

Prediction of Potential Forest Risk Areas for Phytopythium helicoides in China Under Climate Change Based on Maximum Entropy Modeling


Abstract
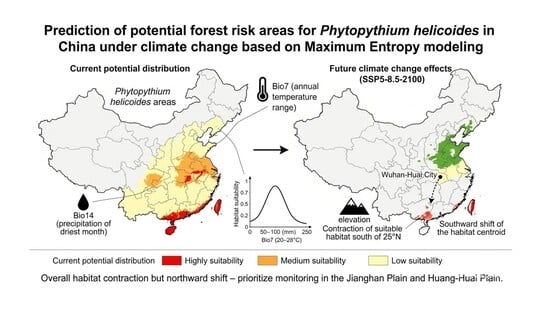
1. Introduction
2. Materials and Methods
2.1. Species Occurrence Data
2.2. Data Acquisition and Processing
2.3. Study Area and Background Extent (M Region)
2.4. Background Point Selection Strategy
2.5. Construction of the MaxEnt Model
2.6. Model Accuracy Testing
2.7. Optimization of the MaxEnt Model
2.8. Classification of Suitable Areas
2.9. Changes in Potential Suitable Areas and Center of Gravity Shift
3. Results and Analysis
3.1. Model Performance Results
3.2. Selection of Key Environmental Factors
3.3. Key Environmental Drivers of P. helicoides Distribution
3.4. Present Suitability Map for P. helicoides
3.5. Future Climate Distribution of P. helicoides
3.6. Centroid Migration of P. helicoides
4. Discussion
4.1. Dominance of Bio14: The Dry-Month Precipitation Bottleneck
4.2. Southern Contraction Under High-Emission Scenarios
4.3. Local Expansion Within Overall Contraction
4.4. Differences and Similarities with Traditional Views
4.5. Stable Centroid Versus Shifting Boundaries
4.6. Refining Traditional Views on Pathogen Range Shifts
4.7. Implications for Forest Disease Management and Quarantine
5. Considerations of Model Uncertainty
5.1. Climate Model Source and Limitations
5.2. Extrapolation Risk and Model Transferability
5.3. Static Assumptions for Non-Climatic Variables
5.4. Small-Sample Bias and Model Generalization
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ghimire, B.; Baysal-Gurel, F. A Diagnostic Guide to Phytopythium helicoides and Phytopythium vexans Causing Root and Crown Rot Diseases. Plant Health Prog. 2023, 24, 527–538. [Google Scholar] [CrossRef]
- Weiland, J.E.; Beck, B.R.; Davis, A. Pathogenicity and Virulence of Pythium Species Obtained from Forest Nursery Soils on Douglas-Fir Seedlings. Plant Dis. 2013, 97, 744–748. [Google Scholar] [CrossRef]
- Legrifi, I.; Taoussi, M.; Figuigui, J.A.; Lazraq, A.; Hussain, T.; Lahlali, R. Oomycetes Root Rot Caused by Pythium spp. and Phytophthora spp.: Host Range, Detection, and Management Strategies, Special Case of Olive Trees. J. Crop Health 2024, 76, 19–47. [Google Scholar] [CrossRef]
- Jesus, A.L.D.; GonçAlves, D.R.; Rocha, S.C.O.; Marano, A.V.; Jer?Nimo, G.H.; Souza, J.I.D.; Castilho Boro, M.; Pires-Zottarelli, C.L.A. Morphological and Phylogenetic Analyses of Three Phytopythium Species (Peronosporales, Oomycota) from Brazil. Cryptogam. Mycol. 2016, 37, 117–128. [Google Scholar] [CrossRef]
- Jung, T.; Pérez-Sierra, A.; Durán, A.; Horta, J.M.; Balci, Y.; Scanu, B. Canker and decline diseases caused by soil- and airborne Phytophthora species in forests and woodlands. Persoonia-Mol. Phylogeny Evol. Fungi 2018, 40, 182–220. [Google Scholar] [CrossRef]
- Gadelha, L.; Siracusa, P.C.D.; Dalcin, E.; Silva, L.A.E.; Siqueira, M.F.D. A survey of biodiversity informatics: Concepts, practices, and challenges. Wiley Interdiscip. Rev. Data Min. Knowl. Discov. 2020, 2020, e1394. [Google Scholar] [CrossRef]
- Hu, X.; Shi, Y.; Song, B.; Hu, L.; Zhao, Q.; Pang, D.; Yin, J.; Li, Y. Habitat distribution of major wildlife in the Sanjiangyuan National park under climate scenarios based on maxent modeling. Sci. Rep. 2025, 15, 39789. [Google Scholar] [CrossRef] [PubMed]
- Chiou, K.L.; Blair, M.E. Modeling niches and mapping distributions: Progress and promise of ecological niche models for primate research. In Spatial Analysis in Field Primatology: Applying GIS at Varying Scales; Cambridge University Press: Cambridge, UK, 2015. [Google Scholar]
- Davoudi Moghaddam, D.; Rahmati, O.; Haghizadeh, A.; Kalantari, Z. A modeling comparison of groundwater potential mapping in a mountain bedrock aquifer: QUEST, GARP, and RF models. Water 2020, 12, 679. [Google Scholar] [CrossRef]
- Tang, X.; Deng, Y.; He, Z.; Zhou, M.; Yuan, Y.; Zeng, K. Modelling the potential distribution and niche shift of Solenopsis invicta Buren under climate change and invasion process. Front. For. Glob. Change 2025, 8, 1659630. [Google Scholar] [CrossRef]
- Sabir, I.A.; Li, C.; Xu, C.; Hu, G.; Qin, Y. Agronomic and Environmental Suitability of ‘Tabtim Siam’ and ‘Xishi’ Pomelo in Dabu County (China). Horticulturae 2025, 11, 1290. [Google Scholar] [CrossRef]
- Gao, B.; Yuan, S.; Guo, Y.; Zhao, Z. Potential geographical distribution of Actinidia spp. and its predominant indices under climate change. Ecol. Inform. 2022, 72, 101865. [Google Scholar] [CrossRef]
- Pradhan, A.; Adhikari, D. Predicting the distribution of suitable habitats for pandanus unguifer hook.f.—A dwarf endemic species from sikkim himalayas, through ecological niche modeling. Int. J. Conserv. Sci. 2020, 11, 145–152. [Google Scholar]
- Liu, Y.; Yang, Q.; Li, S.; Zhang, Y.; Xiang, Y.; Yang, Y.; Zhang, J. Spatiotemporal Dynamics of Ilex macrocarpa Distribution Under Future Climate Scenarios: Implications for Conservation Planning. Forests 2025, 16, 370. [Google Scholar] [CrossRef]
- Li, J.; Deng, C.; Duan, G.; Wang, Z.; Zhang, Y.; Fan, G. Potentially suitable habitats of Daodi goji berry in China under climate change. Front. Plant Sci. 2023, 14, 1279019. [Google Scholar] [CrossRef]
- Morin, C.W.; Semenza, J.C.; Trtanj, J.M.; Glass, G.E.; Boyer, C.; Ebi, K.L. Unexplored Opportunities: Use of Climate- and Weather-Driven Early Warning Systems to Reduce the Burden of Infectious Diseases. Curr. Environ. Health Rep. 2018, 5, 430–438. [Google Scholar] [CrossRef]
- Ren, L.Y.; Wen, K.; Cheng, B.P.; Jin, J.H.; Srivastava, V.; Chen, X.R. Rapid detection of the phytopathogenic oomycete Phytopythium helicoides with a visualized loop-mediated isothermal amplification assay. Braz. J. Microbiol. 2025, 56, 563–572. [Google Scholar] [CrossRef]
- Gallego-Tévar, B.; Gil-Martínez, M.; Perea, A.; Pérez-Ramos, I.M.; Gómez-Aparicio, L. Interactive Effects of Climate Change and Pathogens on Plant Performance: A Global Meta-Analysis. Glob. Change Biol. 2024, 30, e17535. [Google Scholar] [CrossRef] [PubMed]
- Shikano, I.; Cory, J.S. Impact of Environmental Variation on Host Performance Differs with Pathogen Identity: Implications for Host-Pathogen Interactions in a Changing Climate. Sci. Rep. 2015, 5, 15351. [Google Scholar] [CrossRef]
- Chang, X.; Feng, S.; Ullah, F.; Zhang, Y.; Zhang, Y.; Qin, Y.; Nderitu, J.H.; Dong, Y.; Huang, W.; Zhang, Z.; et al. Adapting distribution patterns of desert locusts, Schistocerca gregaria in response to global climate change. Bull. Entomol. Res. 2025, 115, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Lv, L.; Bao, S.; Cai, Z.; Fu, S.; Shi, J. Evaluation and Application of the MaxEnt Model to Quantify L. nanum Habitat Distribution Under Current and Future Climate Conditions. Agronomy 2025, 15, 1869. [Google Scholar] [CrossRef]
- Van de Vuurst, P.; Qiao, H.; Soler-Tovar, D.; Escobar, L.E. Climate change linked to vampire bat expansion and rabies virus spillover. Ecography 2024, 2024, e06714. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Dudík, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef]
- Shen, S.; Zhang, X.; Jian, S. The Distributional Range Changes of European Heterobasidion Under Future Climate Change. Forests 2024, 15, 1863. [Google Scholar] [CrossRef]
- Ma, R.; Li, C.; Tian, H.; Zhang, Y.; Feng, X.; Li, J.; Hu, W. The current distribution of tick species in Inner Mongolia and inferring potential suitability areas for dominant tick species based on the MaxEnt model. Parasit. Vectors 2023, 16, 286. [Google Scholar] [CrossRef]
- Zhang, J.; Li, X.; Li, S.; Yang, Q.; Li, Y.; Xiang, Y.; Yao, B. MaxEnt Modeling of Future Habitat Shifts of Itea yunnanensis in China Under Climate Change Scenarios. Biology 2025, 14, 899. [Google Scholar] [CrossRef]
- Fu, C.; Wen, X.; Shi, Z.; Rui, L.; Jiang, N.; Zhao, G.; Wang, R.; Zhao, J.; Yang, Y. Potential distribution prediction of Ceracris kiangsu Tsai in China. Sci. Rep. 2024, 14, 13375. [Google Scholar] [CrossRef]
- Chen, S.; Jiang, Z.; Song, J.; Xie, T.; Xue, Y.; Yang, Q. Prediction of potential habitat of Verbena officinalis in China under climate change based on optimized MaxEnt model. Front. Plant Sci. 2025, 16, 1563070. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Deng, Z. Analysis of a Potentially Suitable Habitat for Solanum aculeatissimum in Southwest China Under Climate Change Scenarios. Plants 2025, 14, 1979. [Google Scholar] [CrossRef]
- Dong, P.; Wang, L.; Wang, L.; Lei, M.; Qiu, D.; Bai, G.; Guo, F.-X.; Chen, Y. Investigating the distributional response of the rare and endangered plant Fritillaria przewalskii to climate change based on optimized MaxEnt model. Sci. Rep. 2025, 15, 35939. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Yang, Y.; Peng, X.; Wang, J.; Wu, M.; Zhang, Y.; Liu, X.; Peng, P. Habitat Suitability and Driving Factors of Cycas panzhihuaensis in the Hengduan Mountains. Plants 2025, 14, 2797. [Google Scholar] [CrossRef]
- Jaturapruek, R.; Fontaneto, D.; Maiphae, S. The influence of environmental variables on bdelloid rotifers of the genus Rotaria in Thailand. J. Trop. Ecol. 2020, 36, 267–274. [Google Scholar] [CrossRef]
- Fan, Y.; Zhang, X.; Yang, J.; Yang, J.; Zhang, H.; Yang, B.; Li, X. Prediction of global potential distribution and assessment of habitat suitability for Xanthium spinosum driven by climate change. Front. Plant Sci. 2025, 16, 1690546. [Google Scholar] [CrossRef]
- Xiao, Y.; Guo, Q.; Xie, N.; Yuan, G.; Liao, M.; Gui, Q.; Ding, G. Predicting the global potential distribution of Bursaphelenchus xylophilus using an ecological niche model: Expansion trend and the main driving factors. BMC Ecol. Evol. 2024, 24, 48. [Google Scholar] [CrossRef]
- McCoy, A.G.; Belanger, R.R.; Bradley, C.A.; Cerritos-Garcia, D.G.; Garnica, V.C.; Giesler, L.J.; Grijalba, P.E.; Guillin, E.; Henriquez, M.A.; Kim, Y.M. A global-temporal analysis on Phytophthora sojae resistance-gene efficacy. Nat. Commun. 2023, 14, 6043. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadi, A.; Al-Masoudi, R.; Al-Habsi, N.; Al-Said, F.; Al-Rawahy, S.; Ahmed, M.; Deadman, M. Effect of salinity on pythium damping-off of cucumber and on the tolerance of Pythium aphanidermatum. Plant Pathol. 2010, 59, 112–120. [Google Scholar] [CrossRef]
- Gsell, A.S.; Biere, A.; de Boer, W.; de Bruijn, I.; Eichhorn, G.; Frenken, T.; Geisen, S.; van Der Jeugd, H.; Mason-Jones, K.; Meisner, A. Environmental refuges from disease in host–parasite interactions under global change. Ecology 2023, 104, e4001. [Google Scholar] [CrossRef] [PubMed]
- Urayama, S.-I.; Takaki, Y.; Nunoura, T. FLDS: A comprehensive dsRNA sequencing method for intracellular RNA virus surveillance. Microbes Environ. 2016, 31, 33–40. [Google Scholar] [CrossRef]
- Bebber, D.P.; Ramotowski, M.A.; Gurr, S.J. Crop pests and pathogens move polewards in a warming world. Nat. Clim. Change 2013, 3, 985–988. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Variable | Decription | Perent Contribution | Permutation Importance |
|---|---|---|---|
| Bio14 | The lowest monthly precipitation | 72.4 | 35.8 |
| Bio7 | Annual range of temperature | 8.1 | 5.3 |
| elev | elevation | 5.7 | 32 |
| t_gravel | Percentage of crushed stone volume | 5.4 | 13.8 |
| Bio13 | The monthly precipitation of the wettest month | 3.2 | 4.2 |
| t_texture | Topsoil texture | 1.9 | 2.5 |
| slope | Slope gradient | 1.9 | 2.1 |
| t_esp | Slope orientation | 0.4 | 1.8 |
| bio15 | Seasonal variation in precipitation | 0.4 | 0.9 |
| aspect | Aspect | 0.3 | 1 |
| t_silt | Silt content | 0.2 | 0.6 |
| Climate Change Scenario | Year | AUC Value |
|---|---|---|
| Current | 1970–2000 | 0.974 |
| 2021–2040 | 0.976 | |
| 2041–2060 | 0.978 | |
| scenario SSP1-2.6 | 2061–2080 | 0.976 |
| 2081–2100 | 0.977 | |
| 2021–2040 | 0.975 | |
| 2041–2060 | 0.979 | |
| scenario SSP3-7.0 | 2061–2080 | 0.978 |
| 2081–2100 | 0.978 | |
| 2021–2040 | 0.979 | |
| 2041–2060 | 0.975 | |
| scenario SSP5-8.5 | 2061–2080 | 0.979 |
| 2081–2100 | 0.976 |
| Climate Scenario | Area Unit (×104 km2) | ||||
|---|---|---|---|---|---|
| Unsuitable Habitat | Sightly Suitable Habitat | Moderately Suitable Habitat | Highly Suitable Habitat | Total Suitable Habitat | |
| SSP1-2.6 2021–2040 | 755.03 | 87.49 | 62.84 | 33.08 | 183.41 |
| SSP1-2.6 2041–2060 | 772.90 | 82.75 | 52.15 | 30.64 | 165.54 |
| SSP1-2.6 2061–2080 | 755.81 | 90.75 | 57.13 | 34.75 | 182.63 |
| SSP1-2.6 2081–2100 | 762.40 | 85.53 | 53.73 | 36.77 | 176.03 |
| SSP3-7.0 2021–2040 | 739.56 | 86.49 | 67.00 | 45.38 | 198.87 |
| SSP3-7.0 2041–2060 | 774.10 | 79.93 | 54.65 | 29.76 | 164.34 |
| SSP3-7.0 2061–2080 | 756.34 | 94.19 | 54.12 | 33.77 | 182.08 |
| SSP3-7.0 2081–2040 | 761.67 | 91.59 | 53.03 | 32.15 | 176.77 |
| SSP5-8.5 2021–2040 | 752.47 | 90.99 | 60.65 | 34.32 | 185.96 |
| SSP5-8.5 2041–2060 | 753.97 | 88.94 | 60.73 | 34.80 | 184.47 |
| SSP5-8.5 2061–2080 | 748.46 | 92.86 | 58.32 | 38.79 | 189.97 |
| SSP5-8.5 2081–2100 | 763.71 | 84.95 | 56.80 | 32.99 | 174.74 |
| Decades Scenarios | Predicted Area Unit: ×104 km2 | ||
|---|---|---|---|
| Suitable | Contraction | Gain | |
| 2021–2040 SSP1-2.6 vs. current | 172.804 | 11.005 | 10.380 |
| 2041–2060 SSP1-2.6 vs. current | 161.250 | 22.498 | 4.619 |
| 2061–2080 SSP1-2.6 vs. current | 172.493 | 11.347 | 9.949 |
| 2081–2100 SSP1-2.6 vs. current | 168.389 | 15.418 | 7.256 |
| 2021–2040 SSP3-7.0 vs. current | 178.929 | 4.974 | 19.304 |
| 2041–2060 SSP3-7.0 vs. current | 161.517 | 22.252 | 2.670 |
| 2061–2080 SSP3-7.0 vs. current | 169.203 | 14.616 | 12.635 |
| 2081–2100 SSP3-7.0 vs. current | 169.158 | 14.597 | 7.651 |
| 2021–2040 SSP5-8.5 vs. current | 174.012 | 9.813 | 11.597 |
| 2041–2060 SSP5-8.5 vs. current | 173.517 | 10.347 | 10.849 |
| 2061–2080 SSP5-8.5 vs. current | 173.042 | 10.800 | 16.842 |
| 2081–2100 SSP5-8.5 vs. current | 168.188 | 15.599 | 6.701 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Kong, Y.; Jiao, B.; Dai, S.; Yang, C.; Chen, Q.; Dai, T. Prediction of Potential Forest Risk Areas for Phytopythium helicoides in China Under Climate Change Based on Maximum Entropy Modeling. Forests 2026, 17, 626. https://doi.org/10.3390/f17050626
Kong Y, Jiao B, Dai S, Yang C, Chen Q, Dai T. Prediction of Potential Forest Risk Areas for Phytopythium helicoides in China Under Climate Change Based on Maximum Entropy Modeling. Forests. 2026; 17(5):626. https://doi.org/10.3390/f17050626
Chicago/Turabian StyleKong, Yuzhe, Binbin Jiao, Size Dai, Chun Yang, Qing Chen, and Tingting Dai. 2026. "Prediction of Potential Forest Risk Areas for Phytopythium helicoides in China Under Climate Change Based on Maximum Entropy Modeling" Forests 17, no. 5: 626. https://doi.org/10.3390/f17050626
APA StyleKong, Y., Jiao, B., Dai, S., Yang, C., Chen, Q., & Dai, T. (2026). Prediction of Potential Forest Risk Areas for Phytopythium helicoides in China Under Climate Change Based on Maximum Entropy Modeling. Forests, 17(5), 626. https://doi.org/10.3390/f17050626