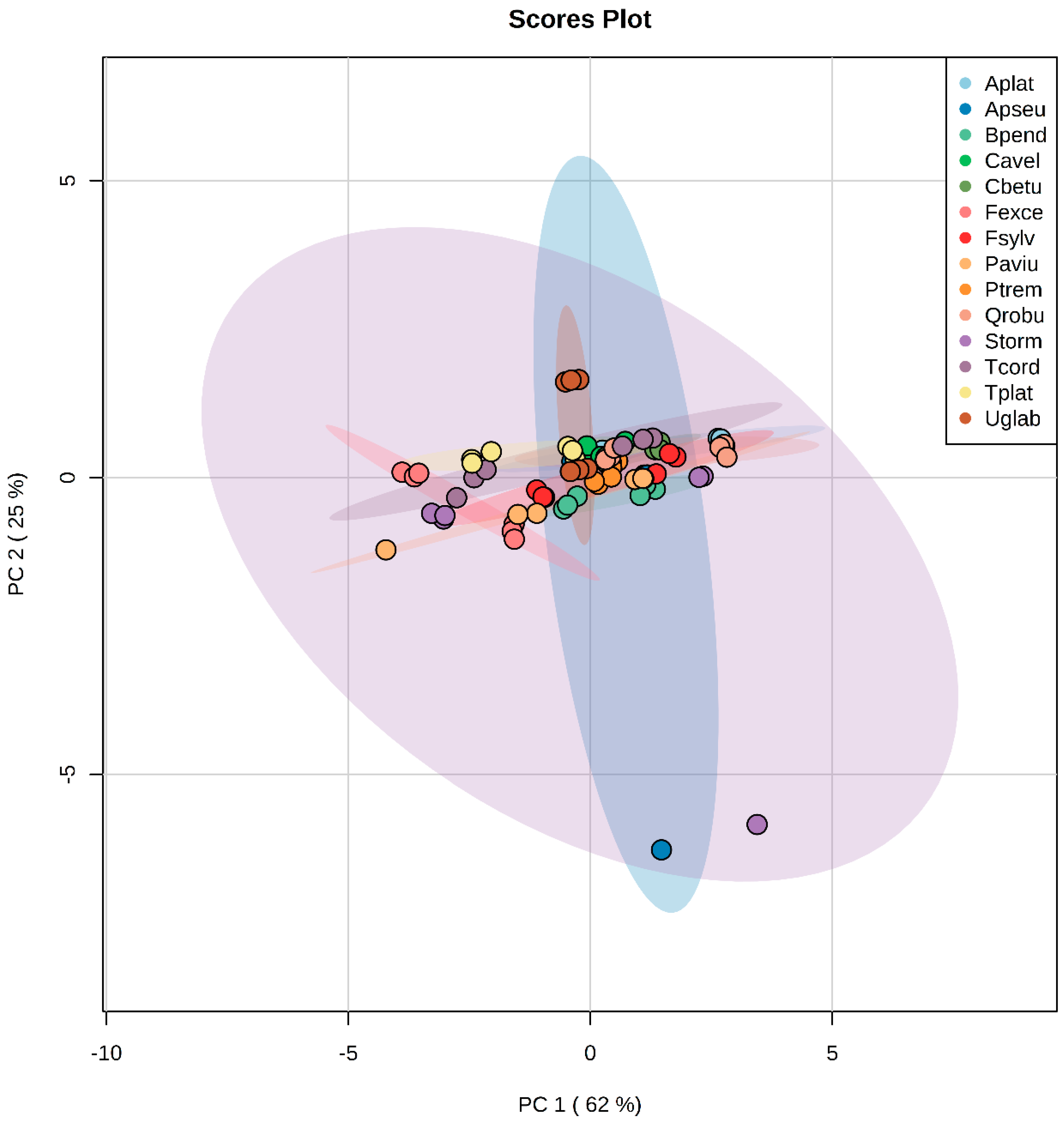
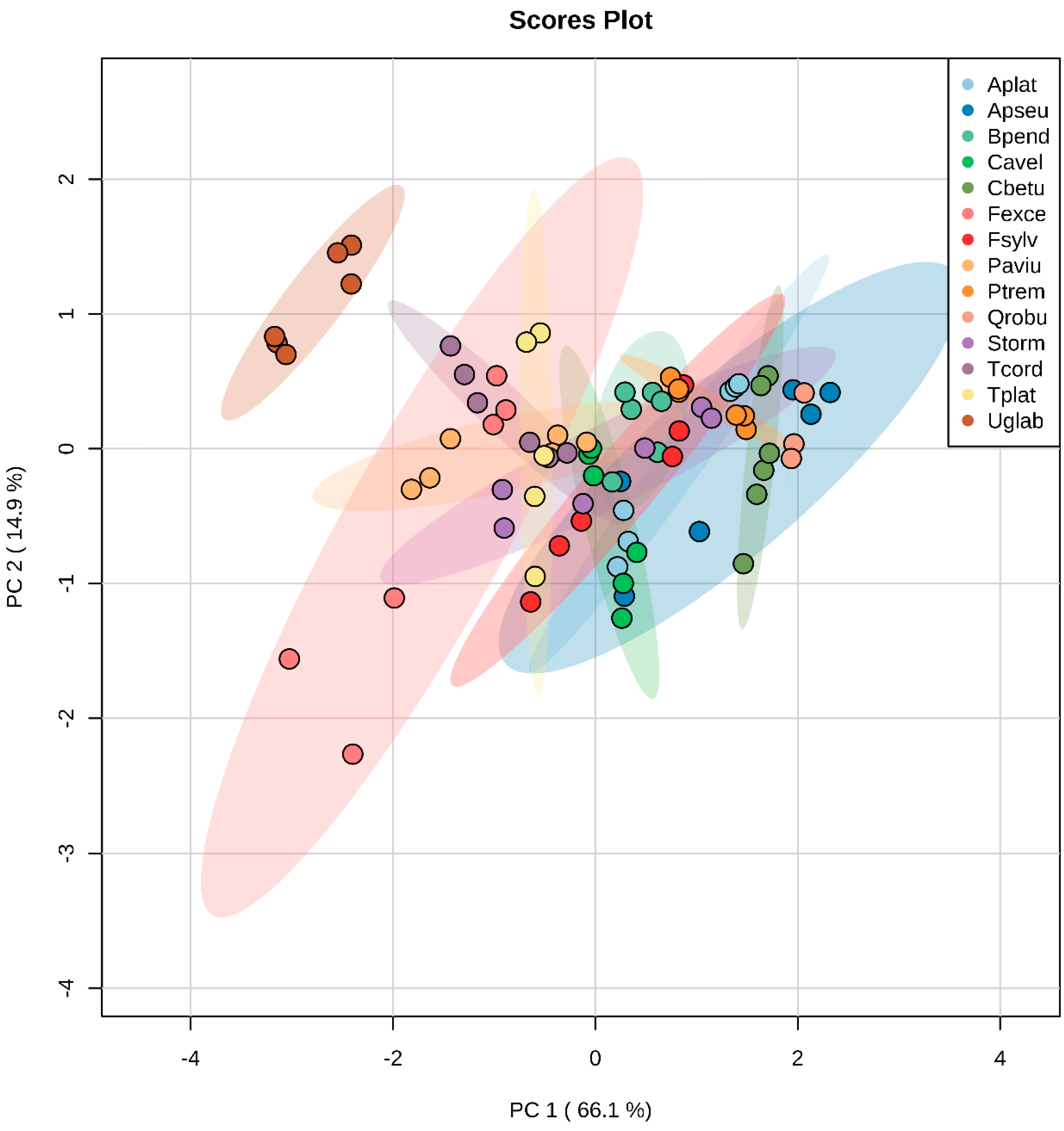
3.1.3. Principal Component Analysis in Green and Litter Leaves
4. Discussion
4.1. No General Patterns across Metabolite Profiles in Leaf Litter and Green Leaves between Species
4.2. Variation between Metabolites in Leaf Litter between Different Temperate Woody Species
4.3. Variation between Metabolites in Green Leaves between Different Temperate Woody Species
4.4. Metabolite Levels in Green Leaves and Leaf Litter Differ between Years
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Parton, W.; Silver, W.L.; Burke, I.C.; Grassens, L.; Harmon, M.E.; Currie, W.S.; King, J.Y.; Adair, E.C.; Brandt, L.A.; Hart, S.C.; et al. Global-scale similarities in nitrogen release patterns during long-term decomposition. Science 2007, 315, 361–364. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Cornelissen, J.H.C.; Amatangelo, K.; Dorrepaal, E.; Eviner, V.T.; Godoy, O.; Hobbie, S.E.; Hoorens, B.; Kurokawa, H.; Pérez-Harguindeguy, N.; et al. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol. Lett. 2008, 11, 1065–1071. [Google Scholar] [CrossRef]
- Schmidt, M.W.I.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kögel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef]
- Garcia-Palacios, P.; Shaw, E.A.; Wall, D.H.; Hattenschwiler, S. Temporal dynamics of biotic and abiotic drivers of litter decomposition. Ecol. Lett. 2016, 19, 554–563. [Google Scholar] [CrossRef]
- Aber, J.D.; Melillo, J.M.; McClaugherty, C.A. Predicting long-term patterns of mass loss, nitrogen dynamics, and soil organic matter formation from initial fine litter chemistry in temperate forest ecosystems. Can. J. Bot. 1990, 68, 2201–2208. [Google Scholar] [CrossRef]
- Gholz, H.L.; Wedin, D.A.; Smitherman, S.M.; Harmon, M.E.; Parton, W.J. Long-term dynamics of pine and hardwood litter in contrasting environments: Toward a global model of decomposition. Glob. Change Biol. 2000, 6, 751–765. [Google Scholar] [CrossRef]
- Horner, J.D.; Gosz, J.R.; Cates, R.G. The role of carbon-based plant secondary metabolites in decomposition in terrestrial ecosystems. Am. Nat. 1988, 132, 869–883. [Google Scholar] [CrossRef]
- Kraus, T.E.C.; Dahlgren, R.A.; Zasoski, R.J. Tannins in nutrient dynamics of forest ecosystems—A review. Plant Soil 2003, 256, 41–66. [Google Scholar] [CrossRef]
- Aerts, R.; Chapin, F.S., III. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns. Adv. Ecol. Res. 2000, 30, 1–67. [Google Scholar] [CrossRef]
- Güsewell, S.; Gessner, M.O. N:P ratios influence litter decomposition and colonization by fungi and bacteria in microcosms. Funct. Ecol. 2009, 23, 211–219. [Google Scholar] [CrossRef]
- Hättenschwiler, S.; Coq, S.; Barantal, S.; Handa, I.T. Leaf traits and decomposition in tropical rainforest: Revisiting some commonly held views towards a new hypothesis. New Phytol. 2011, 189, 950–965. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-Y.; Lu, X.-T.; Hartmann, H.; Keller, A.; Han, X.-G.; Trumbore, S.; Phillips, R.P. Foliar nutrient resorption differs between arbuscular mycorrhizal and ectomycorrhizal trees at local and global scale. Glob. Ecol. Biogeogr. 2018, 27, 875–885. [Google Scholar] [CrossRef]
- Horwath, W. Carbon Cycling: The dynamics and formation of organic matter in soil microbiology, ecology, and biochemistry. In Soil Microbiology, Ecology and Biochemistry, 4th ed.; Paul, E.A., Ed.; Academic Press: Boston, MA, USA, 2015; pp. 339–382. ISBN 9789124159556. [Google Scholar]
- Chomel, M.; Guittonny-Larcheveque, M.; Fernandez, C.; Gallet, C.; Desrochers, A.; Paré, D.; Jackson, B.G.; Baldy, V. Plant secondary metabolites: A key driver of litter decomposition and nutrient cycling. J. Ecol. 2016, 104, 1527–1541. [Google Scholar] [CrossRef]
- Adamczyk, B.; Simon, J.; Kitunen, V.; Adamczyk, S.; Smolander, A. Tannins and their complex interaction with different organic nitrogen compounds and enzymes: Old paradigms versus recent advances. ChemistryOpen 2017, 6, 610–614. [Google Scholar] [CrossRef] [PubMed]
- Suseela, V.; Tharayil, N. Decoupling the direct and indirect effects of climate on plant litter decomposition: Accounting for stress-induced modifications in plant chemistry. Glob. Change Biol. 2018, 24, 1428–1451. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.A.; Warren, R.J., II; Baldrian, P.; Crowther, T.W.; Maynard, D.S.; Oldfield, E.E.; Wieder, W.R.; Wood, S.A.; King, J.R. Climate fails to predict wood decomposition at regional scales. Nat. Clim. Chang. 2014, 4, 625–630. [Google Scholar] [CrossRef]
- Bradford, M.A.; Berg, B.; Maynard, D.S.; Wieder, W.R.; Wood, S.A. Understanding the dominant controls on litter decomposition. J. Ecol. 2016, 104, 229–238. [Google Scholar] [CrossRef]
- Ehrenfeld, J.G.; Ravit, B.; Elgersma, K. Feedback in the plant-soil system. Annu. Rev. Environ. Resour. 2005, 30, 75–115. [Google Scholar] [CrossRef]
- Wurzburger, N.; Hendrick, R.L. Plant litter chemistry and mycorrhizal roots promote a nitrogen feedback in temperate forests. J. Ecol. 2009, 97, 528–536. [Google Scholar] [CrossRef]
- Craine, J.M.; Morrow, C.; Fierer, N. Microbial nitrogen limitation increases decomposition. Ecology 2007, 88, 2105–2113. [Google Scholar] [CrossRef]
- Knicker, H. Soil organic N—An under-rated player for C sequestration in soils? Soil Biol. Biochem. 2011, 43, 1118–1129. [Google Scholar] [CrossRef]
- Sariyildiz, T.; Anderson, J.M. Interactions between litter quality, decomposition, and soil fertility: A laboratory study. Soil Biol. Biochem. 2003, 35, 391–399. [Google Scholar] [CrossRef]
- Sariyildiz, T.; Anderson, J.M. Variation in the chemical composition of green leaves and leaf litters from three deciduous tree species growing in different soil types. For. Ecol. Manag. 2005, 210, 303–319. [Google Scholar] [CrossRef]
- Nordell, K.O.; Karlsson, P.S. Resorption of nitrogen and dry matter prior to leaf abscission: Variation among individuals, sites and years in the mountain birch. Funct. Ecol. 1995, 9, 326–333. [Google Scholar] [CrossRef]
- Hagen-Thorn, A.; Varnagiryte, I.; Nihlgard, B.; Armolaitis, K. Autumn nutrient resorption and losses in four deciduous tree species. For. Ecol. Manag. 2006, 228, 33–39. [Google Scholar] [CrossRef]
- Hättenschwiler, S.; Bracht-Jorgensen, H. Carbon quality rather than stoichiometry controls litter decomposition in a tropical rain forest. J. Ecol. 2010, 98, 754–763. [Google Scholar] [CrossRef]
- Coq, S.; Souquet, J.M.; Meudec, E.; Cheynier, V.; Hättenschwiler, S. Interspecific variation in leaf litter tannins drives decomposition in a tropical rain forest of French Guiana. Ecology 2010, 91, 2080–2091. [Google Scholar] [CrossRef]
- Jacob, M.; Weland, N.; Platner, C.; Schaefer, M.; Leuschner, C.; Thomas, F.M. Nutrient release from decompositing leaf litter of temperate deciduous forest trees along a gradient of increasing tree species diversity. Soil Biol. Biochem. 2009, 41, 2122–2130. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Reich, P.B.; Oleksyn, J.; Ogdahl, M.; Zytkowiak, R.; Hale, C.; Karolewski, P. Tree species effects on decomposition and forest floor dynamics in a common garden. Ecology 2006, 87, 2288–2297. [Google Scholar] [CrossRef]
- Skorupski, M.; Jagodzinski, A.M.; Zytkowiak, R.; Karolewski, P. Differences in chemical composition of needle and leaf litter from exotic and native tree species stands. Dendrobiology 2012, 68, 101–112. [Google Scholar]
- Cotrufo, M.F.; Ineson, P. Elevated CO2 reduces field decomposition rates of Betula pendula (Roth.) leaf litter. Oecologia 1996, 106, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, K.; Preston, C.M.; Krumrei, S.; Feger, K.-H. Decomposition of needle/leaf litter from Scots pine, black cherry, common oak and European beech at a conurbation site. Eur. J. For. Res. 2004, 123, 177–188. [Google Scholar] [CrossRef]
- Top, S.M.; Filley, T.R. Effects of elevated CO2 on the extractable amino acids of leaf litter and fine roots. New Phytol. 2014, 202, 1257–1265. [Google Scholar] [CrossRef] [PubMed]
- Harris, C.W.; Silvester, E.; Rees, G.N.; Pengelly, J.; Puskar, L. Proteins are a major component of dissolved organic nitrogen (DON) leached from terrestrially aged Eucalyptus camaldulensis leaves. Environ. Chem. 2016, 13, 877–887. [Google Scholar] [CrossRef]
- Rawlins, A.J.; Bull, I.D.; Poirier, N.; Ineson, P.; Evershed, R.P. The biochemical transformation of oak (Quercus robur) leaf litter consumed by the pill millipede (Glomeris marginata). Soil Biol. Biochem. 2006, 38, 1063–1076. [Google Scholar] [CrossRef]
- Hicks, R.E.; Lee, C.; Marinucci, A.C. Loss and recycling of amino acids and protein from smooth cordgrass (Spartina alterniflora) litter. Estuaries 1991, 14, 430–439. [Google Scholar] [CrossRef]
- Ellenberg, H.; Leuschner, C. Vegetation Mitteleuropas mit den Alpen in Ökologischer, Dynamischer und Historischer SICHT, 6th ed.; Ulmer Verlag: Stuttgart, Germany, 2014; p. 1333. ISBN 9783825281045. [Google Scholar]
- Kiermeier, P. Die Lebensbereiche der Gehölze Eingeteilt nach dem Kennziffernsystem, 3rd ed.; Verlagsgesellschaft Grün ist Leben mbH: Pinneberg, Germany, 1993; p. 31. ISBN 38781510047. [Google Scholar]
- Professur für Waldbau und Professur für Forstschutz und Dendrologie der ETH Zürich. Mitteleuropäische Waldbaumarten; Artbeschreibung und Ökologie unter Besonderer Berücksichtigung der Schweiz; ETH-Zürich: Zürich, Switzerland, 2002. [Google Scholar]
- Grüning, M.; Simon, J.; Rennenberg, H.; l-M-Arnold, A. Defoliating insect mass infestation affects soil N fluxes and tree N nutrition in Scots pine forests. Front. Plant Sci. 2017, 8, 954. [Google Scholar] [CrossRef]
- Winter, H.; Lohaus, G.; Heldt, H.W. Phloem transport of amino acids in relation to their cytosolic levels in barley leaves. Plant Physiol. 1992, 99, 996–1004. [Google Scholar] [CrossRef]
- Li, X.; Rennenberg, H.; Simon, J. Seasonal variation in N uptake strategies in the understorey of a beech-dominated N-limited forest ecosystem depends on N source and species. Tree Physiol. 2016, 36, 589–600. [Google Scholar] [CrossRef]
- Liu, X.-P.; Grams, T.E.E.; Matyssek, R.; Rennenberg, H. Effects of elevated pCO2 and/or pO3 on C-, N-, and S-metabolites in the leaves of juvenile beech and spruce differ between trees grown in monoculture and mixed culture. Plant Physiol. Biochem. 2005, 43, 147–154. [Google Scholar] [CrossRef]
- Simon, J.; Gleadow, R.M.; Woodrow, I.E. Allocation of nitrogen to chemical defence and plant functional traits is constrained by soil N. Tree Physiol. 2010, 30, 1111–1117. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Psychogios, N.; Young, N.; Wishart, D.S. MetaboAnalyst: A web server for metabolomic data analysis and interpretation. Nucleic Acids Res. 2009, 37, W652–W660. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wishart, D.S. Metabolomic data processing, analysis, and interpretation using MetaboAnalyst. Curr. Protoc. Bioinform. 2011, 14.10.1–14.10.48. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wishart, D.S. Web-based inference of biological patterns, functions and pathways from metabolomic data using MetaboAnalyst. Nat. Prococ. 2011, 6, 743–760. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Mandal, R.; Sinelnikov, I.; Broadhurst, D.; Wishart, D.S. MetaboAnalyst 2.0—A comprehensive server for metabolomic data analysis. Nucleic Acids Res. 2012, 40, W127–W133. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Sinelnikov, I.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0—Making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. Using MetaboAnalyst 3.0 for comprehensive metabolomics data analysis. Curr. Protoc. Bioinform. 2016, 55, 14.10.1–14.10.91. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018. [Google Scholar] [CrossRef]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S. From tropics to tundra: Global convergence in plant functioning. Proc. Natl. Acad. Sci. USA 1997, 94, 13730–13734. [Google Scholar] [CrossRef]
- Grime, J.P.; Hunt, R. Relative growth rate: Its range and adaptive significance in a local flora. J. Ecol. 1975, 63, 393–422. [Google Scholar] [CrossRef]
- Lambers, H.; Poorter, H.; van Vuuren, M. Research on variation in plant growth rate—Introduction. In Inherent Variation in Plant Growth. Physiological Mechanism and Ecological Consequences; Lambers, H., Poorter, H., van Vuuren, M., Eds.; Backhuys Publishers: Leiden, The Netherlands, 1998; pp. 1–4. ISBN 90-73348-96-X. [Google Scholar]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Poorter, H.; Garnier, E. Ecological significance of inherent variation in relative growth rate and its components. In Handbook of Functional Plant Ecology; Pugnaire, F.I., Valladares, F., Eds.; M Dekker: New York, NY, USA, 1999; pp. 81–120. ISBN 9780849374883. [Google Scholar]
- Reich, P.B.; Buschena, C.; Tjoelker, M.G.; Wrage, K.; Knops, J.; Tilman, D.; Machado, J.L. Variation in growth rate among 34 grassland and savanna species under contrasting N-supply: A test of functional group differences. New Phytol. 2003, 157, 617–631. [Google Scholar] [CrossRef]
- Langenbruch, C.; Helfrich, M.; Flessa, H. Effects of beech (Fagus sylvatica), ash (Fraxinus excelsior) and lime (Tilia spec.) on soil chemical properties in a mixed deciduous forest. Plant Soil 2012, 352, 389–403. [Google Scholar] [CrossRef]
- Zhou, G.; Zhang, J.; Qiu, X.; Wie, F.; Xu, X. Decomposing litter and associated microbial activity responses to nitrogen deposition in two subtropical forests containing nitrogen-fixing or non-nitrogen-fixing tree species. Sci. Rep. 2018, 8, 12934. [Google Scholar] [CrossRef] [PubMed]
- Cotrufo, M.F.; Miller, M.; Zeller, B. Litter decomposition. In Ecological Studies 142—Carbon and Nitrogen Cycling in European Forest Ecosystems; Schulze, E.-D., Ed.; Springer-Verlag: Berlin/Heidelberg, Germany, 2000; pp. 276–296. ISBN 3-540-67239-7. [Google Scholar]
- Austin, A.T.; Vivanco, L.; González-Arzac, A.; Pérez, L.I. There’s no place like home? An exploration of the mechanisms behind plant litter-decomposer affinity in terrestrial ecosystems. New Phytol. 2014, 204, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Handa, I.T.; Aerts, R.; Berendse, F.; Berg, M.P.; Bruder, A.; Butenschoen, O.; Chauvet, E.; Gessner, M.O.; Jabiol, J.; Makkonen, M.; et al. Consequences of biodiversity loss for litter decomposition across biomes. Nature 2014, 509, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Barbe, L.; Jung, V.; Prinzing, A.; Bittebiere, A.-K.; Butenschoen, O.; Mony, C. Functionally dissimilar neighbors accelerate litter decomposition in two grass species. New Phytol. 2017, 214, 1092–1102. [Google Scholar] [CrossRef] [PubMed]
- Eviner, V.T.; Chapin, F.S.; Vaughn, C.E. Seasonal variation in plant species effects on soil N and P dynamics. Ecology 2006, 87, 974–986. [Google Scholar] [CrossRef]
- Vivanco, L.; Rascovan, N.; Austin, A.T. Plant, fungal, bacterial, and nitrogen interactions in the litter layer of a native Patagonian forest. PeerJ 2018, 6, e4754. [Google Scholar] [CrossRef]
- Butenschoen, O.; Scheu, S. Climate change triggers effects of fungal pathogens and insect herbivores on litter decomposition. Acta Oecol. 2014, 60, 49–56. [Google Scholar] [CrossRef]
- Tagliavini, M.; Quartieri, M.; Millard, P. Remobilised nitrogen and root uptake of nitrate for spring leaf growth, flowers and developing fruits of pear (Pyrus communis L.) trees. Plant Soil 1997, 196, 137–142. [Google Scholar] [CrossRef]
- Millard, P. Ecophysiology of the internal cycling of nitrogen for tree growth. Z. Pflanzenern. Bodenk. 1996, 159, 1–10. [Google Scholar] [CrossRef]
- Dyckmans, J.; Flessa, H. Influence of tree internal N status on uptake and translocation of C and N in beech: A dual 13C and 15N labelling approach. Tree Physiol. 2001, 21, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Fotelli, M.N.; Rienks, M.; Rennenberg, H.; Gessler, A. Climate and forest management affect 15N-uptake, N balance and biomass of European beech seedlings. Trees 2004, 18, 157–166. [Google Scholar] [CrossRef]
- Simon, J.; Dannenmann, M.; Gasche, R.; Holst, J.; Mayer, H.; Papen, H.; Rennenberg, H. Competition for nitrogen between adult European beech and its offspring is reduced by avoidance strategy. For. Ecol. Manag. 2011, 262, 105–114. [Google Scholar] [CrossRef]
- Millard, P.; Grelet, G.-A. Nitrogen storage and remobilization by trees: Ecophysiological relevance in a changing world. Tree Physiol. 2010, 30, 1083–1095. [Google Scholar] [CrossRef] [PubMed]
- Ruuhola, T.; Ossipov, V.; Lempa, K.; Haukioja, E. Amino acids during leaf development of mountain birch leaves. Chemoecology 2003, 13, 95–101. [Google Scholar] [CrossRef]
- Jeong, M.L.; Jiang, H.; Chen, H.-S.; Tsai, C.-J.; Harding, S.A. Metabolomic profiling of the sink-to-source transition in developing leaves of quaking aspen. Plant Physiol. 2004, 136, 3364–3375. [Google Scholar] [CrossRef]
- Tegelberg, R.; Julkunen-Tiitto, R.; Vartiainen, M.; Paunonen, R.; Rousi, M.; Kellomäki, S. Exposures to elevated CO2, elevated temperature and enhanced UV-B radiation modify activities of polyphenol oxidase and guaiacol peroxidase and concentrations of chlorophylls, polyamines and soluble proteins in the leaves of Betula pendula seedlings. Environ. Exp. Bot. 2008, 62, 308–315. [Google Scholar] [CrossRef]
- Zhang, L.; Tu, Z.C.; Yuan, T.; Ma, H.; Niesen, D.B.; Wang, H.; Seeram, N.P. New Gallotannin and other Phytochemicals from Sycamore Maple (Acer pseudoplatanus) Leaves. Nat. Prod. Commun. 2015, 10, 1977–1980. [Google Scholar]
- Chernyak, E.I.; Yushkova, Y.A.; Pavlushin, S.V.; Nikolenko, S.O.; Matem’yanov, V.V.; Morozov, S.V. Dynamics of biologically active compound contents from Betula pendula leaves during early leaf development. Chem. Nat. Compd. 2016, 52, 193–198. [Google Scholar] [CrossRef]
- Hofmann, T.; Nebehaj, E.; Albert, L. Antioxidant properties and detailed polyphenol profiling of European hornbeam (Carpinus betulus L.) leaves by multiple antioxidant capacity assays and high-performance liquid chromatograph/multistage electrospray mass spectrometry. Ind. Crops Prod. 2016, 87, 340–349. [Google Scholar] [CrossRef]
- Riethmüller, E.; Tóth, G.; Alberti, A.; Végh, K.; Burlini, I.; Könczöl, A.; Balogh, G.T.; Kéry, A. First characterisation of flavonoid- and diarylheptanoid-type antioxidant phenolics in Corylus maxima by HPLC-DAD-ESI-MS. J. Pharm. Biomed. Anal. 2015, 107, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Riethmüller, E.; Alberti, A.; Tóth, G.; Béni, S.; Ortolano, F.; Kéry, A. Characterisation of diarylheptanoid- and flavonoid-type phenolics in Corylus avellana L. leaves and bark by HPLC/DAD-ESI/MS. Phytochem. Anal. 2013, 24, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Amaral, J.S.; Ferreres, F.; Andrade, P.B.; Valentao, P.; Pinheiro, C.; Santos, A.; Seabra, R. Phenolic profile of hazelnut (Corylus avellana L.) leaves cultivars grown in Portugal. Nat. Prod. Res. 2005, 19, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Tharayil, N.; Suseela, V.; Triebwasser, D.J.; Preston, C.M.; Gerard, P.D.; Dukes, J.S. Changes in the structural composition and reactivity of Acer rubrum leaf litter tannins exposed to warming and altered precipitation: Climatic stress-induced tannins are more reactive. New Phytol. 2011, 191, 132–145. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, E.G.; Johnson, D.W.; Ledford, J.; Todd, D.E. Acute seasonal drought does not permanently alter mass loss and nitrogen dynamics during decomposition of red maple (Acer rubrum L.) litter. Glob. Change Biol. 2003, 9, 117–123. [Google Scholar] [CrossRef]
- Zimmer, M.; Auge, H.; von Wühlisch, G.; Schueler, S.; Haase, J. Environment rather than genetic background explains intraspecific variation in the protein-precipitating capacity of phenolic compounds in beech litter. Plant Ecol. Divers. 2015, 8, 73–79. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Relative Growth Rate | Nutrient Demand | Drought Resistance | Shade Tolerance |
|---|---|---|---|---|
| Acer platanoides | fast | medium/high | high | high |
| Acer pseudoplatanus | fast | medium/high | high | medium |
| Betula pendula | fast | low | medium | low |
| Carpinus betulus | slow | medium | medium | medium |
| Corylus avellana | medium | medium/high | medium | medium/high |
| Fagus sylvatica | slow | low/medium | high | high |
| Fraxinus excelsior | fast | medium/high | medium | low/medium |
| Populus tremula | fast | low | medium | low |
| Prunus avium | fast | medium/high | high | low |
| Quercus robur | medium/fast | low | medium | low |
| Sorbus torminalis | slow | high | medium/high | medium |
| Tilia cordata | slow | medium | high | high |
| Tilia platyphyllos | slow | medium/high | high | high |
| Ulmus glabra | fast | high | high | medium |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simon, J.; Dörken, V.M.; L.-M.-Arnold, A.; Adamczyk, B. Environmental Conditions and Species Identity Drive Metabolite Levels in Green Leaves and Leaf Litter of 14 Temperate Woody Species. Forests 2018, 9, 775. https://doi.org/10.3390/f9120775
Simon J, Dörken VM, L.-M.-Arnold A, Adamczyk B. Environmental Conditions and Species Identity Drive Metabolite Levels in Green Leaves and Leaf Litter of 14 Temperate Woody Species. Forests. 2018; 9(12):775. https://doi.org/10.3390/f9120775
Chicago/Turabian StyleSimon, Judy, Veit M. Dörken, Anne L.-M.-Arnold, and Bartosz Adamczyk. 2018. "Environmental Conditions and Species Identity Drive Metabolite Levels in Green Leaves and Leaf Litter of 14 Temperate Woody Species" Forests 9, no. 12: 775. https://doi.org/10.3390/f9120775
APA StyleSimon, J., Dörken, V. M., L.-M.-Arnold, A., & Adamczyk, B. (2018). Environmental Conditions and Species Identity Drive Metabolite Levels in Green Leaves and Leaf Litter of 14 Temperate Woody Species. Forests, 9(12), 775. https://doi.org/10.3390/f9120775