1. Introduction
Brutian pine is the most important tree species in Turkey, both ecologically and economically. Brutian pine forests cover about 25% of Turkey’s total forest area which is about 5.6 million hectares with a current standing volume of approximately 270 million m
3 [
1]. Because of its valuable wood properties, it is one of the most important pine species for the forest products industry in Turkey [
2]. Furthermore, brutian pine plays a key role in providing important benefits and environmental services such as protection of soil and water resources, conservation of biological diversity, and climate change mitigation and adaptation in Turkey [
3]. Therefore, detailed information about stand structure, total biomass, or biomass of different tree components is needed for sustainable forest management and harvesting of utilizable potential of the brutian pine forest.
Accurate estimation of tree or forest biomass is a key requirement for calculating biomass energy, carbon sequestration, as well as for studying climate change, forest health, site productivity, and nutrient cycling [
4]. Furthermore, the increasing use of weight or biomass as a measure of forest productivity with ever changing market conditions has heightened the need for accurate estimates of total and component biomass of trees.
The approaches in biomass estimation depend on the scale of analysis, need for detail, user group interest, and purpose of estimation [
5]. Generally, there are three approaches used to estimate total and component biomass. In the first group of methods, total tree and component biomass (e.g., stem, crown, branches, leaves, and bark) are regressed against easily measurable tree attributes such as diameter at breast height (DBH) or DBH and height using linear and nonlinear regression. Such methods, however, do not ensure that the sum of biomass predictions from component models is equal to the biomass prediction from the total aboveground biomass (AGB) model. This issue of additivity can be resolved by fitting component and total biomass equations as a simultaneous system. Such methods, if fitted with the ordinary least squares approach, ignore the inherent correlations among the component models [
6]. Therefore, the second group of methods is a regression-based approach that uses a system of equations to deal with this issue of non-additivity or incompatibility. Different estimation methods have been suggested to ensure the additivity in a system of biomass equations for both linear and nonlinear models. In this framework, seemingly unrelated regression (SUR) and non-linear seemingly unrelated regression (NSUR) have become more popular in recent years [
4,
7]. The third group of methods, fairly new in biomass estimation, predicts the proportions of biomass in each component using generalized linear models such as beta, Dirichlet, and multinomial logistic regression. Predicted proportions are then applied to the observed total AGB [
8] or the total AGB obtained from fitting a separate equation [
9].
Information regarding estimations of total and component tree biomass is currently lacking in Turkey. Four published sources for brutian pine biomass include studies based on a sample of 30 trees from Eastern Mediterranean Region [
10], a sample of 24 trees in north and south Aegean Islands of Greece [
11], a sample of 201 trees from Syria and Lebanon [
12], and a sample of 164 trees in southern of Mediterranean Region of Turkey [
13]. However, two of these studies were conducted outside of Turkey. Allometric equations used in these studies are simple expressions relating tree level biomass to expressions of tree size except for Özçelik et al. [
13]. Common independent variables of biomass models are DBH and tree height, although some studies have used crown dimensions as well. Poudel and Temesgen [
8] indicated that factors that affect growth in tree diameter and height (e.g., genetics, site quality, environmental factors, stand density, tree and stand age) also affect AGB and component biomass. Therefore, there is a need to develop tree-level biomass models using data from areas within the natural distribution of this species in the Mediterranean Region.
Recently, Chaturvedi and Raghubanshi [
14] indicated that woody individuals of small diameter classes have a significant role in the estimation of total AGB, since this component of the forest comprises a significant proportion, by number, of the tree population. These trees are not of significance for volumetric production but can contribute substantially towards biomass and bioenergy as they have a faster growth than the trees in larger diameter classes. The information about small diameter trees can be used in inventories of fuel or wood energy, to assess the potential of young stands as fiber sources and the carbon sequestration potential of natural stands, and as indicators of net primary production [
15]. Additionally, accurate assessment of wildfire behavior requires quantitative estimates of available fuel load by size class and condition in terms of forest management [
16].
Many of the studies concerning biomass estimation have focused solely on the estimation of individual trees having DBH greater than 8 cm, ignoring the AGB of small diameter trees at the sapling stage. As a result, available woody biomass and the carbon stored at early stages are often neglected. Only a few studies address the estimation of small diameter tree biomass in tropical dry forests [
14,
17], in temperate deciduous forest [
15,
18], and in temperate pine forest [
19]. Ideally, the biomass equations should be developed covering all size classes without discontinuity at any tree size.
Small diameter tree biomass estimates in Turkey are limited to a few studies and their predictions are mainly for crown biomass components and are based on a small sample size [
20,
21]. Trees with less than 8 cm DBH are considered small diameter trees in Turkey and are not measured in regular forest inventory applications such as industrial roundwood production. Therefore, there is no reliable information about tree volume and total tree biomass or biomass components, such as stem and branches, for such trees. Reliable small tree biomass models are especially important in fire-prone forest ecosystems such as brutian pine forests in the Mediterranean Region of Turkey, where nearly 15% of the forested area is dominated by sapling sized (0.1 cm–8 cm DBH) stands. The lack of aboveground small diameter tree biomass equations has also affected the accuracy of assessing the amount of utilizable woody biomass, forest fuel inventories, and carbon sequestration potential.
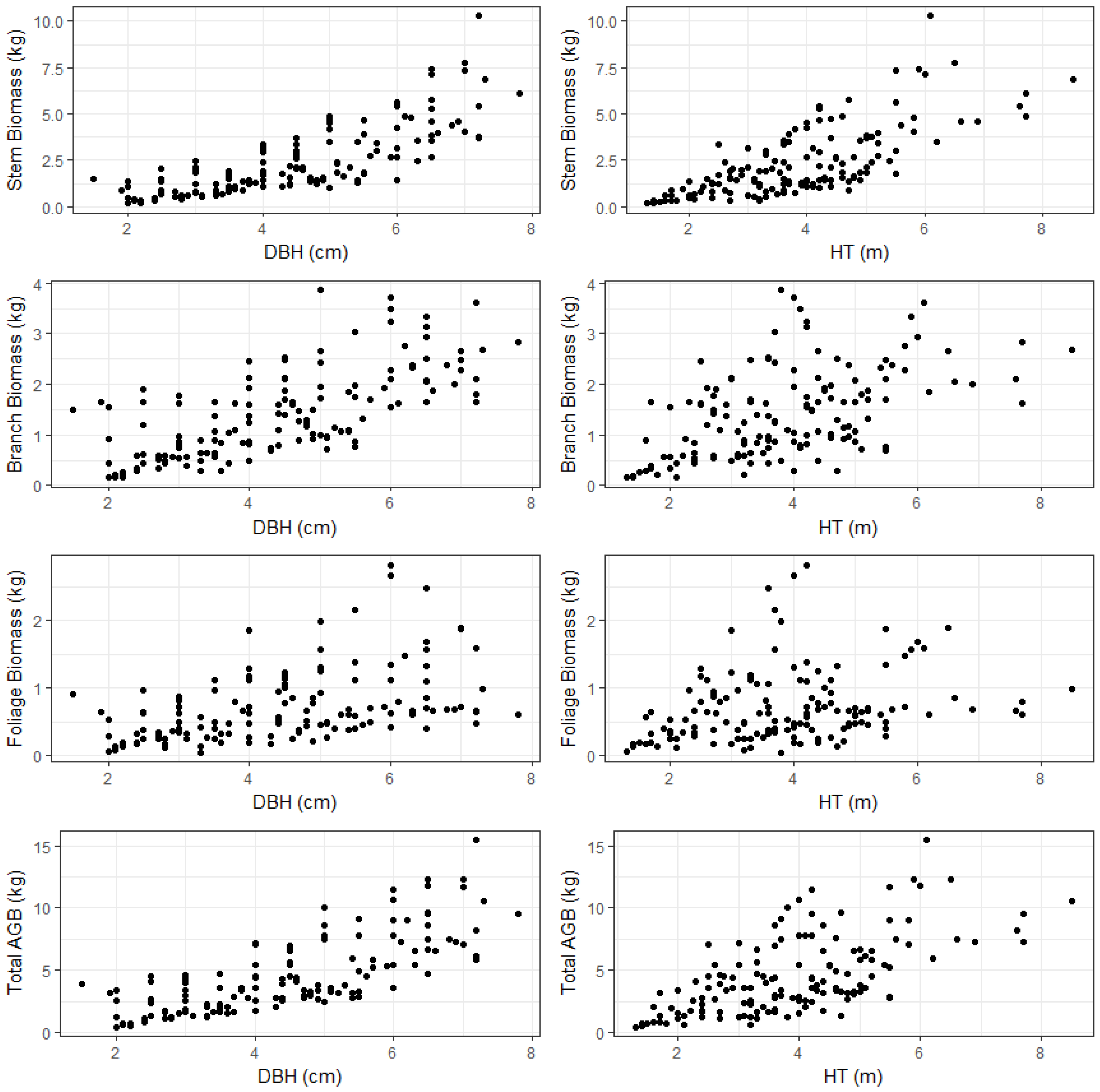
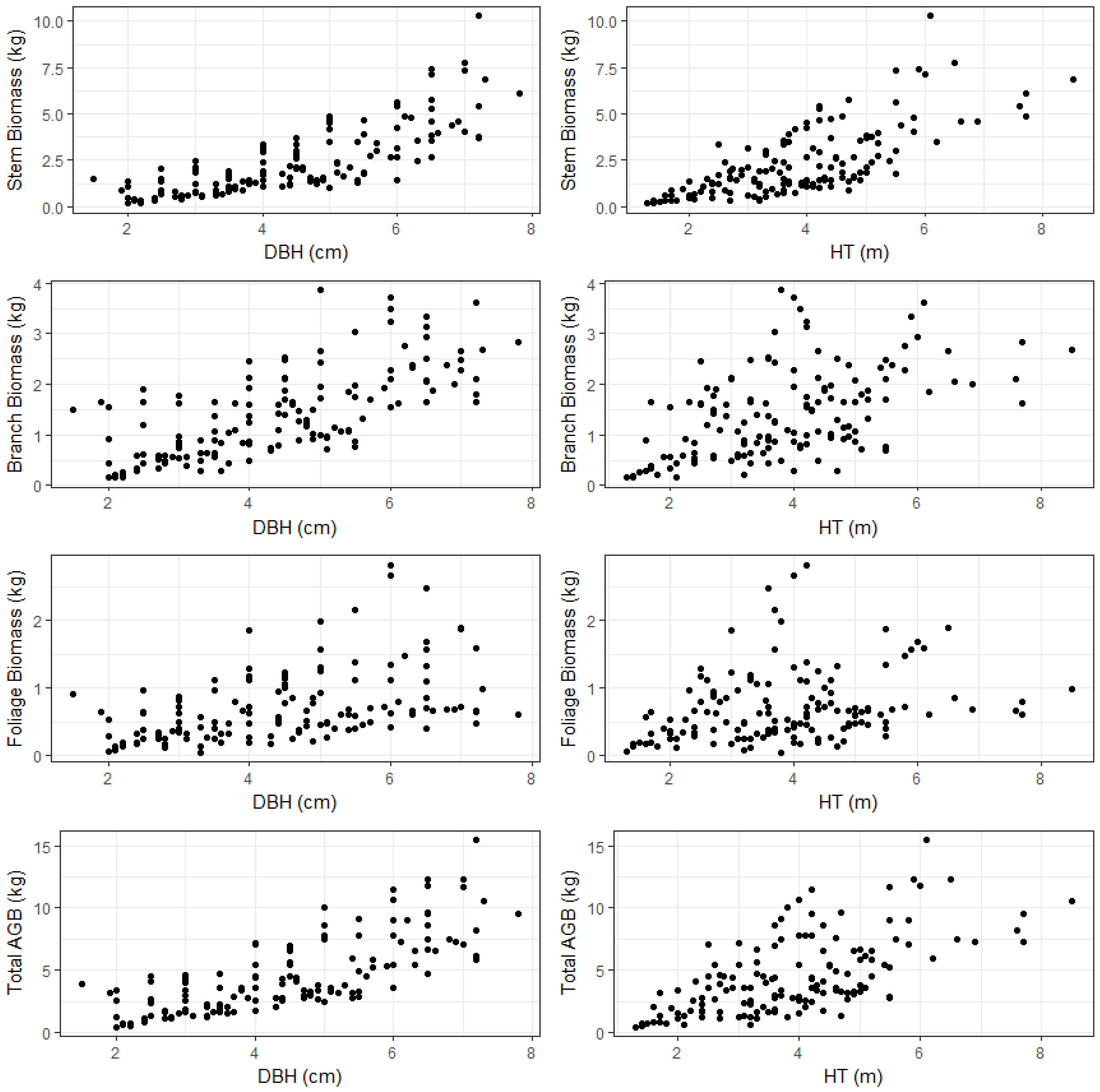
AGB is commonly divided into three major components: stem, stem bark and crown (branch and leaves/foliage). The component biomass models are useful to account for the variability within the tree. In addition, tree component biomass is used for different purposes that require separate estimates. The stem wood is used for industrial wood, chip-board wood, and fuel/energy wood production; crown biomass can provide information on fuel load and wildfire assessment, and woody sections of branches are also useful in bioenergy production. Therefore, component biomass estimates are necessary to determine available forest products within the concept of sustainable management and harvesting of small trees.
In this study, destructive sampling was used to measure the biomass of foliage, branch, and bole (stem) of sapling stage brutian pine in the Mediterranean Region of Turkey. The objective of the study was to develop estimation models for total AGB and component biomass of small diameter brutian pine. Thus, the aim was to estimate the type and amount of biomass that emerged after silvicultural interventions were applied to young and small diameter trees. A nonlinear mixed effects model was fitted to predict total tree biomass as a function of DBH and total tree height accounting for the variation among plots. Predicted total tree biomass was then apportioned into different components according to the predicted proportions from beta, Dirichlet, and multinomial logistic regressions.
3. Results and Discussion
Parameter estimates and their standard errors for the nonlinear mixed effects model used to predict total AGB are given in
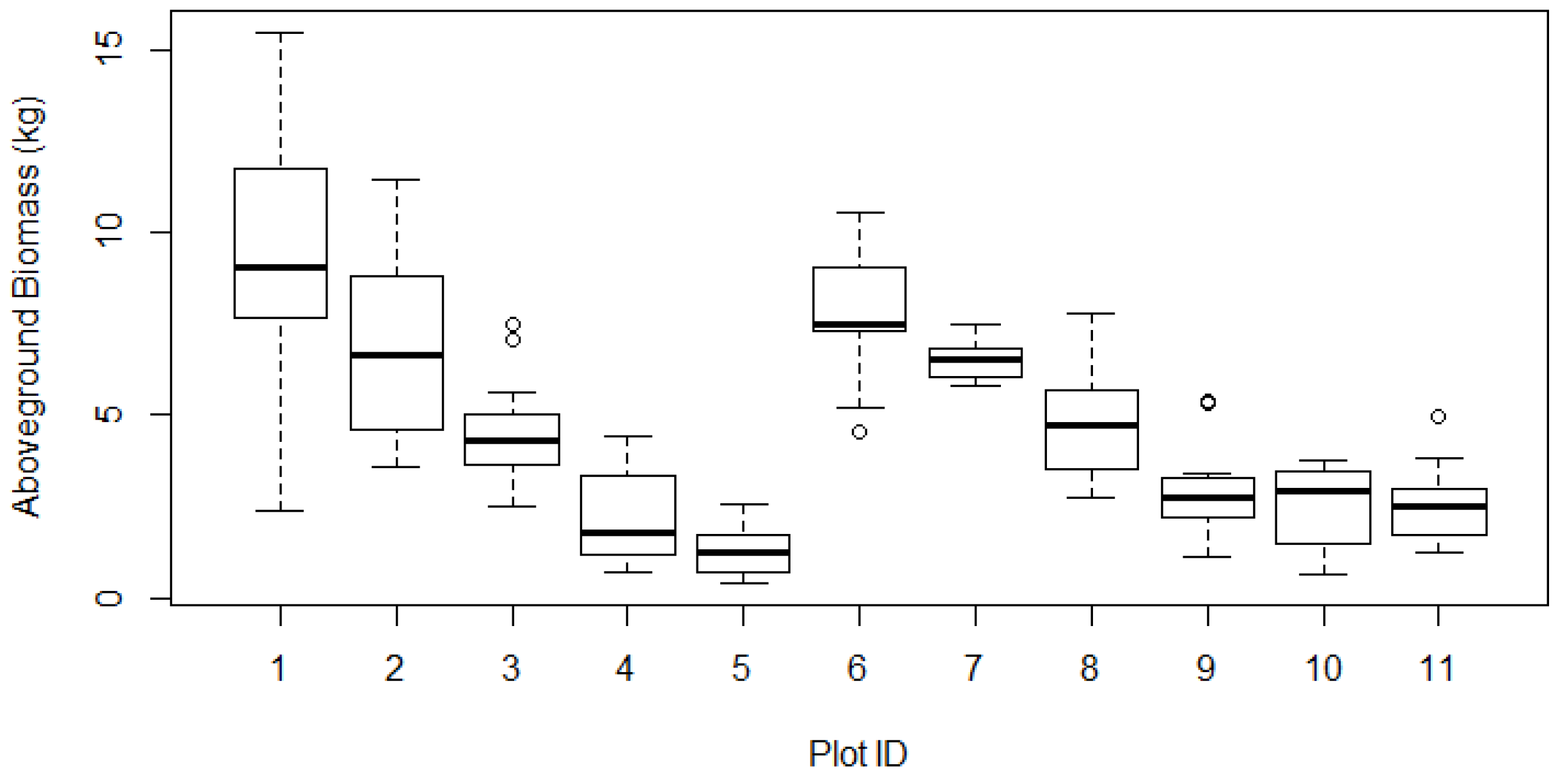
Table 2. The boxplot of AGB in each plot shows the variability in total AGB among 11 plots (
Figure 2).
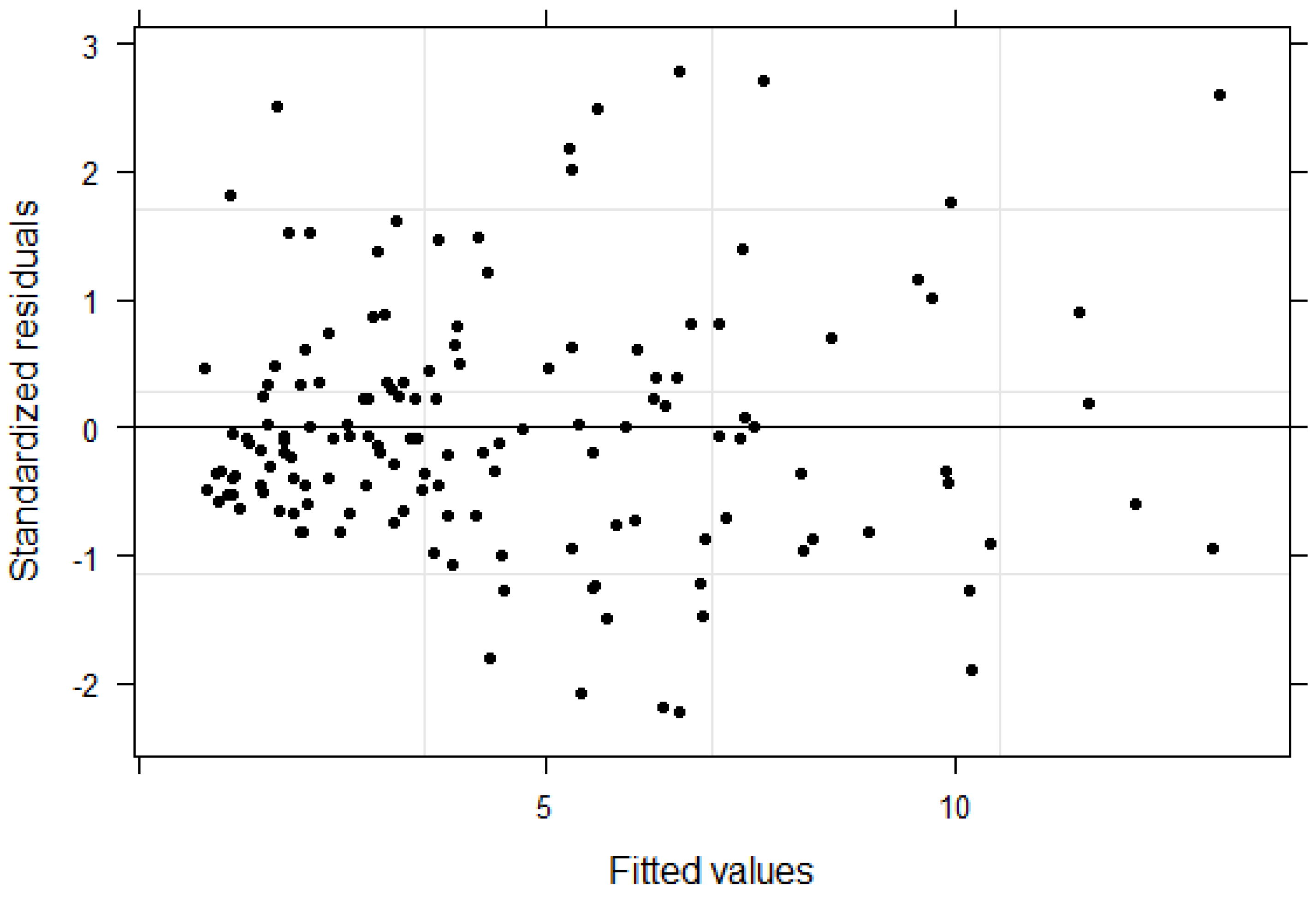
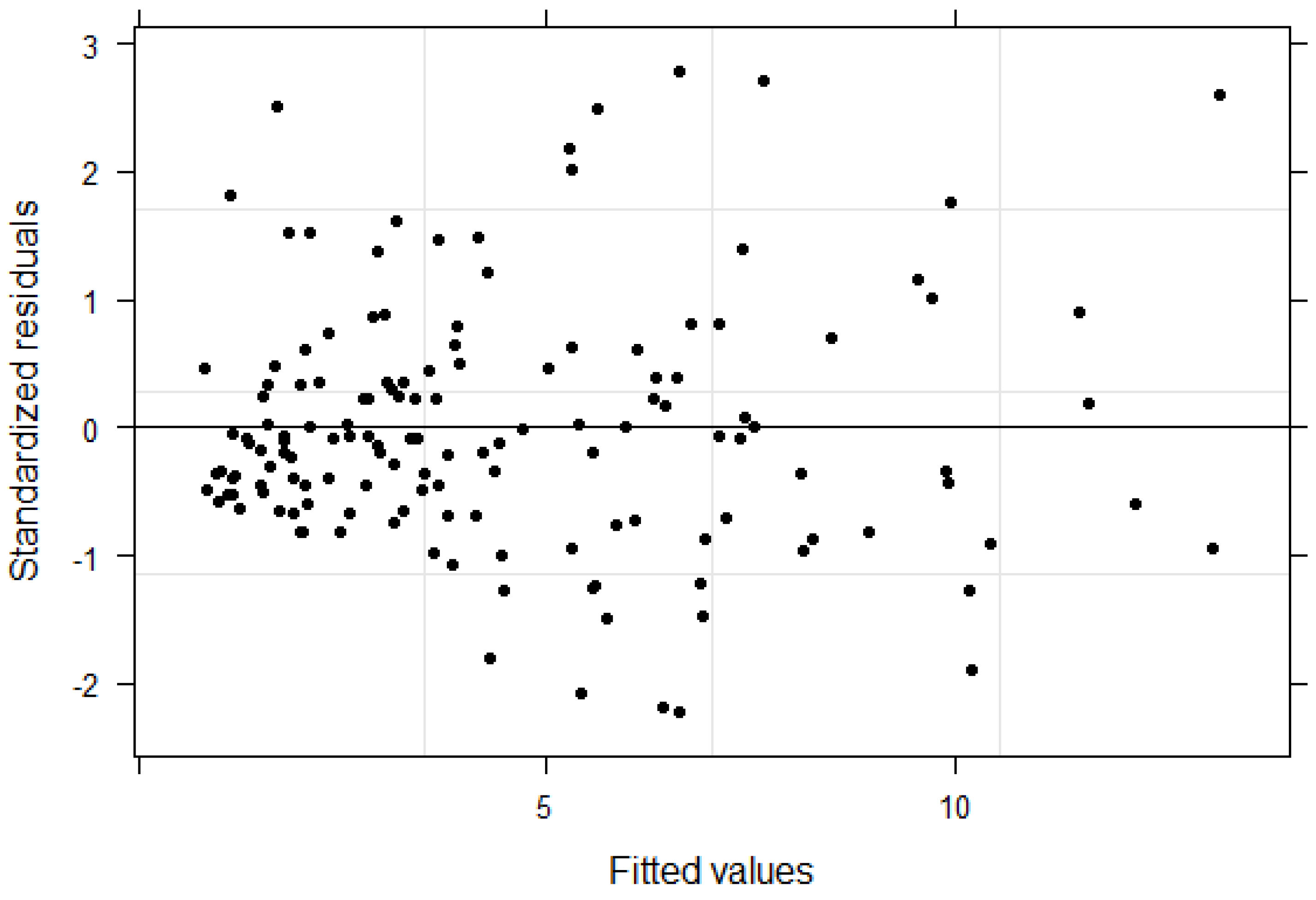
The relationship between AGB and dendrometric variables such as DBH and height varies by stands or plots. Therefore, the mixed effects model was appropriate in our study because it addressed the hierarchical nature of the data by incorporating plot level variation in the model. The bias and RMSE of this model for the modeling data were −0.01 kg and 0.84 kg (−0.17% and 18.84% of mean AGB, respectively). The residual analysis did not show any problems with the model fit (
Figure 3).
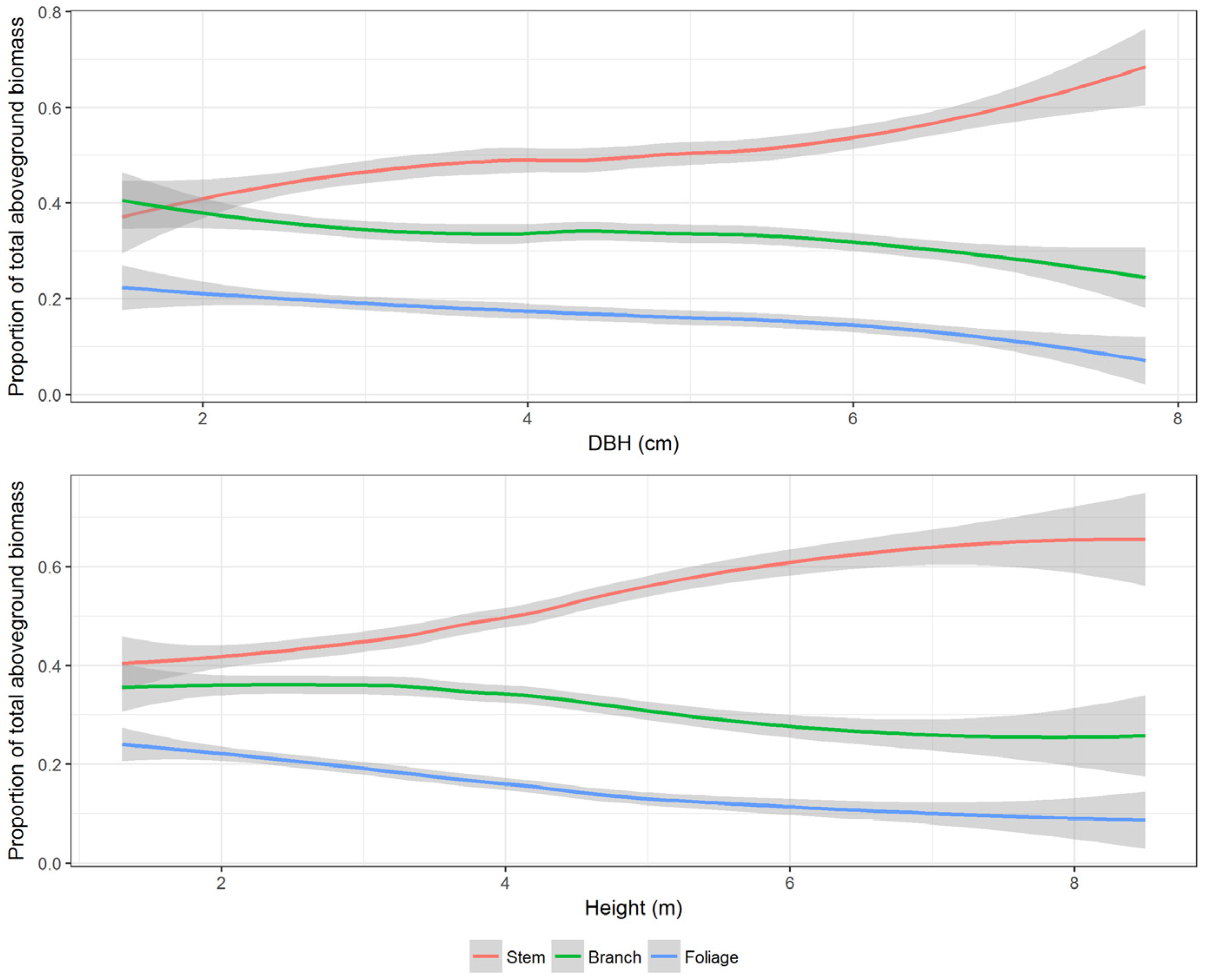
Allocation of component biomass is influenced by various site factors such as stand density, site productivity, competition at the tree level, soil characteristics such as texture and moisture content, and tree characteristics such as species and age [
8]. In mature stands and trees, most of the biomass is contained in the main stem. For our sample trees, stem, branch, and foliage biomass, on average, accounted for 50%, 33%, and 17% of total AGB. Stem biomass ranged from 27% to 68%, branch biomass ranged from 18% to 53%, and foliage biomass ranged from 4% to 29% of total AGB. The proportion of stem biomass increased with increasing DBH and height (
Figure 4). Similar findings for brutian pine were reported by de-Miguel et al. [
12]. They found that the proportion of stem biomass is lower in small or young trees whereas the proportion of crown biomass diminishes as the tree grows. Note that the proportions in compositional data are inversely related, i.e., if the proportion of one component increases, the proportion of the other components decreases—also seen in
Figure 4. Foliage biomass decreased with increasing diameter and height. However, the rate of decline in foliage biomass was higher with increasing height (in meter) than with increasing DBH (in centimeter). This could be because the vertical competition has more effect on crown biomass than the horizontal competition. Branch biomass proportion showed a similar trend as the foliage proportion. After 4 cm DBH and 4 m height, both branch and foliage biomass declined monotonically while the stem biomass increased monotonically. This can have both ecological significance as well as management implications. One such implication is assessing the potential for supplying branch and foliage biomass (the logging residues) for bioenergy production. Kuuluvainen [
37] found that the proportion of stem biomass from total AGB increased from smaller to larger trees and then stabilized. This reflects, in line with the pipe-model theory [
38], the increasing need for biomass allocation into stem at early stages of tree development until a balance between stem and crown biomass accumulation is achieved. The results are consistent with current biological knowledge on stand dynamics of light-demanding species. Brutian pine is managed under even-aged schedules and trees growing in such stands have longer stems and smaller crowns because of the competition for light.
In this study beta, Dirichlet, and multinomial logistic regressions were used to predict the proportion of biomass in stem, branch, and foliage. These are all generalized linear models and were unbiased in predicting biomass proportions. However, the prediction of proportion and error in proportion itself is not as relevant and requires that both estimates and error in biomass units are obtained. This can be obtained by applying predicted proportions to the total biomass obtained from AGB equation. This underscores the importance of developing the best possible model to obtain total AGB because partitioning an inaccurate AGB would provide inaccurate estimates of the component masses as well. On the other hand, accurate models to predict component biomass are essential to meet other purposes such as to assess availability of biomass feedstock. Therefore, biomass in different components was obtained by applying predicted proportions to AGB predicted from the nonlinear mixed effects model.
The beta regression models predicted proportion of biomass in stem, branch, and foliage independently of each other. Parameter estimates and their standard errors of beta regression models are given in
Table 3.
Component models are generally not as good as the model to fit AGB. The model to predict branch biomass proportion had the smaller
pseudo-R2 (0.1827) than the models to predict stem and foliage biomass. This is justified by the flatter smooth line observed in
Figure 4. The stem and foliage proportion models had
pseudo-R2 0.4245 and 0.4780, respectively. The evaluation statistics produced by the beta regression models are given in
Table 4. Branch and stem biomass were over predicted by the beta regression models by 1.62% and 0.25% whereas the foliage biomass was under predicted by 2.93%. RMSEs for the beta models were 39.11%, 31.50%, and 18.77% for foliage, branch, and stem biomass estimation, respectively. Note that, even though the foliage model had a higher
pseudo-R2 value than the branch model, it had a higher RMSE percent than the branch model.
Dirichlet regression assumes that the dependent variable is a vector of proportions with unit sum and follows a Dirichlet distribution which is a multivariate generalization of the beta distribution. Unlike in beta regression, component models are fitted simultaneously, thus ensuring the unit sum of the predicted proportions. Parameter estimates and their standard errors along with fit statistics are presented in
Table 5. The biomass in stem was used as the reference group, hence there were no parameter estimates for stem biomass. One can change the reference group to obtain model coefficients for stem biomass. However, the component proportions estimated in such a manner would not necessarily have unit sum.
Similar to beta regression, the Dirichlet regression over predicted branch and stem biomass by 1.52% and 0.12% whereas the foliage biomass was under predicted by 2.36%. RMSEs produced by the Dirichlet regression (
Table 6) were practically identical to that produced by the beta regression but the Dirichlet regression should be preferred to the beta regression due to the assurance of the desired additive property.
Multinomial logistic regression is similar to the Dirichlet regression in the sense that both fit the components simultaneously and ensures that the sum of predicted proportions is equal to one. Parameter estimates and their standard errors of the multinomial logistic regression are given in
Table 7.
Multinomial logistic regression over predicted the biomass proportions for all components by no more than 0.2% (
Table 8). It produced the smallest RMSEs, compared to both beta and Dirichlet regressions. However, these RMSEs were within 0.5% of each other.
4. Conclusions
Using the data collected by destructively sampling 143 trees in 11 research plots installed in natural stands that were 11 to 26 years old, total and component biomass of brutian pine trees in Turkey were modeled. Total AGB was modeled using a nonlinear mixed effects model which accounted for the variability among plots. The predicted total AGB was then distributed into different tree components using the predicted proportions obtained from generalized linear models.
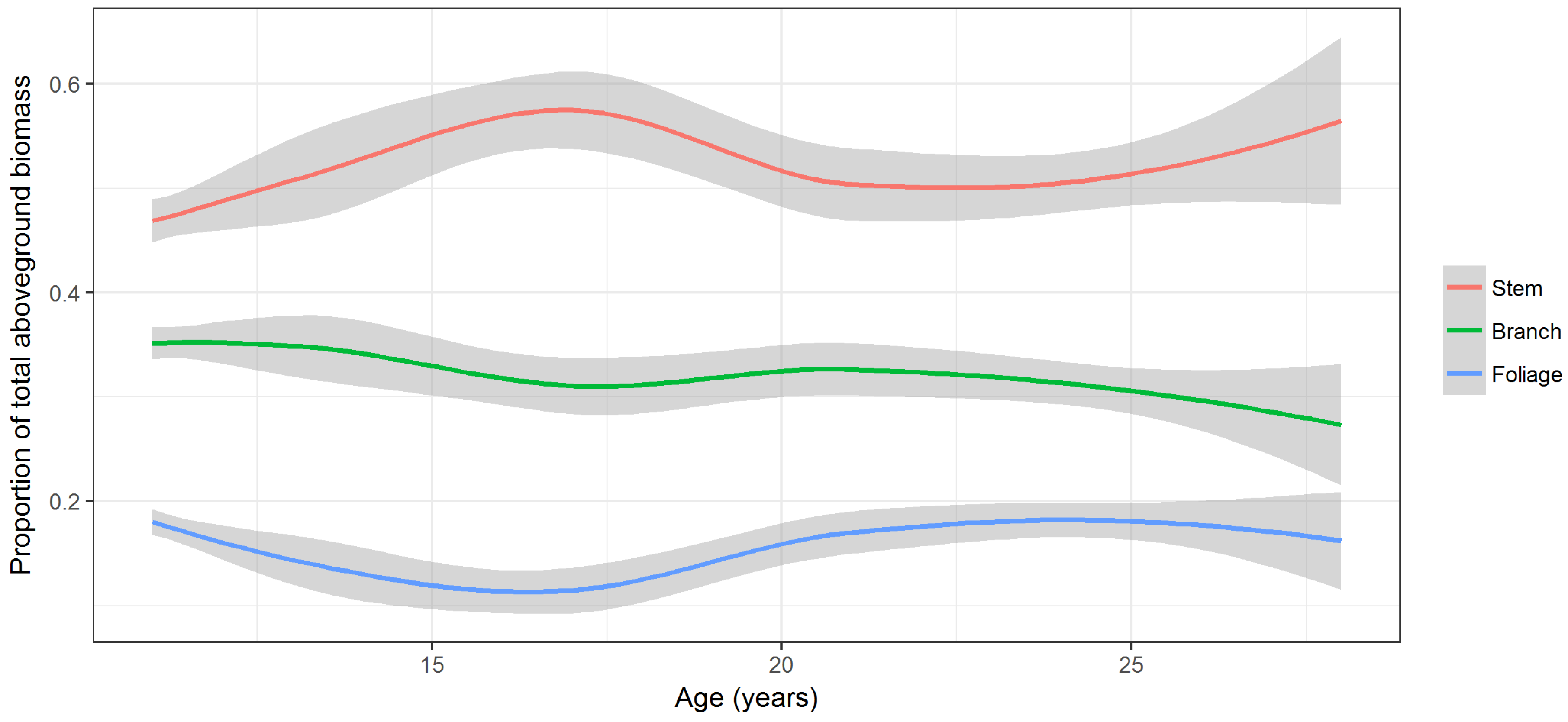
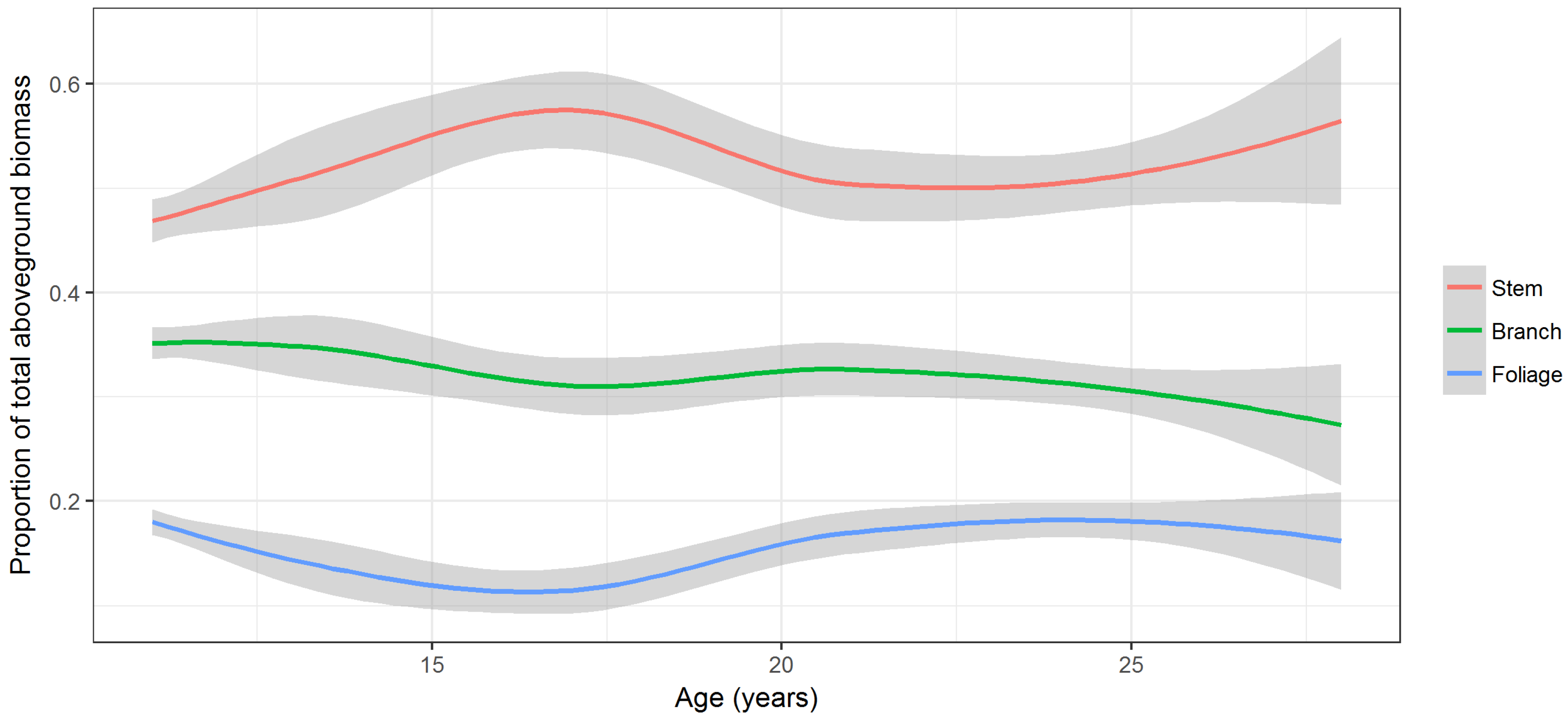
Stem, branch, and foliage biomass, on average, accounted for 50% (range 27–68%), 33% (range 18–53%), and 17% (range 4–29%) of total AGB. Biomass of different components did not follow a consistent trend with respect to tree age (
Figure 5). Proportion of stem wood biomass increased until age 16 years, then declined until age 22 years, and increased again. Proportions of branch and foliage biomass declined until age 16, then increased until age 21, and declined again thereafter. The foliage proportion had a similar trend to the branch proportion but the second decline began after around age 25 years (
Figure 5).
The beta, Dirichlet, and multinomial logistic regressions produced unbiased estimates of biomass proportions. These methods produced similar bias and root mean squared error values. The beta regression fits the component models independently. Therefore, it does not guarantee that the sum of the predicted proportions is equal to one. However, the Dirichlet and multinomial logistic regressions fit component models simultaneously and ensure the additivity of component masses. Note that the use of multinomial logistic regression requires arbitrary categorization and provides the predicted probabilities of those categories. In component biomass modeling, such predicted probabilities are considered the predicted proportions. The Dirichlet regression has the additive property, does not require such non-standard data categorization, and has similar performance as the multinomial logistic regression and may be preferred over the multinomial logistic regression. Models developed in this study can also be used in feasibility analysis of theoretical potential of establishing bioenergy plants, assessment of wildfire fuel load, and potential of brutian pine for carbon sequestration. Since the prediction accuracy of component biomass is dependent on the accuracy of the model to predict total AGB, future work on testing model forms and modeling approaches for AGB prediction with larger datasets is also critical.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}