Abstract
Species-specific and mixed-species volume and above ground biomass allometric equations were developed for 15 indigenous tree species and four tree fern species in New Zealand. A mixed-species tree equation based on breast height diameter (DBH) and tree height (H) provided acceptable estimates of stem plus branch (>10 cm in diameter over bark) volume, which was multiplied by live tree density to estimate dry matter. For dead standing spars, DBH, estimated original height, actual spar height and compatible volume/taper functions provided estimates of dead stem volume, which was multiplied by live tree density and a density modifier based on log decay class from field assessments to estimate dry matter. Live tree density was estimated using ratio estimators. Ratio estimators were based on biomass sample trees, and utilized density data from outerwood basic density surveys which were available for 35 tree species sampled throughout New Zealand. Foliage and branch (<10 cm in diameter over bark) dry matter were estimated directly from tree DBH. Tree fern above ground dry matter was estimated using allometric equations based on DBH and H. Due to insufficient data, below ground carbon for trees was estimated using the default IPCC root/shoot ratio of 25%, but for tree ferns it was estimated using measured root/shoot ratios which averaged 20%.
1. Introduction
Mankind has the potential to greatly influence atmospheric CO2 concentrations through the effects of land use change and forest management activities on the terrestrial biomass carbon pool [1]. A large number of biomass data sets and allometric equations have recently been compiled to predict above ground carbon in forest trees [2,3,4,5], motivated by the need to estimate carbon stocks for national greenhouse gas balance estimates [6]. It is important that accurate allometric equations are available to estimate carbon stocks from national plot inventory data. New Zealand has approximately 8.1 million ha of natural forest [7]. Most natural forest types in New Zealand consist of species rich mixtures of hardwood and softwood trees and tree ferns [8]. Allometric equations have been based on diameter at breast height (DBH), or DBH and height (H); however, DBH, H and density were included in allometric equations developed previously in New Zealand to estimate above ground biomass from plot data [9,10] because tree height H is not proportional to DBH [11] and wood density differs widely among species.
New Zealand’s natural forest carbon balance is estimated using a national network of inventory plots installed on an 8 km grid, with plots first measured between 2002 and 2007 [12], as part of NZ’s Land Use and Carbon Analysis System (LUCAS) operated by the Ministry for the Environment. The plot size and tree measurement protocols currently being used were based on 20 × 20 m National Vegetation Survey (NVS) plots [10,13] to ensure consistency with site-specific surveys undertaken previously in New Zealand. Some enhancements to field protocols and calculation methods were necessary to provide improved estimates of stocks in the five IPCC terrestrial carbon pools [14,15,16], and these were tested in a pilot study across a range of terrain and vegetation types in the South Island [10]. The pilot study showed that almost 60% of the forest carbon stock consisted of live biomass with an additional 10% in the dead wood pool [10], and it was recognized that more biomass and wood density data were needed to better represent the range of forest types and tree species [10].
A systematic biomass sampling programme across the whole of New Zealand’s natural forest area was not possible because tree felling was not permitted in the conservation estate. New biomass data were therefore collected from sites where recent tree falls had occurred following cyclonic storms and from plantations where felling was permitted. Biomass data collection procedures were designed to allow the development of species-specific allometric equations for estimating dry matter directly from DBH and H. In addition, indirect calculation methods that utilized supplementary density data from national breast height outerwood density surveys were included, because these allowed improved estimates of whole stem plus branch wood density by species using ratio estimators [17] at reduced cost relative to traditional biomass methods.
This paper documents the forest biomass study sites and sampling procedures that allowed the development of allometric equations for calculating tree and tree fern carbon stocks in pools 1–3 below, consistent with New Zealand’s definitions of carbon pools [16] for natural forest:
- 1. Above ground biomass (AGB) for stems ≥2.5 cm DBH;
- 2. Below ground biomass in live roots (BGB);
- 3. Dead wood (includes standing dead trees, spars, and dead roots, and coarse woody debris ≥10 cm in diameter on the forest floor);
- 4. Litter (includes dead leaves, reproductive parts, and woody debris <10 cm in diameter), and fermenting (F) and humus (H) material on the forest floor, and
- 5. Mineral soil organic carbon to 30 cm depth.
The litter and soil pools are directly assessed in LUCAS natural forest plots. Carbon stock calculation methods for pools 4 and 5 and for regenerating natural forest are described elsewhere.
2. Methods
2.1. Sites
Biomass measurements were made at six sites in the North Island and one site in the South Island of New Zealand.
2.1.1. Whirinaki Forest
Biomass sampling of podocarp forest occurred at a 40 ha trial established in 1979 at Okurapoto (latitude 38°39' S and longitude 176°42' E) in Whirinaki Forest. Altitude averaged 460 m [18,19], precipitation averaged ca. 1500 mm per year, and mean annual temperature (MAT) averaged 11 °C. The terrain was flat to gently rolling and the soil parent material was derived from ejecta from the Taupo volcanic centre. The present forest arose since the last cataclysmic Taupo eruption approximately 1800 years BP. The canopy ranged 45–60 m in height, and was dominated by rimu (Dacrydium cupressinum Lamb.) and matai (Prumnopitys taxifolia (D. Don) de Laub.), with occasional kahikatea (Dacrycarpus dacrydioides (A. Rich.) de Laub.) and totara (Podocarpus totara D. Don). The podocarps emerge above a subcanopy dominated by miro (Prumnopitys ferruginea (D. Don) de Laub.), tawa (Beilschmiedia tawa (A. Cunn.) Kirk) and other minor hardwood species up to 35 m in height. Tree species sampled for biomass between 2001 and 2007 included rimu, miro, matai, totara, kahikatea, and tawa. A continuous record of natural tree mortality from windfall events was available at this site. Only the recent windfalls had green foliage attached, while older windfalls had lost most foliage and sapwood had evidence of pinhole borer (Platypus spp.). Trees selected for biomass sampling had been lying on the forest floor for one to five years, which we considered to be representative of live standing trees in the area because little decay had occurred [17].
2.1.2. Kaimanawa Forest
This study site is situated in southern beech forest at Clements Mill Road in Kaimanawa Forest (latitude 38°57' S and longitude 176°13' E). Altitude averaged 750 m, precipitation averaged ca. 1700 mm per year, and MAT averaged 9.5 °C. The terrain was undulating, and as at Okurapoto, the soil parent material was derived from ejecta from the Taupo volcanic centre, the last major episode being the Taupo eruption (ca. 1800 years BP). The forest canopy ranged 25–35 m in height, and was dominated by red beech (Nothofagus fusca (Hook. f.) Oerst.) and silver beech (Nothofagus menziesii (Hook. f.) Oerst.). Gaps created by previous windthrow events were occupied predominantly by even-aged pole beech stands. Species sampled between 2005 and 2008 included red beech and silver beech. Very recent windfalls had green foliage attached, while older windfalls had lost most foliage, and sapwood had evidence of pinhole borer (Platypus spp.). As at Whirinaki the trees selected for biomass sampling had been lying on the forest floor for one to five years, by which time little decay was evident [17].
2.1.3. Mt Maungatautari
Trees were felled during construction of a predator-proof fence at Mount Maungatautari (797 m altitude, latitude 38°01' S and longitude 175°34' E), in the Waikato District. Altitude of biomass tree locations ranged 300–400 m. Annual rainfall at Maungatautari averaged 1374 mm and annual daily temperature at Cambridge, 18 km to the northwest averaged 13.3 °C. The forest canopy ranged 20–30 m in height, and was dominated by tawa, mangeao (Litsea calicaris (A. Cunn.) Benth. and Hook.f.), rewarewa (Knightia excelsa (Mull. Hal.) R. Br.), pukatea (Laurelia novae zealandiae A.Cunn.), and kamahi (Weinmannia racemosa J.R.Forst), with a subcanopy of hinau (Elaeocarpus dentatus (J.R.Forst. and G.Forst.)Vahl), pigeonwood (Hedycarya arborea J.R.Forst. and G.Forst), mahoe (Melicytus ramiflorus J.R.Forst. and G.Forst), and tree ferns. Species sampled between 2005 and 2006 included kahikatea, tawa, mangeao, rewarewa, pukatea, pigeonwood, and one species of tree fern. Only recently felled trees were sampled.
2.1.4. Taranaki
Kauri (Agathis australis (D. Don) Lindl.) trees were being felled as part of a stand thinning operation at the Fred Cowling reserve near New Plymouth (latitude 39°05' S and longitude 174°03' E). This planted stand was 69 years old when sampled, and was still close to its original stocking of around 1300 trees per hectare. Annual rainfall and temperature averaged 1614 mm and 13.7 °C, respectively. The New Plymouth District Council manages the reserve and arranged for the careful felling of twenty trees for biomass measurement during November 2004.
2.1.5. Hunua Forest
A naturally regenerated kauri stand at Mangatangi in the Hunua ranges (latitude 37°07' S and longitude 175°13' E) was being thinned in February 1980 and biomass trees were selected as part of the thinning operation. Mean annual precipitation and temperature at Hunua were 1411 mm and 14.1 °C, respectively. Tree height at three plots installed in the vicinity of the kauri biomass study averaged 10.5 m and contained 1440 stems/ha with a basal area of 29.1 m2/ha. Ring counts indicated a stand age of approximately 130 years. Over 90% of the stems and 93% of the basal area comprised kauri, the remaining trees being tanekaha (Phyllocladus trichomanoides D. Don) [20].
2.1.6. Maimai
Stem and crown biomass of canopy and understorey species were measured in mature beech/podocarp forest at Tawhai State Forest, Maimai (latitude 42°05' S and longitude 171°48' E) [21]. Precipitation was evenly distributed and averaged 2600 mm annually. Temperature at Reefton 2 km to the east averaged 10.9 °C. The terrain was steeply dissected with shallow strongly weathered acidic-pedal allophanic brown soil of low fertility formed over compacted gravels. The forest canopy ranged from 20 to 36 m in height and was rather open with hard beech (Nothofagus truncata (Colenso) Cockayne) as the dominant species in association with scattered miro and rimu. At gully-heads and along stream margins hard beech was displaced by red beech and silver beech. The subcanopy ranged from 7 to 19 m in height and the most common species was kamahi. Tree species sampled for biomass from experimental catchments in 1975 included red beech, hard beech, miro, kamahi and two tree fern species [21].
2.1.7. Whakarewarewa Forest
Above and below ground biomass of tree ferns was measured at Whakarewarewa Forest, Rotorua (latitude 38°11' S and longitude 176°18' E). Altitude ranged 465–505 m, annual rainfall averaged 1500 mm, and temperature averaged 12.7 °C. The terrain was undulating and the soil type was Whakarewarewa hill soil [22] corresponding to an Immature Orthic Pumice Soil in the New Zealand Soil Classification system [23]. The tree canopy was comprised of a mixture of planted softwood and hardwood trees with an understorey of naturally regenerated shrub hardwoods and tree ferns. The four most abundant tree fern species in New Zealand were selected for biomass determination during March and April 2010, including silver fern (Cyathea dealbata (G.Forst.) Sw.), mamaku (C. medullaris (G.Forst.) Sw.), soft tree fern (C. smithii Hook.f.), and rough tree fern (Dicksonia squarrosa (G.Forst.) Sw.).
2.2. Biomass Procedures
2.2.1. Trees
Tree biomass measurement protocols largely follow those described in Beets et al. [17]. For large trees total height (H) and diameter over bark were measured at 0.15 m, 0.7 m, breast height (DBH, 1.4 m above ground measured from the uphill side of the tree), and thereafter at 1–3 m intervals along the stem to the top of the tree, including all forks and large branches with a small end diameter (SED) ≥10 cm over bark. Volume over bark of stem and branches ≥10 cm was calculated from the sectional measurements using the formula for truncated cones. Disk samples (ca. 40 mm thick) were cut at 1–6 m intervals along the stem and large branches (10 and 20 disks per tree), disk thickness was measured at 4 to 8 points around the perimeter of the disk using callipers, and volume over bark calculated from the over bark diameter and mean thickness. The mean density of the stem and large branches was calculated per tree as the area weighted mean density of the sample disk, and was therefore compatible with the volume measurement because both include the effects of any hollowing associated with decay and stem fluting. The dry mass of the stem and large branches ≥10 cm was calculated by multiplying volume by the corresponding mean density. Branches <10 cm in diameter and attached foliage of felled or recently fallen trees were weighed fresh (total and sample weights) in the field and samples were oven-dried and weighed. The volume of small stems was not measured; however, the total and sample fresh weight of stems and crowns were measured, and samples oven-dried and weighed. The total dry mass of branches <10 cm in diameter and foliage dry mass per tree were calculated by applying the respective fresh weight sampling fractions to the sample data. Samples were oven-dried at 70 °C. Carbon was assumed to comprise 50% of the oven-dry mass [24].
For biomass trees, breast height basic density or specific gravity (oven-dry mass at 105 °C/green saturated volume from water displacement) was measured at 0–5 and 5–15 cm depths under bark on two opposing parallel sided radial strips cut from breast height disks, to allow the development of ratio estimators (described in Section 2.3).
2.2.2. Tree Ferns
Height of the caudex was measured from ground level to the top of the solid woody part of the caudex, caudex diameter was measured at the base (0.15 m), breast height (1.4 m) and at 1m intervals along the caudex, and volume calculated using the using the formula for truncated cones. Live and dead fronds were removed and weighed. The caudex was weighed fresh and 50 mm thick disk (except the basal disk which was 25 mm thick) samples were cut at 1m intervals along the caudex and weighed. The oven dry weight of caudices, live fronds, and dead fronds were calculated by applying the respective fresh weight sampling fractions to the sample data. Samples were oven-dried at 70 °C.
Tree fern root systems were extracted by manual digging, cut into manageable sections, and the entire root system processed in the laboratory. Cutting roots into sections revealed substantial amounts of mineral soil trapped within the fibrous root matrix. Root sections were therefore washed over a 3 mm screen with a moderate pressure water hose to remove mineral soil visible on the surface, oven-dried at 70 °C and weighed. Mineral soil imbedded inside the fibrous root matrix was removed after the sections were pulverized in a chipper. The pulverized material was thoroughly mixed and a sample (approximately 10% by weight of the entire root system) immediately weighed to determine the sampling fraction by weight. Mineral soil in the sample was separated from the root material using a combination of flotation and manual sorting. Carbon was assumed to comprise 50% of the root oven dry mass.
2.3. Supplementary Wood Density Data
Breast height outerwood basic density measurements acquired throughout New Zealand were compiled in a Wood Density database, together with information on species, region, tree age, and number of trees sampled per survey. When calculating carbon in stems and branches from volume and density, the national carbon stock estimate can be improved by combining the wood density survey data with data from the biomass sample trees. The ratio estimator [25] that achieved this was:
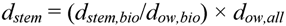
where dstem is the survey adjusted ratio estimator of mean whole stem and branch ≥10 cm density; dstem,bio is mean whole stem and branch ≥10 cm density of the biomass trees; dow,bio is the mean breast height outerwood basic density of the biomass trees; and dow,all is the mean breast height outerwood basic density of the national survey sample. Ratios (dstem,bio/dow,bio) were developed by species and across species for both the outer 0–5 cm and 5–15 cm core depths.
The wood density database contained various measures of wood density, including merchantable log basic density, breast height outerwood (0–5 cm depth or in some cases the outer 5 rings) basic density, and breast height (5–15 cm depth) basic density. To make use of all the data the various density measures were converted as follows:
- 1. If breast height basic density at 5–15 cm depth was measured, the ratio estimator was applied (following Beets et al. [17]).
- 2. If merchantable log basic density was measured, the ratio estimator was multiplied by 1.03 (1.03 was derived from the wood density database and converts breast height basic density at 0–5 cm depth to breast height basic density at 5–15 cm depth. It should be noted that merchantable log basic density and breast height basic density at 0–5 cm depth have a 1:1 conversion, based on the database).
- 3. If breast height outerwood (0–5 cm) basic density was measured, the ratio estimator was multiplied by 1.03.
2.4. Allometric Equations for Live Trees and Tree Ferns
The components available for analysis included volume and carbon in the stem plus large branch ≥10 cm diameter over bark, carbon in branch <10 cm diameter, and carbon in foliage. Allometric equations were developed to estimate carbon in stem plus branch ≥10 cm diameter directly from DBH and H. In addition, indirect estimates of carbon in stem plus branch ≥10 cm diameter were obtained by multiplying volume (from the allometric equation based on DBH and H) by the corresponding density. Carbon in branches <10 cm and foliage were estimated directly from DBH and the components were summed to give above ground live tree carbon. Tree components were estimated separately and summed because all components were not measured on every tree. The tree and tree fern biomass data are given in Appendix 1.
The following allometric regression equation was used:
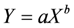
where a and b are parameters; with asp replacing a when testing for species effects; X for trees is DBH (cm) or DBH2 × H (cm2 m); Y is stem plus large branch over bark volumeob (m3 tree−1) or component carbon weights (kg tree−1).
Equation 2 was fitted using a generalized linear regression model with gamma error distribution and log link function [26], using SAS Version 9.1 (2004) procedure PROC GENMOD. This procedure gives a less biased fit than the alternative method of fitting a linear regression equation to the logs of the x and y variables. Tree species were analysed separately from tree fern species. To test for species differences, models using species specific “a” parameter (asp) and a common “b” parameter were fitted (which is equivalent to fitting separate intercepts and a common slope on the log-log scale). Species were compared using an approximate χ2 test. This analysis helps to inform whether tree carbon should be estimated from allometric volume equations coupled with density or estimated directly from DBH and H.
2.5. Allometric Equations for Dead Stems and Spars
The volume of standing dead stems, which often occur as truncated spars, was calculated using the following compatible volume (Equation 3) and taper (Equation 4) functions [27,28,29] which were fitted to the subset of biomass trees that had stem sectional measurements:

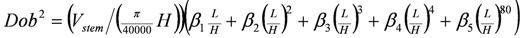
where b1, b2 and β1–β5 are parameters; Vstem is intact stem wood plus bark volume (excluding volume in large branches); Dob is stem diameter over bark at height L; and DBH and H are as defined previously. The parameters of the volume function (b1 and b2) were fitted using log transformations with b1 adjusted for logarithmic bias. The taper function (Equation 4) is a 4th order polynomial with an additional higher order term to account for butt swell [29]. The parameters of the taper function (β1–β5) were fitted subject to a constraint which ensured that when integrated over total tree height the function yielded the same volume as Equation 3 (see [28,29] for details). An equation for estimating the volume of the spar was then obtained by integrating Equation 4 from the base of the stem to a height representing the top of an intact dead tree or a truncated spar.
3. Results
3.1. Allometric Equations
3.1.1. Characteristics of Biomass Trees and Tree Ferns
A list of tree species, the number of biomass trees with crown and stem data used to develop allometric equations, and their mean dimensions are given by site in Table 1. Trees ranged between 0.7 and 142 cm in DBH, and between 2.4 and 59 m in total height (Appendix 1).

Table 1.
Tree and tree fern species sampled at biomass study sites throughout New Zealand, sample size (n for stem, crown, root), and mean diameter at breast height (DBH) and height (H).
| Site | Species | Sample size | ||||
|---|---|---|---|---|---|---|
| Means | ||||||
| nstem | ncrown | nroot | DBH (cm) | H (m) | ||
| Whirinaki | B. tawa | 10 | 0 | 36.9 | 26.5 | |
| D. cupressinum | 18 | 0 | 95.8 | 50.1 | ||
| D. dacrydioides | 2 | 0 | 140.0 | 52.0 | ||
| P. taxifolia | 12 | 0 | 75.3 | 38.8 | ||
| P. totara | 1 | 0 | 107.6 | 46.7 | ||
| P. ferruginea | 7 | 0 | 49.2 | 34.3 | ||
| Kaimanawa | N. fusca | 16 | 0 | 39.5 | 24.1 | |
| N. menziesii | 12 | 1 | 59.3 | 27.2 | ||
| Hunua | A. australis | 6 | 6 | 19.4 | 14.6 | |
| Taranaki | A. australis | 20 | 10 | 33.2 | 23.1 | |
| Maimai | N. fusca | 2 | 2 | 89.0 | 32.2 | |
| N. truncata | 11 | 11 | 41.8 | 20.9 | ||
| P. ferruginea | 1 | 1 | 32.0 | 23.9 | ||
| W. racemosa | 36 | 36 | 8.6 | 7.8 | ||
| Maungatautari/Manawahe | L. calicaris | 7 | 1 | 40.1 | 20.7 | |
| H. arborea | 2 | 2 | 30.0 | 14.3 | ||
| L. n. zealandiae | 6 | 0 | 40.0 | 18.0 | ||
| K. excelsa | 4 | 0 | 35.5 | 19.5 | ||
| B. tawa | 15 | 0 | 36.5 | 19.3 | ||
| D. dacrydioides | 1 | 0 | 20.4 | 8.2 | ||
| W. racemosa | 1 | 0 | 33.0 | - | ||
| Maimai | C. smithii | 2 | 2 | 19.4 | 7.7 | |
| Maungatautari | C. dealbata | 1 | 1 | 28.5 | 8.2 | |
| Whakarewarewa | C. dealbata | 20 | 20 | 4 | 27.3 | 2.7 |
| C. medullaris | 20 | 20 | 5 | 21.6 | 4.6 | |
| C. smithii | 20 | 20 | 5 | 14.3 | 2.9 | |
| D. squarrosa | 20 | 20 | 4 | 15.9 | 3.2 | |
3.1.2. Stem Volume and Carbon in Live Trees
Mixed-species allometric equations are given in Table 2. These provide generalised estimates of volume over bark and biomass carbon for stems and branches ≥10 cm, and carbon in small branches <10 cm and foliage across species and sites. The fit of each equation is indicated by the percentage of the deviance explained, which is analogous to the R2 coefficient.
Table 2.
Mixed-species allometric equations for estimating stem and large branch volume and component carbon for natural forest trees, sample size (n), and deviance explained. The equation form is Y = aXb.
| Y | Stem & Branch (≥10 cm) Volumeob (m3/tree) | Stem & Branch (≥10 cm) Carbon (kg/tree) | Branch (<10 cm) Carbon (kg/tree) | Foliage Carbon (kg/tree) |
|---|---|---|---|---|
| X | DBH2·H (cm2·m) | DBH2·H (cm2·m) | DBH (cm) | DBH (cm) |
| a | 4.83 × 10−5 | 1.62 × 10−2 | 1.75 × 10−2 | 1.71 × 10−2 |
| b | 0.978 | 0.943 | 2.20 | 1.75 |
| Deviance explained % | 98.9 | 98.8 | 96.7 | 85.2 |
| Sample size n | 141 | 127 | 70 | 70 |
Tests of species differences in these allometric equations were highly significant (i.e., when p < 0.05) when estimating carbon in stem and branch ≥10 cm but barely significant (minor) for volume, due to large species differences in density (Table 3). The “a” parameter for volume averaged 5.39 × 10−5 for softwoods and 5.19 × 10−5 for hardwoods, and will reflect the inclusion of large branches with stems. Species effects were not significant for estimating carbon in branches <10 cm; however, species effects were significant for foliage, due to the inclusion of naturally regenerated and planted pole kauri trees within the same regression equation as mature trees.
Table 3.
Allometric equations for estimating stem and large branch volume and component carbon for natural forest trees with species specific parameters. Values in a column followed by the same letter do not differ significantly (least significant difference test with p = 0.05).
| Y | Stem & Brch (≥10 cm) Volumeob (m3/tree) | Stem & Brch (≥10 cm) Carbon (kg/tree) | Branch (<10 cm) Carbon (kg/tree) | Foliage Carbon (kg/tree) | |||
|---|---|---|---|---|---|---|---|
| X | DBH2·H (cm2·m) | DBH2·H (cm2·m) | DBH (cm) | DBH (cm) | |||
| b | 0.968 | 0.936 | 0.936 | 1.595 | |||
| asp parameters for: | |||||||
| A. australis | 5.39 | bc | 0.0157 | de | 0.0156 | 0.0541 | a |
| B. tawa | 5.46 | ab | 0.0190 | bc | |||
| D. cupressinum | 5.46 | b | 0.0182 | bcd | |||
| D. dacrydioides | 4.90 | bc | 0.0105 | f | |||
| H. arborea | 4.65 | bc | 0.0141 | def | 0.0274 | 0.0045 | d |
| K. excelsa | 4.67 | bc | 0.0170 | bcde | |||
| L. calicaris | 4.33 | ac | 0.0135 | ef | 0.0147 | 0.0071 | cd |
| L. n. zealandiae | 4.84 | bc | 0.0116 | f | |||
| N. fusca | 5.47 | b | 0.0189 | abcd | 0.0147 | 0.0090 | cd |
| N. menziesii | 6.18 | a | 0.0256 | a | 0.0220 | 0.0474 | a |
| N. truncata | 5.14 | bc | 0.0196 | ab | 0.0219 | 0.0167 | bc |
| P. ferruginea | 5.53 | ab | 0.0208 | ab | 0.0138 | 0.0132 | bc |
| P. taxifolia | 5.58 | ab | 0.0222 | ab | |||
| P. totara | 5.45 | abc | 0.0129 | cdef | |||
| W. racemosa | 5.15 | bc | 0.0168 | cde | 0.0172 | 0.0186 | b |
| asp parameters ×10−5 | |||||||
| Deviance explained % | 99.1 | 99.3 | 99.3 | 95.0 | |||
| Sample size n | 141 | 127 | 70 | 70 | |||
| Test for species differences | F14,125 = 1.94, p = 0.028 | F14,111 = 5.91, p ≤ 0.0001 | F14,111 = 1.35, p = 0.22 | F7,61 = 17.13, p ≤ 0.0001 | |||
3.1.3. Stem Volume in Standing Dead Trees and Spars
The number of biomass trees with sectional measurements for developing compatible volume and taper functions and their mean dimensions are given in Table 4. Trees ranged between 5.3 and 142 cm in DBH, and between 7.7 and 59 m in total height.
Table 4.
Sample size (n) and dimensions of biomass trees with stem sectional measurements suitable for developing volume/taper functions.
| n | Minimum | Mean | Maximum | Standard Deviation | |
|---|---|---|---|---|---|
| DBH | 115 | 5.3 | 52.9 | 142.0 | 34.7 |
| H | 115 | 7.7 | 27.9 | 59.2 | 12.7 |
| Stem volumeob | 115 | 0.011 | 5.46 | 32.1 | 7.91 |
| Form factor | 115 | 0.254 | 0.485 | 0.691 | 0.078 |
Based on these trees, the over-bark volume Vstem of an intact stem of height H was estimated as:
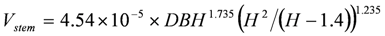
And the following equation for estimating the volume of a dead stem or spar with measured height Hdead was obtained by integrating the fitted Equation 4 along the stem length up to the measured height:
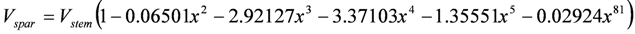
where x = (H − Hdead)/H.
To estimate the volume of standing dead stems (or spars) using Equations 5 and 6 the height H of a live stem with the same DBH as the dead stem must be estimated using the live stem height/diameter function for the species and plot where the dead stem occurs. If H is less than Hdead, x is set to zero.
3.1.4. Caudex Volume and Carbon in Tree Ferns
Allometric equations for estimating the volume of the caudex and above ground carbon of tree ferns at Whakarewarewa are given in Table 5. These provide generalised estimates of volume and biomass carbon of the caudex, caudex plus fronds, and roots across species. Three tree ferns sampled at other sites fell within the range of data scatter found at Whakarewarewa, which provides weak evidence that site differences were small. Tree fern caudices were not expected to break during storms so the volume of dead tree fern caudices was calculated using the allometric equation for live tree ferns. The parameter, b (for volume using Equation 2) was 0.978 for trees and 1.22 tree ferns. The diameter of tree fern caudices tended to increase both below and above the diameter at breast height, which explains why the value exceeds 1.0.
Table 5.
Mixed-species allometric equations for estimating caudex volume and component carbon for tree ferns, sample size (n), and variance explained. The equation form is Y = aXb.
| Y | Caudex Volume (m3/tree) | Caudex & Frond Carbon (kg/tree) | Caudex Carbon (kg/tree) | Root Carbon (kg/tree) |
|---|---|---|---|---|
| X | DBH2·H (cm2·m) | DBH2·H (cm2·m) | DBH2·H (cm2·m) | DBH2·H (cm2·m) |
| a | 1.34 × 10−5 | 2.70 × 10−3 | 1.10 × 10−3 | 6.99 × 10−4 |
| b | 1.22 | 1.19 | 1.25 | 1.14 |
| Deviance explained % | 93.1 | 91.6 | 91.9 | 81.0 |
| n | 80 | 80 | 80 | 18 |
The fibrous nature of tree fern caudices precludes density determination from breast height cores, and above ground biomass carbon (caudex and fronds) of tree ferns therefore needs to be estimated using the generic allometric equation in Table 5. Tests of species differences in these allometric equations showed small although statistically significant effects which resulted from species differences in fern caudex density and volume parameters (Table 6). There was also weak evidence for species differences in root/shoot ratios, which averaged 14%, 21%, 19%, and 32% for C. medularis, C. dealbata, C. smithii, and D. squarrosa, respectively. Analysis of variance indicated that the root/shoot ratio of the first three of these species did not differ significantly from each other but the root/shoot ratio of D. squarrosa was significantly higher than in other species. The overall mean root/shoot ratio was 20%.
Table 6.
Allometric equations for estimating caudex volume and component carbon for tree ferns with species specific parameters. Values in a column followed by the same letter do not differ significantly (least significant difference test with p = 0.05).
| Y | Caudex Volume (m3/tree) | Caudex & Frond Carbon (kg/tree) | Caudex Carbon (kg/tree) | Root Carbon (kg/tree) |
|---|---|---|---|---|
| X | DBH2·H (cm2·m) | DBH2·H (cm2·m) | DBH2·H (cm2·m) | DBH2·H (cm2·m) |
| b | 1.26 | 1.06 | 1.23 | 1.20 |
| asp parameters for: | ||||
| C. medullaris | 1.06 × 10−5 a | 8.45 × 10−3 a | 1.32 × 10−3 ab | 4.05 × 10−4 b |
| C. dealbata | 8.15 × 10−6 b | 7.59 × 10−3 a | 1.36 × 10−3 ab | 4.37 × 10−4 b |
| C. smithii | 1.01 × 10−5 a | 5.31 × 10−3 c | 1.13 × 10−3 b | 3.38 × 10−4 b |
| D. squarrosa | 1.03 × 10−5 a | 6.33 × 10−3 b | 1.48 × 10−3 a | 7.90 × 10−4 a |
| Deviance explained % | 94.1 | 93.6 | 92.7 | 90.4 |
| Sample size n | 80 | 80 | 80 | 18 |
| Test for species differences | F3,75 = 4.12, p = 0.0093 | F3,75 = 7.87, p = 0.0001 | F3,75 = 2.80, p = 0.046 | F3,13 = 4.29, p = 0.026 |
3.1.5. Carbon in Below Ground Biomass
A small number of root biomass studies have been made in New Zealand, and more studies of tree root biomass should be undertaken. While the few studies in natural forest were comprehensive, sampling variation was large and encompassed IPCC defaults for root/shoot ratios in temperate broadleaf forest [21]. Because of the lack of New Zealand data, the below ground biomass carbon pool was calculated using a root/shoot ratio of 25% for trees, based on IPCC guidelines for mature temperate broadleaf forests [16], and 20% for tree ferns, based on biomass data in this paper.
3.2. Ratio Estimators
Ratios of whole stem and branch ≥10 cm density to breast height outerwood basic density and the number of biomass sample trees on which the ratios were based are given in Table 7. Species differences were more pronounced for ratios based on the outer 0–5 cm compared with 5–15 cm breast height core depths. For the 5–15 cm depth, the species with ratios differing significantly from the mean were D. dacrydioides, H. arborea, and L. calicaris, but these species were each represented by only two trees in the biomass data set, and had significantly lower ratios than the mean. The use of a common ratio (0.905) based on 68 trees with breast height cores at 5–15 cm depth was considered reasonable, given that the sample size for most species with biomass data was usually too small to justify the use of individual species ratios.
Table 7.
Number of biomass trees with the requisite data (N) and the calculated mean ratios of whole stem and branch density to breast height outerwood basic density by species and over all species. Values in a column followed by the same letter do not differ significantly (least significant difference test with p = 0.05).
| Species | N | Ratio of whole stem to 0–5 cm BH basic density | Ratio of whole stem to 5–15 cm BH basic density | ||||
|---|---|---|---|---|---|---|---|
| Mean | s.e. | Mean | s.e. | ||||
| A. australis | 20 | 0.969 | cde | 0.017 | 0.938 | a | 0.015 |
| B. tawa | 5 | 0.891 | def | 0.035 | 0.898 | ac | 0.029 |
| D. cupressinum | 10 | 1.083 | ab | 0.024 | 0.933 | ab | 0.021 |
| D. dacrydioides | 2 | 0.909 | cdef | 0.055 | 0.807 | ac | 0.047 |
| H. arborea | 2 | 0.831 | ef | 0.055 | 0.785 | ac | 0.047 |
| K. excelsa | 2 | 0.976 | bcde | 0.055 | 0.993 | a | 0.047 |
| L. calicaris | 7 | 0.846 | f | 0.029 | 0.844 | c | 0.025 |
| L. n. zealandiae | 5 | 0.940 | cde | 0.035 | 0.913 | a | 0.029 |
| N. fusca | 2 | 0.869 | def | 0.055 | 0.879 | ac | 0.047 |
| N. menziesii | 2 | 1.010 | bcd | 0.055 | 0.886 | abc | 0.047 |
| P. ferruginea | 6 | 1.018 | bc | 0.032 | 0.904 | ab | 0.027 |
| P. taxifolia | 4 | 0.999 | bcd | 0.039 | 0.891 | ab | 0.033 |
| P. totara | 1 | 1.234 | a | 0.077 | 0.806 | b | 0.066 |
| All species | 68 | 0.968 | 0.013 | 0.905 | 0.009 | ||
| Test for species differences | F12,55 = 5.92, p ≤ 0.0001 | F12,55 = 2.52, p = 0.010 | |||||
National estimates of whole stem and branch ≥10 cm density were obtained by applying the ratio estimator for all species to the breast height outerwood basic density means of the 35 tree species in national wood density surveys (Appendix 2). Some commonly occurring species were represented by more than 1000 sample trees nationally. Statistically significant regional differences in density within species were rarely found, and regional differences from national means were generally small ( 2) for most species. For example, K. excelsa (3%), N. menziesii (3%), N. solandri var. cliffortioides (4%), for regions with 10 or more trees with density data per region. Only two species included information on tree age, which was not significant in either case.
4. Discussion
Carbon stocks/ha can be estimated by applying allometric equations to independent variables such as the DBH and height measured in inventory plots. The new allometric equations for live and dead trees and tree ferns reported in this paper are intended to replace methods used previously by Beets [21], Hall et al. [13], and Coomes et al. [10], to estimate carbon in New Zealand’s natural forest. The new biomass equations were based on approximately 140 trees and 80 tree ferns, whereas equations used previously were based on 50 trees and two tree ferns from one site in the South Island. The new equations included 12 large biomass sample trees >1 m DBH—the old equations were based on one large tree. Predictions from the old and new equations were compared using two hundred tree records selected at random from the ten most common species in the LUCAS natural forest database. Above ground carbon predictions were on average 11% higher using the new equations. Both predictions were based on a mixed-species volume equation (requiring DBH, height) and species-specific density. The comparison showed that, after almost trebling the sample size, predictions from the new volume equation (Equation 7) decreased by 2% for trees <10 cm DBH and increased by 21% for trees ≥1 m DBH compared with predictions from the old volume equation in Coomes et al. [10]; however, the new equation includes the volume of branches ≥10 cm in diameter. Predictions of carbon in small branches and foliage necessarily decreased using the new equations, because they apply to branches <10 cm in diameter. The density adjustment factor for hollow trunks used in the old equation was not robust, because it gave negative predictions for stem carbon in very large trees (over 5.3 m DBH). Substantially increasing the sample size would undoubtedly allow further improvement of allometric functions at considerable cost, although it seems unlikely that the predictions will change appreciably.
Our tree biomass procedures combined the stem with the large branches because they were often difficult to distinguish, particularly in hardwood species. In addition, because sampling of windfall trees was necessary, biomass measurement focused on the stem and large branches ≥10 cm diameter owing to the loss of foliage and small branches. Biomass of small branches, twigs, and foliage of recent windblown trees with green foliage present, and of felled trees, were measured. After allowing for the effects of stem diameter, biomass of branches <10 cm in diameter over bark did not differ significantly among species; however for foliage the parameter for kauri exceeded that in other species. This may reflect the comparatively young age of the kauri stands. The foliage parameter for silver beech also appeared to be higher than in other species, although uncertainty was large because only three silver beech crowns were measured. Nevertheless, the overall impact of using mixed-species equations to estimate branches <10 cm in diameter and foliage carbon can be expected to be small, because 85% of the above ground biomass carbon was contained in stems and large limbs, while only 13% was in branches <10 cm diameter, and 1.7% in foliage.
Species differences in stem form were statistically significant but small when examined by species group–for example, the model error for volume of stems plus branches ≥10 cm diameter averaged +0.3% for podocarps, −0.3% for Kauri, −2.9% for hardwoods and −0.5% overall, which justifies the use of a mixed-species allometric model to estimate volume. Model error for trees ≥5.0 to ≤10 cm diameter averaged −5.7%, which indicates that volume of small trees was slightly underestimated. The model error was +0.4% for planted kauri and −1.2% for kauri in naturally regenerating natural forest, which indicates that plantation grown kauri is similar in stem form to naturally regenerated kauri.
The species-specific allometric equations for estimating above ground dry matter directly from DBH and H demonstrated the importance of including density as an independent variable when using mixed-species allometric equations. With only a limited amount of tree biomass data, we think it is important to utilize the supplementary breast height outerwood density survey data when estimating carbon stocks in New Zealand’s natural forest. It was assumed that carbon comprised 50% of the dry matter. This assumption is expected to be reasonable when applied over a wide range of tree species and components; however, this assumption should ideally be tested.
A mixed-species function involving DBH, H, and density was developed by Chave et al. [9] who considered it important to test their tropical equations using temperate forest tree biomass data. We did this by comparing above ground biomass carbon predictions from their best wet tropical forest regression model (with tree DBH, total height, and specific gravity) with our biomass data. For calculation purposes, we used the species mean breast height outerwood basic density (at 5–15 cm depth) from survey cores as the measure of specific gravity. The ratio estimator provided species-specific estimates of whole stem and large branch density for use with our natural forest above ground biomass carbon allometric Equation 8. Note that the “b” parameter in the wet tropical forest equation applies to both the volume index (DBH2 × H) and wood specific gravity, whereas the natural forest “b” parameter applies only to the volume index. Model error for our natural forest allometric equation and the wet tropical forest equation averaged +3.0% versus +2.0%, respectively for trees ≥2.5 cm DBH with complete above ground biomass data (Figure 1). Errors tended to be larger using specific subsets of our biomass dataset, for example +8.7% versus +6.8% for 13 beech trees, −6.6% versus −7.1% for 29 trees of the subcanopy species Weinmannia racemosa, +3.8% versus +1.9% for 45 hardwood trees, +2.4% versus +6.1% for 16 pole kauri. Model error for small trees ≥2.5 to ≤10 cm averaged −11.4% using the natural forest allometric equation and −8.3% using the wet tropical forest allometric equation; however, the impact on forest carbon stock prediction will likely be small. It is clear that both equations provided reasonable predictions of above ground biomass carbon across a range of groups of species and tree size classes.
Various other researchers have compared allometric equations they developed for estimating above-ground biomass in tropical forest [30,31] with the pan-tropical equations developed by Chave et al. [9], and concluded that general allometric equations were biased when applied at their sites. This was surprising to us, considering the results of the comparisons we made. The reported biases may be due to large differences in stem form factor among species, although biases were up to 50%. Model comparisons they made presumably involved equations based on DBH alone, DBH and density, or DBH, H, and density. If the latter type gave large differences between studies it would be useful to know if merchantable height was used instead of total height. Ignoring height assumes an invariant DBH/height relationship, which is unlikely in mixed-species forests. Furthermore, tree species composition can change both spatially and temporally, following for example the selective harvesting of desirable species. Comparisons we made, using various groupings (hardwood, podocarps, kauri, small trees), found few notable model prediction errors using either our natural forest allometric equation or the best wet tropical equation of Chave et al. [9].
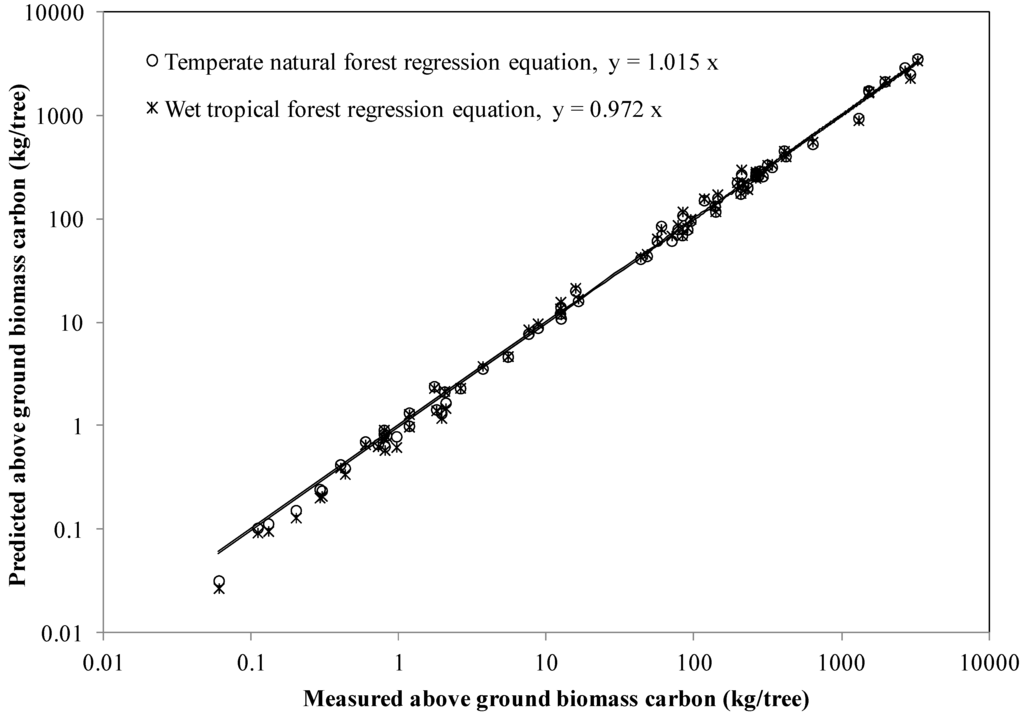
Figure 1.
Measured versus predicted above ground carbon for temperate hardwood/softwood trees (DBH range 2.8–142 cm) in New Zealand obtained using the natural forest equation for New Zealand and wet pan-tropical forest equation of Chave et al. [9]. Independent variables were breast height diameter, total height, and wood density.
Figure 1.
Measured versus predicted above ground carbon for temperate hardwood/softwood trees (DBH range 2.8–142 cm) in New Zealand obtained using the natural forest equation for New Zealand and wet pan-tropical forest equation of Chave et al. [9]. Independent variables were breast height diameter, total height, and wood density.
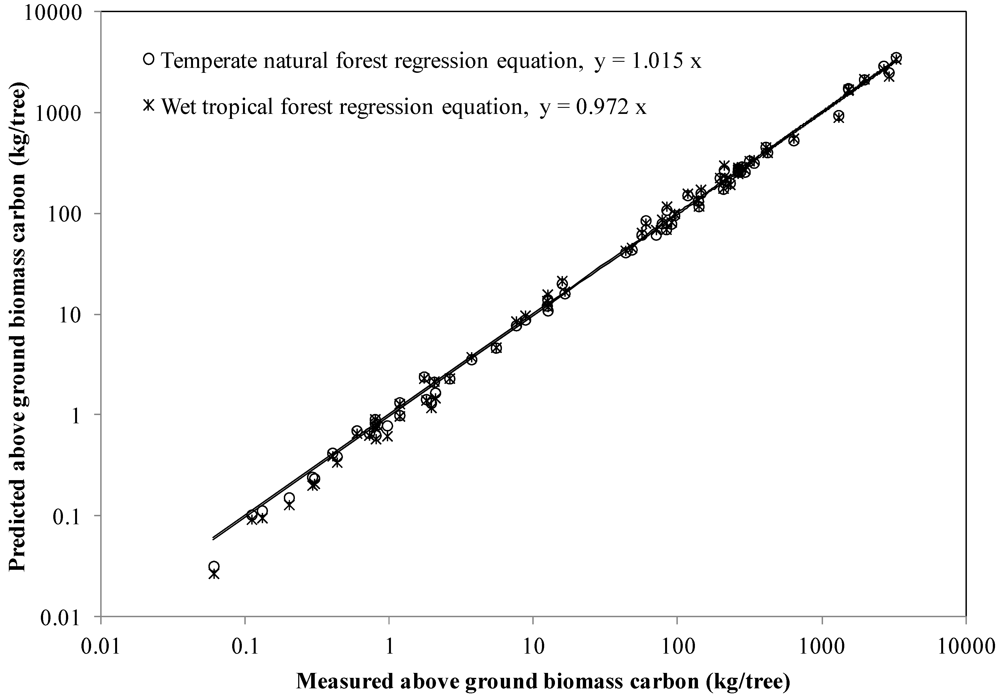
Our analysis showed that regional variation in mean breast height outerwood basic density was not statistically significant in all but a few species, so national estimates of whole stem and branch ≥10 cm density over bark were obtained by species using a ratio estimator, following methods in Beets et al. [17]. Unlike outerwood basic density, the ratio estimator incorporates the effect of stem hollowing and fluting, because whole stem wood and branch density was calculated from the oven-dry mass divided by the over-bark volume of the biomass sample trees. Our results showed that the predictions of above ground biomass carbon (using Equation 8) based on ratio estimators of density, which average the density variation between trees within a species, agreed reasonably well with our above ground biomass data (Figure 1). The scatter in Figure 1 is hence largely due to tree-to-tree differences in density within a species and, to a lesser extent, stem form (from biomass trees). Other researchers recommended using species mean wood density in biomass regression equations [32], which is a conclusion largely supported by our analysis of New Zealand’s national wood density database. We note that the appropriate density measure has not always been used. For example, Basuki et al. [30] estimated above ground biomass by multiplying the volume of stems and large branches, calculated from stem sectional measurements over bark, by the specific gravity of wood and bark disks or sector samples along the stem. Specific gravity data overestimate stem dry weight if the stem is fluted, hollow, or has butt swell. The ratio estimator we developed using biomass sample trees reduced this potential source of bias, by adjusting the basic density data from breast height core samples.
We expect the tree fern equations to be reasonably robust, given the comparatively large sample size (80 tree ferns). Tree ferns at Maimai and Maungatautari sites were within the range found at Whakarewarewa; however, sampling would need to be extend to a broader range of sites to demonstrate how broadly applicable they are in New Zealand. Our tree fern root biomass study gave an overall average root/shoot ratio of 20%. We recommend that this ratio be applied to all tree fern species, because root biomass measurements were restricted to only four or five individuals per species and some species were not sampled at all.
The following sections provide a summary of the calculations steps needed to estimate the carbon stock in live biomass and dead wood pools.
4.1. Natural Forest Allometric Equations for Live Trees and Tree Ferns
Volume of the stem plus large branches ≥10 cm diameter Vstem+br≥10 (m3/tree) is estimated from DBH (cm) and total height H (m) using the mixed-species allometric equation:

Above ground biomass carbon per tree, Cagtree (kg/tree) is obtained by multiplying the estimated volume by the corresponding density assuming 50% of the mass is carbon, to which the mass of carbon in branches <10 cm in diameter over bark and foliage are added:
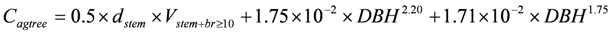
where dstem (kg.m−3) is the species-specific stem plus branch ≥10 cm diameter density over bark from the ratio estimator.
For tree ferns, supplementary wood density data cannot be obtained using coring methods. Tree fern above ground carbon Cagtrfn (kg/tree) is therefore estimated directly from DBH (cm) and H (m) as follows:
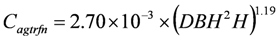
Below-ground biomass is estimated from above-ground biomass, using a root/shoot ratio of 25% for trees and 20% for tree ferns.
4.2. Equations for Standing Dead Stems and Logs
The volume of dead stems and truncated spars is estimated from DBH (cm), the expected total height when live H (m), and the measured spar height Hdead (m) in two steps using compatible volume and taper equations. Firstly, over-bark volume of an intact live stem Vstem (m3/tree) (i.e., excluding volume in large branches) with the same DBH and expected total height H as the dead stem is estimated as follows:
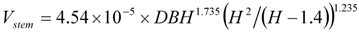
Secondly, the volume of a standing dead spar Vspar (m3/tree), is estimated from the estimated intact live stem volume up to the measured spar height using:
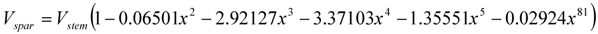
where x = (H − Hdead)/H.
The volume of dead spars is converted to carbon by multiplying by the species-specific density, assuming 50% of the mass is carbon, and a decay modifier, m (tabulated by decay class and species in Appendix 3 and in Coomes et al. [10]), to allow for density reductions due to decay. Hence, carbon in dead standing spars, Cspar is calculated as follows:
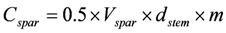
The equations for dead spars were not available when Richardson et al. [33] estimated dead wood in New Zealand’s natural forest. They applied the allometric volume equation developed for intact live stems to predict the volume of truncated spars from DBH and spar height, thereby significantly underestimating carbon in the dead wood pool.
A similar approach can be used to estimate carbon in logs lying on the forest floor (coarse woody debris from fallen and snapped stems and limbs), by firstly calculating log volume (VCWD) from the large and small end diameters and lengths of log and branch sections, using the formula for a truncated cone, which when converted to carbon is as follows:
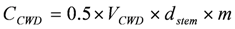
The measurement of dead wood volume on the forest floor can be onerous and difficult owing to fragmentation, and furthermore allocating decay classes is fraught with difficulties. Methods involving the use of structural equations also appear to be inadequate for estimating dead wood pools [33]. In the future, we anticipate that the allometric equations given in this paper will be linked to decay functions described in Beets et al. [17], to estimate carbon stocks and changes in the dead wood pool, without requiring direct measurements of this intractable pool. This linkage is possible through their common use of ratio estimators for density. This novel approach will allow predictions of carbon stocks and changes in dead wood (pool 3) of periodically remeasured plots, provided that the date of mortality can be estimated reasonably accurately.
5. Conclusions
The tree and tree fern biomass data described in this paper enabled the development of new allometric functions for predicting the above ground biomass carbon in natural forest in New Zealand from diameter and height measurements in plots. These equations can likely be further improved by measuring more biomass trees so that (1) species-specific volume equations can be developed for a range of species, and (2) species-specific ratio estimators for density can be developed. In addition, national wood density surveys could be extended to cover a wider range of species. Nevertheless, the new allometric equations for estimating above ground tree carbon from volume and density appear to provide reliable estimates for NZ’s natural forest even with relatively limited biomass data.
In comparison, above ground biomass measurements of an astoundingly large number of topical trees have been acquired in the past (approximately 2500 trees). It was clear from comparisons we made that the best allometric equation of Chave et al. [9] for tropical trees predicted above ground biomass carbon of our biomass trees surprisingly well in comparison with our natural forest equation. The additional tree sampling we undertook increased the sample size from 50 trees originally to 140 trees currently; however, the effect on carbon estimates was relatively small presumably because wood density, which was one of the most important variables influencing tree biomass, was well estimated using supplementary breast height outerwood density survey data.
A cost-effective approach for countries with limited biomass data for developing allometric equations may be to measure volume and the corresponding stem (and large branch) density, above ground biomass, and breast height outerwood density of approximately 200 sample trees selected across a range of species. The biomass data would need to be supplemented with national breast height outerwood basic density survey data across a range of important species. In addition site- and species-specific DBH/Height relations will need to be developed from plot inventory data as inputs to general allometric equations that estimate above ground biomass carbon from volume (from DBH and H) and wood density. Finally, consideration should be given on how best to estimate dead wood.
Above and below-ground biomass data for tree ferns showed that root/shoot ratios were on average approximately 20%, which is slightly lower than the default value of 25% used previously for natural forest by Coomes et al. [10]. Limited root biomass data exist for New Zealand trees and tree ferns. Root data should ideally be based on country-specific data, so additional root biomass studies are warranted, especially for New Zealand tree species.
Appendix 1
Diameter, height, volume, whole stem and branch wood plus bark density, basic density at breast height (BH), and biomass carbon data for live hardwood and softwood trees and tree ferns sampled from forest throughout New Zealand.
Appendix 2
Ratio estimators of species mean whole stem and branch >10 cm density (dstem) over bark derived from national wood density surveys. The number of trees sampled, and the p-values for regional density differences are given. If species is not tabulated, means by genus or morphology group should be used.
Acknowledgments
The Ministry for the Environment provided funding to compile biomass datasets and develop new allometric equations for LUCAS. Tree biomass studies were funded by the Ministry of Science and Technology, and the tree fern biomass study was funded by the Ministry of Primary Industries. We thank Scion staff who provided assistance with tree and tree fern biomass measurements over a number of years. The compatible volume and taper functions for estimating the truncated volume of dead standing trees were developed by Mina van der Colff, Scion.
Conflict of Interest
The authors declare no conflict of interest.
References
- Bouwman, A.F. Exchange of Greenhouse Gases between Terrestrial Ecosystems and the Atmosphere, Soils and the Greenhouse Effect. In Proceedings of the International Conference on Soils and the Greenhouse Effect, Wageningen, The Netherlands, 14-18 August 1989; Bouwman, A.F., Ed.; Wiley: Chichester, UK; pp. 61–192.
- Eamus, D.; McGuinness, K.; Burrows, W. Review of Allometric Relationships for Estimating Woody Biomass for Queensland, the Northern Territory and Western Australia; National Carbon Accounting System Technical Report 5A; Australian Greenhouse Office: Canberra , Australia, 2000; p. 56. [Google Scholar]
- Keith, H.; Barrett, D.; Keenan, R.J. Review of Allometric Relationships for Estimating Woody Biomass for New South Wales, the Australian Capital Territory, Victoria, Tasmania, and South Australia; National Carbon Accounting System Technical Report 5B; Australian Greenhouse Office: Canberra, Australia, 2000; p. 114. [Google Scholar]
- Ter-Mikaelian, M.T.; Korzukhin, M.D. Biomass equations for sixty-five North American tree species. For. Ecol. Manag. 1997, 97, 1–24. [Google Scholar] [CrossRef]
- Zianis, D.; Muukonen, P.; Mäkipää, R.; Mencuccini, M. Biomass and stem volume equations for tree species in Europe. In Silva Fennica; Monographs 4; The Finnish Society of Forest Science: Vantaa, Finland, 2005. [Google Scholar]
- Intergovernmental Panel on Climate Change (IPCC), IPCC Guidelines for National Greenhouse Gas Inventories; Intergovernmental Panel on Climate Change, Meteorological Office: Bracknell, UK, 1996.
- New Zealand’s Greenhouse Gas Inventory 1990-2012; Ministry for the Environment: Wellington, New Zealand, 2012; p. 438.
- Wardle, P. Vegetation of New Zealand; Cambridge Universtiy Press: Cambridge, UK, 1991. [Google Scholar]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef]
- Coomes, D.A.; Allen, R.B.; Scott, N.A.; Goulding, C.; Beets, P. Designing systems to monitor carbon stocks in forests and shrublands. For. Ecol. Manag. 2002, 164, 89–108. [Google Scholar] [CrossRef]
- Fehrmann, L.; Kleinn, C. General considerations about the use of allometric equations for biomass estimation on the example of Norway spruce in central Europe. For. Ecol. Manag. 2006, 236, 412–421. [Google Scholar] [CrossRef]
- Beets, P.N.; Brandon, A.; Fraser, B.V.; Goulding, C.J.; Lane, P.M.; Stephens, P.R. National Forest Inventories Reports: New Zealand. In National Forest Inventories—Pathways for Common Reporting; Tomppo, E., Gschwantner, T., Lawrence, M., McRoberts, R.E., Eds.; Springer: Berlin, Germany, 2010; pp. 391–410. [Google Scholar]
- Hall, G.M.J.; Wiser, S.K.; Allen, R.B.; Beets, P.N.; Goulding, C.J. Strategies to estimate national forest carbon stocks from inventory data: The 1990 New Zealand baseline. Glob. Chang. Biol. 2001, 7, 389–403. [Google Scholar] [CrossRef]
- Davis, M.R.; Wilde, R.H.; Garrett, L.; Oliver, G. New Zealand Carbon Monitoring System: Soil Data Collection Manual; Landcare Research New Zealand Limited and New Zealand Forest Research Institute Limited: Christchurch, New Zealand, 2004; pp. 1–56. [Google Scholar]
- Payton, I.J.; Newell, C.L.; Beets, P.N. New Zealand Carbon Monitoring System: Indigenous Forest and Shrubland Data Collection Manual; Caxton Press: Christchurch, New Zealand, 2004; pp. 1–68. [Google Scholar]
- Intergovernmental Panel on Climate Change (IPCC), Intergovernmental Panel on Climate Change. Good Practice Guidance for Land Use, Land-Use Change and Forestry; The Intergovernmental Panel on Climate Change (IPCC): Hayama, Japan, 2003.
- Beets, P.N.; Hood, I.A.; Kimberley, M.O.; Oliver, G.R.; Pearce, S.H.; Gardner, J.F. Coarse woody debris decay rates for seven indigenous tree species in the central North Island of New Zealand. For. Ecol. Manag. 2008, 256, 548–557. [Google Scholar] [CrossRef]
- Smale, M.C.; Bergin, D.O.; Gordon, A.D.; Pardy, G.F.; Steward, G.A. Selective logging of dense podocarp forest at Whirinaki: Early effects. N. Z. J. For. Sci. 1985, 15, 36–58. [Google Scholar]
- Smale, M.C.; Beveridge, A.E.; Herbert, J.W. Selection silviculture trials in North Island native forests: Impacts on the residual forest and their implications for sustainable forest management. N. Z. J. For. Sci. 1998, 42, 19–30. [Google Scholar]
- Madgwick, H.A.I.; Oliver, G.; Holten-Anderson, P. Above-ground biomass, nutrients, and energy content of trees in a second-growth stand old Agathis australis. N. Z. J. For. Sci. 1982, 12, 3–6. [Google Scholar]
- Beets, P.N. Amount and distribution of dry matter in a mature beech/podocarp community [in New Zealand]. N. Z. J. For. Sci. 1980, 10, 395–418. [Google Scholar]
- Rijkse, W.C. Soils of the Rotorua Lakes District, North Island, New Zealand; New Zealand Soil Survey Report 43; New Zealand Soil Bureau,Department of Scientific and Industrial Research: Wellington, New Zealand, 1979. [Google Scholar]
- Hewitt, A.E. New Zealand Soil Classification; Landcare Research Science Series No.1; Manaaki Whenua Press: Lincoln, GA, USA, 1998. [Google Scholar]
- Rowell, R. The Chemistry of Solid Wood; Advances in Chemistry Series, 207; American Chemical Society: Washington, DC, USA, 1984; p. 614. [Google Scholar]
- Cochran, W.G. Sampling Techniques, 3rd ed; John Wiley & Sons: New York, NY, USA, 1977. [Google Scholar]
- McCullagh, P.; Nelder, J.A. Generalised Linear Models, 2nd ed; Chapman and Hall: London, UK, 1989. [Google Scholar]
- Demaerschalk, J.P. Converting volume equations to compatible taper equations. For. Sci. 1972, 18, 241–245. [Google Scholar]
- Goulding, C.J.; Murray, J.C. Polynomial taper equations that are compatible with tree volume equations. N. Z. J. For. Sci. 1976, 5, 313–322. [Google Scholar]
- Gordon, A. Comparison of compatible polynomial taper equations. N. Z. J. For. Sci. 1983, 2, 146–155. [Google Scholar]
- Basuki, T.M.; van Laake, P.E.; Skidmore, A.K.; Hussin, Y.A. Allometric equations for estimating the above-ground biomass in tropical lowland Dipterocarp forests. For. Ecol. Manag. 2009, 257, 1684–1694. [Google Scholar] [CrossRef]
- Kenzo, T.; Furutani, R.; Hattori, D.; Kendawang, J.; Tanaka, S.; Sakurai, K.; Ninomiya, I. Allometric equations for accurate estimation of above-ground biomass in logged-over tropical rainforests in Sarawak, Malaysia. J. For. Res. 2009, 14, 365–372. [Google Scholar] [CrossRef]
- Brown, S.; Gillespie, A.J.R.; Lugo, A.E. Biomass estimation methods for tropical forests with applications to forest inventory data. For. Sci. 1989, 35, 881–902. [Google Scholar]
- Richardson, S.J.; Peltzer, D.A.; Hurst, J.M.; Allen, R.B.; Bellingham, P.J.; Carswell, F.E.; Clinton, P.W.; Griffiths, A.D.; Wiser, S.K.; Wright, E.F. Deadwood in New Zealand’s indigenous forests. For. Ecol. Manag. 2009, 258, 2456–2466. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).