
Linking Leaf Functional Traits to Aboveground Carbon Storage Across Successional Stages in Monsoon Evergreen Broad-Leaved Forests

Abstract
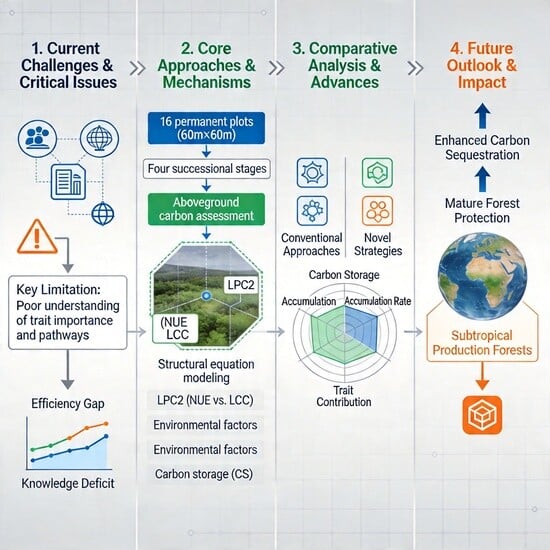
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design
2.3. Functional Trait Determination
2.4. Soil Sampling
2.5. Determination of Forest Carbon Stocks and Carbon Accumulation Rates
2.6. Data Analysis
3. Results
3.1. Carbon Sequestration Across Successional Stages
3.2. Functional Trait and Environmental Variation Across Successional Stages
3.2.1. Functional Trait Variation
3.2.2. Environmental Variation
3.3. Relationships Among Key Functional Traits, Environmental Factors, and Carbon Storage
4. Discussion
4.1. Changes in Carbon Sequestration During Succession
4.2. Changes in Plant Functional Traits and Resource Utilization Strategies During Succession
4.3. Plant Functional Trait and Environmental Effects on Carbon Sequestration During Succession
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pan, S.; Yu, P.T.; Wang, Y.H.; Wang, Z.; Yuan, C.X.; Yu, Z.J.; Hu, Y.Q.; Xiong, W.; Xu, L. Spatial distribution of carbon density for forest vegetation and the influencing factors in Liupan Mountains of Ningxia, NW China. Acta Ecol. Sin. 2014, 34, 6666–6677. [Google Scholar] [CrossRef]
- Zhang, Y.; Yi, A.J. Forest Carbon Sink Value Accounting and Related Issues Study in Chengde City. Innov. Sci. Technol. 2022, 22, 83–92. [Google Scholar] [CrossRef]
- McGarvey, J.C.; Thompson, J.R.; Epstein, H.E.; Shugart, H.H. Carbon storage in old-growth forests of the Mid-Atlantic: Toward better understanding the eastern forest carbon sink. Ecology 2015, 96, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Friedlingstein, P.; O’Sullivan, M.; Jones, M.W.; Andrew, R.M.; Hauck, J.; Olsen, A.; Peters, G.P.; Peters, W.; Pongratz, J.; Sitch, S.; et al. Global Carbon Budget 2020. Earth Syst. Sci. Data 2020, 12, 3269–3340. [Google Scholar] [CrossRef]
- Augusto, L.; Boca, A. Tree functional traits, forest biomass, and tree species diversity interact with site properties to drive forest soil carbon. Nat. Commun. 2022, 13, 1097. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Ma, L.; Chen, Y.; Tang, X. Characteristics of Soil Bacteria Community in Forests Along Monsoon Evergreen Broadleaved Forest Successional Sequence in Dinghushan National Nature Reserve. Ecol. Environ. Sci. 2022, 31, 2275–2282. [Google Scholar] [CrossRef]
- Liu, X.; Trogisch, S.; He, J.-S.; Niklaus, P.A.; Bruelheide, H.; Tang, Z.; Erfmeier, A.; Scherer-Lorenzen, M.; Pietsch, K.A.; Yang, B.; et al. Tree species richness increases ecosystem carbon storage in subtropical forests. Proc. R. Soc. B Biol. Sci. 2018, 285, 20181240. [Google Scholar] [CrossRef]
- Han, T.; Ren, H.; Hui, D.; Zhu, Y.; Lu, H.; Guo, Q.; Wang, J. Dominant ecological processes and plant functional strategies change during the succession of a subtropical forest. Ecol. Indic. 2023, 146, 109885. [Google Scholar] [CrossRef]
- Klumpp, K.; Soussana, J.-F. Using functional traits to predict grassland ecosystem change: A mathematical test of the response-and-effect trait approach. Glob. Change Biol. 2009, 15, 2921–2934. [Google Scholar] [CrossRef]
- Poorter, L.; Bongers, F. Leaf traits are good predictors of plant performance across 53 rain forest species. Ecology 2006, 87, 1733–1743. [Google Scholar] [CrossRef]
- Wang, M.; Lu, N.; An, N.; Fu, B. A Trait-Based Approach for Understanding Changes in Carbon Sequestration in Semi-Arid Grassland During Succession. Ecosystems 2022, 25, 155–171. [Google Scholar] [CrossRef]
- Matsuo, T.; Martinez-Ramos, M.; Onoda, Y.; Bongers, F.; Lohbeck, M.; Poorter, L. Light competition drives species replacement during secondary tropical forest succession. Oecologia 2024, 205, 1–11. [Google Scholar] [CrossRef]
- Díaz, S.; Cabido, M. Vive la différence: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Wang, P.; Sheng, L.X.; Yan, H.; Zhou, D.W.; Song, Y.T. Plant functional traits influence soil carbon sequestration in wetland ecosystem. Acta Ecol. Sin. 2010, 30, 6990–7000. [Google Scholar] [CrossRef]
- Rada, F.; Sarmiento, L.; Garcia-Varela, S. Plant functional traits along an old-field succession in the high tropical andes. Acta Oecol. 2021, 111, 103738. [Google Scholar] [CrossRef]
- An, H. Generality of leaf traits relationships of dominant species along the secondary succession in the Loess Plateau of China. Afr. J. Biotechnol. 2012, 11, 12240–12248. [Google Scholar] [CrossRef]
- Bauters, M.; Vercleyen, O.; Vanlauwe, B.; Six, J.; Bonyoma, B.; Badjoko, H.; Hubau, W.; Hoyt, A.; Boudin, M.; Verbeeck, H.; et al. Long-term recovery of the functional community assembly and carbon pools in an african tropical forest succession. Biotropica 2019, 51, 319–329. [Google Scholar] [CrossRef]
- Reich, P.B. The world-wide “fast-slow” plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Suter, M.; Edwards, P.J. Convergent succession of plant communities is linked to species’ functional traits. Perspect. Plant Ecol. Evol. Syst. 2013, 15, 217–225. [Google Scholar] [CrossRef]
- Li, Y.P.; Pan, L.J.; Chen, J.; Xu, H.; Yang, L.X. Response and influencing factors of leaf functional traits to forest succession in subtropical mixed plantations. Biodivers. Sci. 2024, 32, 6–18. [Google Scholar] [CrossRef]
- Zhang, P.; Ding, J.; Wang, Q.; McDowell, N.G.; Kong, D.; Tong, Y.; Yin, H. Contrasting coordination of non-structural carbohydrates with leaf and root economic strategies of alpine coniferous forests. New Phytol. 2024, 243, 580–590. [Google Scholar] [CrossRef]
- Li, Y.; Yang, F.; Ou, Y.; Zhang, D.; Liu, J.; Chu, G.; Zhang, Y.; Otieno, D.; Zhou, G. Changes in Forest Soil Properties in Different Successional Stages in Lower Tropical China. PLoS ONE 2013, 8, e81359. [Google Scholar] [CrossRef]
- Satdichanh, M.; Ma, H.; Yan, K.; Dossa, G.G.O.; Winowiecki, L.; Vagen, T.G.; Gassner, A.; Xu, J.; Harrison, R.D. Phylogenetic diversity correlated with above-ground biomass production during forest succession: Evidence from tropical forests in southeast Asia. J. Ecol. 2019, 107, 1419–1432. [Google Scholar] [CrossRef]
- Douma, J.C.; de Haan, M.W.A.; Aerts, R.; Witte, J.P.M.; van Bodegom, P.M. Succession-induced trait shifts across a wide range of NW European ecosystems are driven by light and modulated by initial abiotic conditions. J. Ecol. 2012, 100, 366–380. [Google Scholar] [CrossRef]
- Campetella, G.; Botta-Dukat, Z.; Wellstein, C.; Canullo, R.; Gatto, S.; Chelli, S.; Mucina, L.; Bartha, S. Patterns of plant trait-environment relationships along a forest succession chronosequence. Agric. Ecosyst. Environ. 2011, 145, 38–48. [Google Scholar] [CrossRef]
- Ali, A.; Yan, E.R.; Chen, H.Y.H.; Chang, S.X.; Zhao, Y.T.; Yang, X.D.; Xu, M.-S. Stand structural diversity rather than species diversity enhances aboveground carbon storage in secondary subtropical forests in Eastern China. Biogeosciences 2016, 13, 4627–4635. [Google Scholar] [CrossRef]
- Li, Y.; Bao, W.; Bongers, F.; Chen, B.; Chen, G.; Guo, K.; Jiang, M.; Lai, J.; Lin, D.; Liu, C.; et al. Drivers of tree carbon storage in subtropical forests. Sci. Total Environ. 2019, 654, 684–693. [Google Scholar] [CrossRef]
- Wang, N.; Lv, T.; Wang, L.; Chen, S.; Xie, L.; Fang, Y.; Ding, H. Leaf functional traits and ecological strategies of common plant species in evergreen broad-leaved forests on Huangshan Mountain. J. For. Res. 2024, 35, 130. [Google Scholar] [CrossRef]
- Wang, M.; Chen, Z.; Li, S.; Huang, X.; Lang, X.; Hu, Z.; Shang, R.; Liu, W. Spatial pattern of dominant species with different seed dispersal modes in a monsoon evergreen broad-leaved forest in Pu’er, Yunnan Province. Biodivers. Sci. 2023, 31, 23147. [Google Scholar] [CrossRef]
- Li, S.; Lang, X.; Huang, X.; Wang, Y.; Tang, R.; Liu, W.; Xu, C.; Su, J. Effects of plant diversity and big-sized trees on ecosystem function in a tropical montane evergreen broad-leaved forest. Front. Ecol. Evol. 2023, 11, 1188161. [Google Scholar] [CrossRef]
- Hao, M.; Dai, Y.; Yue, Q. Relationship between functional diversity of broadleaved Korean pine forest and forest carbon sink function. J. Beijing For. Univ. 2022, 44, 68–76. [Google Scholar] [CrossRef]
- Jiang, Q.; Jia, L.; Chen, Y.; Yang, Z.; Chen, S.; Xiong, D.; Liu, X.; Wang, X.; Yao, X.; Chen, T.; et al. Substrate and adenylate limit subtropical tree fine-root respiration under soil warming. Plant Cell Environ. 2023, 46, 2827–2840. [Google Scholar] [CrossRef]
- Wu, Z.L.; Dang, C.L. Biomass of the Pinus kesiya Royle ex Gordonforest in the Pu’er region of Yunnan, China. J. Yunnan Univ. 1992, 14, 119–127. [Google Scholar]
- Dang, C.L.; Wu, Z.L. Study on Biomass of the Castanopsis echidnocarpa Community in Monsoon Evergreen Broad-Leaved Forests. J. Yunnan Univ. 1992, 14, 95–107. [Google Scholar]
- Zhai, K.Y.; Yin, P.; Revillini, D.; Chen, X.L.; Liu, S.E.; Lou, Y.L.; Zhang, W.; Wang, S.; Zhou, G.; Delgado-Baquerizo, M. Subordinate tree species diversity explains carbon stock better than dominant tree diversity after decades of forest rewilding. J. Appl. Ecol. 2025, 62, 604–616. [Google Scholar] [CrossRef]
- Luo, W.; Kim, H.S.; Zhao, X.; Ryu, D.; Jung, I.; Cho, H.; Harris, N.; Ghosh, S.; Zhang, C.; Liang, J. New forest biomass carbon stock estimates in Northeast Asia based on multisource data. Glob. Change Biol. 2020, 26, 7045–7066. [Google Scholar] [CrossRef]
- Le Bagousse-Pinguet, Y.; Gross, N.; Maestre, F.T.; Maire, V.; de Bello, F.; Fonseca, C.R.; Kattge, J.; Valencia, E.; Leps, J.; Liancourt, P. Testing the environmental filtering concept in global drylands. J. Ecol. 2017, 105, 1058–1069. [Google Scholar] [CrossRef]
- Deng, F.; Xiao, L.; Huang, J.; Luo, H.; Zang, R. Changes in Leaf Functional Traits Driven by Environmental Filtration in Different Monsoon Tropical Forest Types. Forests 2023, 14, 2101. [Google Scholar] [CrossRef]
- Schweiger, A.K.; Schutz, M.; Risch, A.C.; Kneubuhler, M.; Haller, R.; Schaepman, M.E. How to predict plant functional types using imaging spectroscopy: Linking vegetation community traits, plant functional types and spectral response. Methods Ecol. Evol. 2017, 8, 86–95. [Google Scholar] [CrossRef]
- van der Sande, M.T.; Pena-Claros, M.; Ascarrunz, N.; Arets, E.J.M.M.; Licona, J.C.; Toledo, M.; Poorter, L. Abiotic and biotic drivers of biomass change in a Neotropical forest. J. Ecol. 2017, 105, 1223–1234. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Wen, D.; He, N. Forest carbon storage along the north-south transect of eastern China: Spatial patterns, allocation, and influencing factors. Ecol. Indic. 2016, 61, 960–967. [Google Scholar] [CrossRef]
- Ali, A.; Lin, S.; He, J.; Kong, F.; Yu, J.; Jiang, H. Big-sized trees overrule remaining trees’ attributes and species richness as determinants of aboveground biomass in tropical forests. Glob. Change Biol. 2019, 25, 2810–2824. [Google Scholar] [CrossRef]
- Lohbeck, M.; Poorter, L.; Martinez-Ramos, M.; Bongers, F. Biomass is the main driver of changes in ecosystem process rates during tropical forest succession. Ecology 2015, 96, 1242–1252. [Google Scholar] [CrossRef] [PubMed]
- Loreau, M.; Hector, A. Partitioning selection and complementarity in biodiversity experiments. Nature 2001, 413, 548. [Google Scholar] [CrossRef]
- Kelemen, A.; Tothmeresz, B.; Valko, O.; Miglecz, T.; Deak, B.; Torok, P. New aspects of grassland recovery in old-fields revealed by trait-based analyses of perennial-crop-mediated succession. Ecol. Evol. 2017, 7, 2432–2440. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.; Dang, H.; Yue, M.; Xu, J.; Zhang, L.; Quan, J.; Guo, Y.; Li, T.; Wang, L.; Wang, M.; et al. The role of intraspecific trait variability and soil properties in community assembly during forest secondary succession. Ecosphere 2019, 10, e02940. [Google Scholar] [CrossRef]
- Weigelt, A.; Mommer, L.; Andraczek, K.; Iversen, C.M.; Bergmann, J.; Bruelheide, H.; Fan, Y.; Freschet, G.T.; Guerrero-Ramírez, N.R.; Kattge, J.; et al. An integrated framework of plant form and function: The belowground perspective. New Phytol. 2021, 232, 42–59. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Poorter, L.; Rozendaal, D.M.A.; Bongers, F.; de Almeida-Cortez, J.S.; Zambrano, A.M.A.; Álvarez, F.S.; Andrade, J.L.; Villa, L.F.A.; Balvanera, P.; Becknell, J.M.; et al. Wet and dry tropical forests show opposite successional pathways in wood density but converge over time. Nat. Ecol. Evol. 2019, 3, 928–934. [Google Scholar] [CrossRef]
- Faccion, G.; Alves, A.M.; do Espirito-Santo, M.M.; Silva, J.O.; Sanchez-Azofeifa, A.; Ferreira, K.F. Intra- and interspecific variations on plant functional traits along a successional gradient in a Brazilian tropical dry forest. Flora 2021, 279, 151815. [Google Scholar] [CrossRef]
- Okubo, S.; Tomatsu, A.; Parikesit Muhamad, D.; Harashina, K.; Takeuchi, K. Leaf functional traits and functional diversity of multistoried agroforests in West Java, Indonesia. Agric. Ecosyst. Environ. 2012, 149, 91–99. [Google Scholar] [CrossRef]
- Lasky, J.R.; Uriarte, M.; Boukili, V.K.; Chazdon, R.L. Trait-mediated assembly processes predict successional changes in community diversity of tropical forests. Proc. Natl. Acad. Sci. USA 2014, 111, 5616–5621. [Google Scholar] [CrossRef] [PubMed]
- van der Plas, F.; Manning, P.; Allan, E.; Scherer-Lorenzen, M.; Verheyen, K.; Wirth, C.; Zavala, M.A.; Hector, A.; Ampoorter, E.; Baeten, L.; et al. Jack-of-all-trades effects drive biodiversity–ecosystem multifunctionality relationships in European forests. Nat. Commun. 2016, 7, 11109. [Google Scholar] [CrossRef] [PubMed]
- Onoda, Y.; Westoby, M.; Adler, P.B.; Choong, A.M.F.; Clissold, F.J.; Cornelissen, J.H.C.; Díaz, S.; Dominy, N.J.; Elgart, A.; Enrico, L.; et al. Global patterns of leaf mechanical properties. Ecol. Lett. 2011, 14, 301–312. [Google Scholar] [CrossRef]
- Díaz, S.; Kattge, J.; Cornelissen, J.H.C.; Wright, I.J.; Lavorel, S.; Dray, S.; Reu, B.; Kleyer, M.; Wirth, C.; Prentice, I.C.; et al. The global spectrum of plant form and function. Nature 2016, 529, 167–171. [Google Scholar] [CrossRef]
- Buzzard, V.; Hulshof, C.M.; Birt, T.; Violle, C.; Enquist, B.J. Re-growing a tropical dry forest: Functional plant trait composition and community assembly during succession. Funct. Ecol. 2016, 30, 1006–1013. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Trait | Unit | Ecological Significance |
|---|---|---|
| Leaf area (LA) | mm2 | Reflects a plant’s ability to acquire light. |
| Specific leaf area (SLA) | mm2 g−1 | Reflects a plant’s investment in light acquisition per unit area and its shade tolerance. |
| Leaf dry matter content (LDMC) | mg g−1 | Reflects the leaf energy-water balance and embodies plant growth-survival trade-off strategies. |
| Leaf carbon concentration (LC) | mg g−1 | Reflects the carbon accumulation rate. |
| Leaf nitrogen concentration (LN) | mg g−1 | Reflects a plant’s ability to acquire nitrogen and its maximum photosynthetic rate. |
| Leaf phosphorus concentration (LP) | mg g−1 | Affects plant growth and development, as well as ecosystem structure and function. |
| Leaf carbon–nitrogen ratio (LCNR) | % | Reflects the status of plant carbon and nitrogen metabolism, as well as nutrient use efficiency. |
| Wood density (WD) | g cm−3 | Affects plant growth, adaptability, and carbon storage capacity, while also playing a crucial role in ecosystem function and stability. |
| Tree Species Name | Organ | Allometric Growth Equation |
|---|---|---|
| Castanopsis echidnocarpa | Stem | W = 9.7566 + 0.014877 × DBH3 |
| Branch | W = 1.4497 + 0.0069051 × DBH3 | |
| Leaf | W = 0.03015231 × (−0.262 + DBH)2 | |
| Pinus kesiya var. langbianensis | Stem | W = 0.01218 × (DBH2H) × 0.9998 + 0.02340 × DBH2.4247 |
| Branch | W = 0.00028 × (DBH2H) × 1.2526 | |
| Leaf | W = DBH2H/(0.023 × DBH2H + 1967.57) | |
| Other broadleaf species | Stem | W = 0.080443 × DBH2.5142 |
| Branch | W = 0.00000029416 × (7.5074 + DBH)5 | |
| Leaf | W = 0.8442 × exp(0.1214 × DBH) − 0.9650 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Deng, F.; Qin, J.; Zhao, Y.; Liu, W. Linking Leaf Functional Traits to Aboveground Carbon Storage Across Successional Stages in Monsoon Evergreen Broad-Leaved Forests. Forests 2026, 17, 660. https://doi.org/10.3390/f17060660
Deng F, Qin J, Zhao Y, Liu W. Linking Leaf Functional Traits to Aboveground Carbon Storage Across Successional Stages in Monsoon Evergreen Broad-Leaved Forests. Forests. 2026; 17(6):660. https://doi.org/10.3390/f17060660
Chicago/Turabian StyleDeng, Fuying, Jiali Qin, Yuhan Zhao, and Wande Liu. 2026. "Linking Leaf Functional Traits to Aboveground Carbon Storage Across Successional Stages in Monsoon Evergreen Broad-Leaved Forests" Forests 17, no. 6: 660. https://doi.org/10.3390/f17060660
APA StyleDeng, F., Qin, J., Zhao, Y., & Liu, W. (2026). Linking Leaf Functional Traits to Aboveground Carbon Storage Across Successional Stages in Monsoon Evergreen Broad-Leaved Forests. Forests, 17(6), 660. https://doi.org/10.3390/f17060660