
Mapping the Distribution of Viruses in Wild Apple Populations in the Southeast Region of Kazakhstan
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Botanical Description and Sampling
2.2. Assessment of Disease Distribution
2.3. RNA Isolation and RT-qPCR Detection
2.4. Mapping of Infected Apple Trees
3. Results
3.1. Botanical and Phytopathological Descriptions of Wild Apple Populations
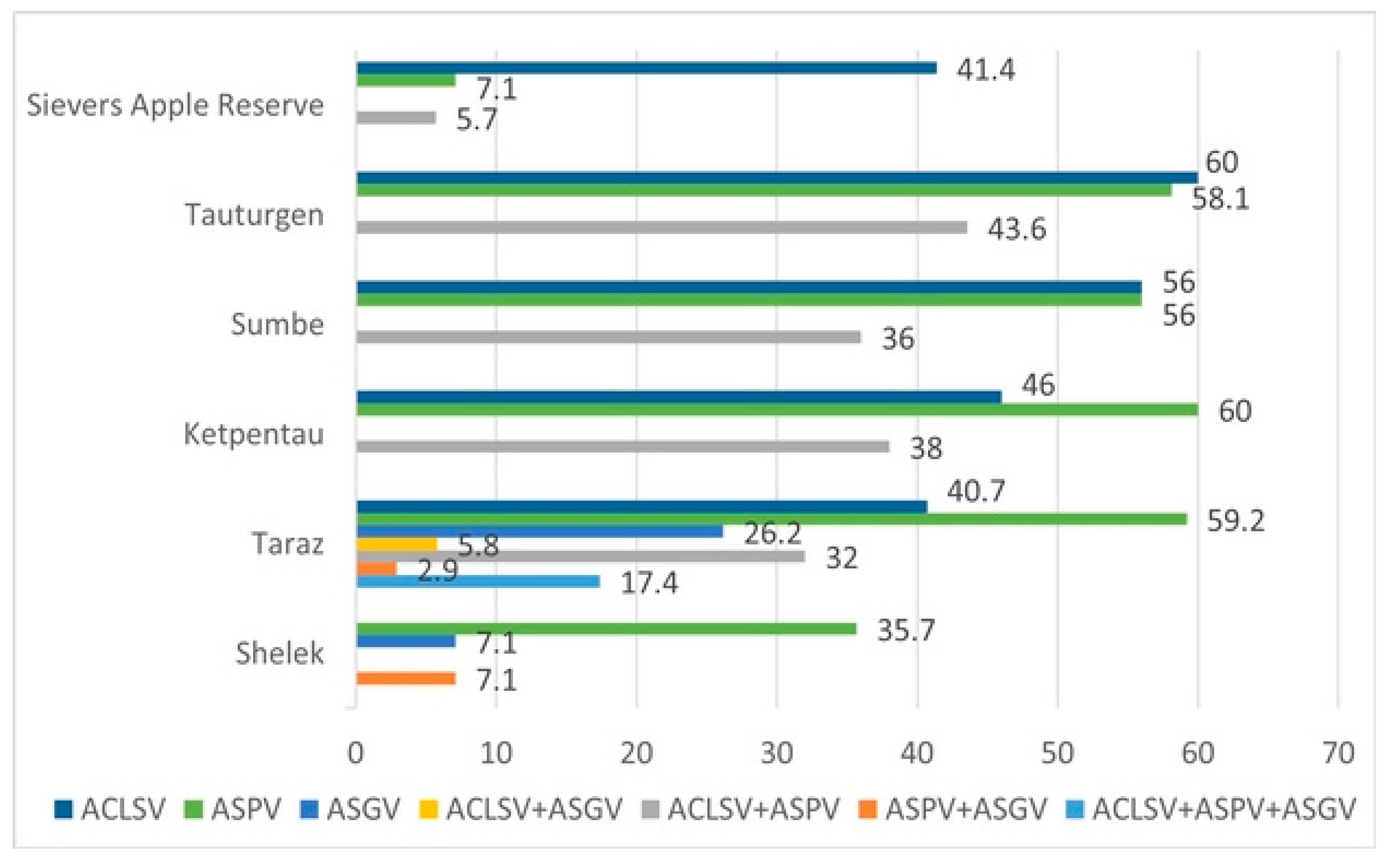
3.2. Distribution of Viruses in Wild Apple Populations
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Panyushkina, I.; Mukhamadiev, N.; Lynch, A.; Ashikbaev, N.; Arizpe, A.; O’Connor, C.; Abjanbaev, D.; Mengdibayeva, G.; Sagitov, A. Wild Apple Growth and Climate Change in Southeast Kazakhstan. Forests 2017, 8, 406. [Google Scholar] [CrossRef]
- Janick, J. (Ed.) Horticultural Reviews: Volume 29: Wild Apple and Fruit Trees of Central Asia; Horticultural Reviews; Wiley: New York, NY, USA; Chichester, UK, 2003; ISBN 978-0-471-21968-2. [Google Scholar]
- Taskuzhina, A.; Pozharskiy, A.; Gritsenko, D. The Contribution of Malus sieversii to the Emergence and Diversity of Domesticated Apple Varieties. In Malus Domestica—New Insights; Ebru Yaşa Kafkas, N., İmrak, B., Eds.; IntechOpen: London, UK, 2024; ISBN 978-0-85014-766-7. [Google Scholar]
- Cornille, A.; Giraud, T.; Smulders, M.J.M.; Roldán-Ruiz, I.; Gladieux, P. The Domestication and Evolutionary Ecology of Apples. Trends Genet. 2014, 30, 57–65. [Google Scholar] [CrossRef]
- Feurtey, A.; Guitton, E.; De Gracia Coquerel, M.; Duvaux, L.; Shiller, J.; Bellanger, M.-N.; Expert, P.; Sannier, M.; Caffier, V.; Giraud, T.; et al. Asian Wild Apples Threatened by Gene Flow from Domesticated Apples and by Their Pestified Pathogen. Available online: https://www.biorxiv.org/content/10.1101/2020.04.15.042242v1 (accessed on 5 March 2025). [CrossRef]
- Feurtey, A.; Guitton, E.; De Gracia Coquerel, M.; Duvaux, L.; Shiller, J.; Bellanger, M.; Expert, P.; Sannier, M.; Caffier, V.; Giraud, T.; et al. Threat to Asian Wild Apple Trees Posed by Gene Flow from Domesticated Apple Trees and Their “Pestified” Pathogens. Mol. Ecol. 2020, 29, 4925–4941. [Google Scholar] [CrossRef]
- Tian, Z.; Song, H.; Wang, Y.; Li, J.; Maimaiti, M.; Liu, Z.; Zhang, H.; Zhang, J. Wild Apples Are Not That Wild: Conservation Status and Potential Threats of Malus sieversii in the Mountains of Central Asia Biodiversity Hotspot. Diversity 2022, 14, 489. [Google Scholar] [CrossRef]
- Uzun, A.; Turgunbaev, K.; Pınar, H.; Yılmaz, K.U. Apple genetic resources in Kyrgyzstan geography: Determination, evaluation, and conservation. Int. J. Agric. Nat. Sci. 2022, 15, 221–225. [Google Scholar]
- Xu, X.; Huai, W.; Hamiti; Zhang, X.; Zhao, W. Phytophthora Species from Xinjiang Wild Apple Forests in China. Forests 2019, 10, 927. [Google Scholar] [CrossRef]
- Velasco, R.; Zharkikh, A.; Affourtit, J.; Dhingra, A.; Cestaro, A.; Kalyanaraman, A.; Fontana, P.; Bhatnagar, S.K.; Troggio, M.; Pruss, D.; et al. The Genome of the Domesticated Apple (Malus × Domestica Borkh.). Nat. Genet. 2010, 42, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Tegtmeier, R.; Švara, A.; Gritsenko, D.; Khan, A. Malus sieversii: A Historical, Genetic, and Conservational Perspective of the Primary Progenitor Species of Domesticated Apples. Hortic. Res. 2025, 12, uhae244. [Google Scholar] [CrossRef]
- Duan, N.; Bai, Y.; Sun, H.; Wang, N.; Ma, Y.; Li, M.; Wang, X.; Jiao, C.; Legall, N.; Mao, L.; et al. Genome Re-Sequencing Reveals the History of Apple and Supports a Two-Stage Model for Fruit Enlargement. Nat. Commun. 2017, 8, 249. [Google Scholar] [CrossRef]
- Bus, V.G.M.; Laurens, F.N.D.; Van De Weg, W.E.; Rusholme, R.L.; Rikkerink, E.H.A.; Gardiner, S.E.; Bassett, H.C.M.; Kodde, L.P.; Plummer, K.M. The Vh8 Locus of a New Gene-for-gene Interaction between Venturia inaequalis and the Wild Apple Malus sieversii Is Closely Linked to the Vh2 Locus in Malus Pumila R12740-7A. New Phytol. 2005, 166, 1035–1049. [Google Scholar] [CrossRef]
- Bus, V.G.M.; Rikkerink, E.H.A.; Caffier, V.; Durel, C.-E.; Plummer, K.M. Revision of the Nomenclature of the Differential Host-Pathogen Interactions of Venturia inaequalis and Malus. Annu. Rev. Phytopathol. 2011, 49, 391–413. [Google Scholar] [CrossRef]
- Kostick, S.A.; Norelli, J.L.; Evans, K.M. Novel Metrics to Classify Fire Blight Resistance of 94 Apple Cultivars. Plant Pathol. 2019, 68, 985–996. [Google Scholar] [CrossRef]
- Barba, M.; Ilardi, V.; Pasquini, G. Control of Pome and Stone Fruit Virus Diseases. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2015; Volume 91, pp. 47–83. ISBN 978-0-12-802762-2. [Google Scholar]
- Wunsch, A.O.; Miranda Sazo, M.; Van Zoeren, J.; Lamour, K.H.; Hurtado-Gonzales, O.P.; Khan, A.; Fuchs, M. Investigating the Role of Viruses in the Rapid Decline of Young Apple Trees in High-Density Orchards in New York. Plants 2024, 13, 2866. [Google Scholar] [CrossRef] [PubMed]
- Grimová, L.; Winkowska, L.; Zíka, L.; Ryšánek, P. Distribution of Viruses in Old Commercial and Abandoned Orchards and Wild Apple Trees. J. Plant Pathol. 2016, 98, 549–554. [Google Scholar] [CrossRef]
- Kuzmitskaya, P.V.; Urbanovich, O.Y. Distribution and genetic diversity of three apple viruses in Belarus. Vestn. VOGiS 2016, 20, 673–682. [Google Scholar] [CrossRef]
- Harshman, J.M.; Evans, K.M.; Allen, H.; Potts, R.; Flamenco, J.; Aldwinckle, H.S.; Wisniewski, M.E.; Norelli, J.L. Fire Blight Resistance in Wild Accessions of Malus sieversii. Plant Dis. 2017, 101, 1738–1745. [Google Scholar] [CrossRef]
- Collum, T.D.; Stone, A.L.; Sherman, D.J.; Damsteegt, V.D.; Schneider, W.L.; Rogers, E.E. Viral Reservoir Capacity of Wild Prunus Alternative Hosts of Plum Pox Virus Through Multiple Cycles of Transmission and Dormancy. Plant Dis. 2022, 106, 101–106. [Google Scholar] [CrossRef]
- Rana, T.; Chandel, V.; Hallan, V.; Zaidi, A.A. Himalayan Wild Cherry (Prunus cerasoides D. Don): A New Host of Apple Chlorotic Leaf Spot Virus. For. Pathol. 2008, 38, 73–77. [Google Scholar] [CrossRef]
- Katsiani, A.T.; Maliogka, V.I.; Candresse, T.; Katis, N.I. Host-range Studies, Genetic Diversity and Evolutionary Relationships of ACLSV Isolates from Ornamental, Wild and Cultivated Rosaceous Species. Plant Pathol. 2014, 63, 63–71. [Google Scholar] [CrossRef]
- Shi, W.; Yao, R.; Sunwu, R.; Huang, K.; Liu, Z.; Li, X.; Yang, Y.; Wang, J. Incidence and Molecular Identification of Apple Necrotic Mosaic Virus (ApNMV) in Southwest China. Plants 2020, 9, 415. [Google Scholar] [CrossRef]
- EPPO Global Database. Available online: https://https://gd.eppo.int/ (accessed on 29 January 2025).
- Jashenko, R.; Tanabekova, G.; Lu, Z. Assessment of Biological and Ecological Characteristics of Sievers Apple Tree Pests in Trans-Ili Alatau, Kazakhstan. Sustainability 2023, 15, 11303. [Google Scholar] [CrossRef]
- Svoboda, J.; Polák, J. Relative Concentration of Apple Mosaic Virus Coat Protein in Different Parts of Apple Tree. Hortic. Sci. 2010, 37, 22–26. [Google Scholar] [CrossRef]
- Alemzadeh, E.; Katsiani, A.T.; Efthimiou, K.; Katis, N.I. Occurrence of Apple Chlorotic Leaf Spot Virus in Apple and Quince in Southern Iran. J. Plant Pathol. 2016, 98, 171–185. [Google Scholar] [CrossRef]
- Rana, T.; Chandel, V.; Hallan, V.; Zaidi, A.A. Characterisation of Apple Chlorotic Leaf Spot Virus Infecting Almonds in India. Austral. Plant Dis. Notes 2008, 3, 65. [Google Scholar] [CrossRef]
- Zhang, Y.-Y.; Yan, J.-M.; Zhou, X.-B.; Zhang, Y.-M.; Tao, Y. Effects of N and P Additions on Twig Traits of Wild Apple (Malus sieversii) Saplings. BMC Plant Biol. 2023, 23, 257. [Google Scholar] [CrossRef]
- Nemeth, C.J. Differential Contributions of Majority and Minority Influence. Psychol. Rev. 1986, 93, 23–32. [Google Scholar] [CrossRef]
- Pedrelli, A.; Nali, C.; Panattoni, A.; Pellegrini, E.; Cotrozzi, L. Apple Chlorotic Leaf Spot Virus Infection Affects the Physiology and Biochemistry of Leaves and the Quality of Fruits of Tuscany (Italy) Autochthonous Apple Varieties. Plant Pathol. 2025, 74, 158–170. [Google Scholar] [CrossRef]
- Wright, A.A.; Cross, A.R.; Harper, S.J. A Bushel of Viruses: Identification of Seventeen Novel Putative Viruses by RNA-Seq in Six Apple Trees. PLoS ONE 2020, 15, e0227669. [Google Scholar] [CrossRef]
- Descriptor List for Apple (MALUS); International Board for Plant Genetic Resources: Rome, Italy, 1982; ISBN 92-9043-101-6.
- International Union for the Protection of New Varieties of Plants. Apple (Fruit Varieties): Guidelines For The Conduct Of Tests For Distinctness, Uniformity And Stability. Available online: https://www.upov.int/edocs/tgdocs/en/tg014.pdf (accessed on 2 October 2024).
- Ma, L.; Zeng, Q.; Huang, W.; Wang, S.; Zhang, Y.; Cheng, Y.; Zhang, Q.; Wang, S.; Hao, L.; Xu, C.; et al. Incidence of Major Pome Fruit Tree Viruses and Viroids in Commercial Pear Orchards in China and in Pyrus betulifolia Seedlings. Plant Pathol. 2021, 70, 1467–1475. [Google Scholar] [CrossRef]
- Nabi, S.U.; Mir, J.I.; Sharma, O.C.; Singh, D.B.; Zaffer, S.; Sheikh, M.A.; Masoodi, L.; Khan, K.A. Optimization of Tissue and Time for Rapid Serological and Molecular Detection of Apple Stem Pitting Virus and Apple Stem Grooving Virus in Apple’. Phytoparasitica 2018, 46, 705–713. [Google Scholar] [CrossRef]
- Nabi, S.U.; Mir, J.I.; Yasmin, S.; Din, A.; Raja, W.H.; Madhu, G.S.; Parveen, S.; Mansoor, S.; Chung, Y.S.; Sharma, O.C.; et al. Tissue and Time Optimization for Real-Time Detection of Apple Mosaic Virus and Apple Necrotic Mosaic Virus Associated with Mosaic Disease of Apple (Malus domestica). Viruses 2023, 15, 795. [Google Scholar] [CrossRef] [PubMed]
- Lat Lon Tools. QGIS Plugins. Available online: https://plugins.qgis.org/plugins/latlontools/#plugin-versions (accessed on 1 February 2025).
- QGIS Association. QGIS Geographic Information System. Available online: http://www.qgis.org (accessed on 1 January 2024).
- Kalogirou, S. lctools: Local Correlation, Spatial Inequalities, Geographically Weighted Regression and Other Tools. R Package Version 0.2-10. 2024. Available online: https://CRAN.R-project.org/package=lctools (accessed on 20 June 2025).
- MacHardy, W.E. Apple Scab: Biology, Epidemiology, and Management; American Phytopathological Society (APS Press): St. Paul, MN, USA, 1996; ISBN 978-0-89054-206-4. [Google Scholar]
- Crusius, L.U.; Forcelini, C.A.; Sanhueza, R.M.V.; Fernandes, J.M.C. Epidemiology of Apple Leaf Spot. Fitopatol. Bras. 2002, 27, 65–70. [Google Scholar] [CrossRef]
- Simmons, E.G. Alternaria Themes and Variations (73). Mycotaxon 1993, 48, 109–140. [Google Scholar]
- Van der Zwet, T.; Keil, H.L. Fire Blight: A Bacterial Disease of Rosaceous Plants; United States Department of Agriculture: Washington, DC, USA, 1979. [Google Scholar]
- Fuchs, M. Virus transmission and grafting practices. N. Y. Fruit Q. 2016, 24, 25–27. [Google Scholar]
- Li, L.; Wen, L.; Wang, G.; Lyu, Y.; Yang, Z.; Yang, X.; Li, Q.; Hong, N. Seed Transmission of Three Viruses in Two Pear Rootstock Species Pyrus betulifolia and P. calleryana. Viruses 2022, 14, 599. [Google Scholar] [CrossRef]
- Yoshikawa, N.; Matsuda, H.; Oda, Y.; Isogai, M.; Takahashi, T.; Ito, T.; Yoshida, K. Genome Heterogeneity Of Apple Stem Pitting Virus in Apple Trees. Acta Hortic. 2001, 550, 285–290. [Google Scholar] [CrossRef]
- Wunsch, A.; Hoff, B.; Sazo, M.M.; Van Zoeren, J.; Lamour, K.H.; Hurtado-Gonzales, O.P.; Fuchs, M. Viruses of Apple Are Seedborne but Likely Not Vertically Transmitted. Viruses 2024, 16, 95. [Google Scholar] [CrossRef]
- Escalante, C.; Sanz-Saez, A.; Jacobson, A.; Otulak-Kozieł, K.; Kozieł, E.; Balkcom, K.S.; Zhao, C.; Conner, K. Plant Virus Transmission during Seed Development and Implications to Plant Defense System. Front. Plant Sci. 2024, 15, 1385456. [Google Scholar] [CrossRef]
- Canales, C.; Morán, F.; Olmos, A.; Ruiz-García, A.B. First Detection and Molecular Characterization of Apple Stem Grooving Virus, Apple Chlorotic Leaf Spot Virus, and Apple Hammerhead Viroid in Loquat in Spain. Plants 2021, 10, 2293. [Google Scholar] [CrossRef]
- Eastwell, K.C.; Petrzik, K. Instant Insights: Viruses Affecting Horticultural Crops; Burleigh Dodds Science Publishing: Cambridge, UK, 2024; Volume 106. [Google Scholar]
- Naqvi, S.A.M.H. Diseases of Fruits and Vegetables; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; ISBN 1-4020-1822-3. [Google Scholar]
- Manzoor, S.; Nabi, S.U.; Ayaz, A.; Rasool, B.; Sharma, S.K.; Chesti, M.H.; Parveen, S.; Verma, M.K.; Diab, M.A.; Rabbee, M.F. Unveiling Viral Threats to Temperate Pome Fruits: Characterization, Transmission, and Sustainable Management Strategies. Front. Microbiol. 2025, 16, 1560720. [Google Scholar] [CrossRef]
- Nabi, S.U.; Baranwal, V.K.; Rao, G.P.; Mansoor, S.; Vladulescu, C.; Raja, W.H.; Jan, B.L.; Alansi, S. High-Throughput RNA Sequencing of Mosaic Infected and Non-Infected Apple (Malus × Domestica Borkh.) Cultivars: From Detection to the Reconstruction of Whole Genome of Viruses and Viroid. Plants 2022, 11, 675. [Google Scholar] [CrossRef] [PubMed]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The Global Burden of Pathogens and Pests on Major Food Crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Nizamani, M.M.; Zhang, Q.; Muhae-Ud-Din, G.; Awais, M.; Qayyum, M.; Farhan, M.; Jabran, M.; Wang, Y. Application of GIS and Remote-Sensing Technology in Ecosystem Services and Biodiversity Conservation. In Deep Learning for Multimedia Processing Applications; CRC Press: Boca Raton, FL, USA, 2023; pp. 284–321. ISBN 978-1-032-64626-8. [Google Scholar]
- White, S.M.; Bullock, J.M.; Hooftman, D.A.P.; Chapman, D.S. Modelling the Spread and Control of Xylella Fastidiosa in the Early Stages of Invasion in Apulia, Italy. Biol. Invasions 2017, 19, 1825–1837. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Coordinates | Number of Samples | Distance (km) Between Orchards and Wild Apple Populations | |
|---|---|---|---|---|
| Wild apple | Ketpentau (E) | 43.296568333, 79.515078333 | 50 | 100–1 * |
| 670–2 * | ||||
| 650–3 * | ||||
| Sumbe (D) | 43.289246667, 79.477243333 | 50 | 101–1 * | |
| 665–2 * | ||||
| 650–3 * | ||||
| Tauturgen (B) | 43.364244848, 77.680407392 | 55 | 50–1 * | |
| 520–2 * | ||||
| 500–3 * | ||||
| Sievers Apple Reserve (A) | 43.366810848, 77.673012393 | 70 | 50–1 * | |
| 520–2 * | ||||
| 505–3 * | ||||
| Orchard | Shelek (C) | 43.554083835, 78.280924394 | 14 | 570–2 * |
| 550–3 * | ||||
| Taraz 1 (F) | 42.923489300, 71.498234084 | 20 | 570–1 * | |
| 16–3 * | ||||
| Taraz 2 (G) | 42.868237638, 71.309830750 | 83 | 550–1 * | |
| 16–2 * |
| Primer Name | Sequence (5′-3′) | Source |
|---|---|---|
| ACLSV-F | TGAGAGGCTCTATTCACATCTTG | [36] |
| ACLSV-R | CAATTGGAATATCCCCTTCTGCGAT | |
| ASPV-F | GAGAGAGTAGCCAATGCCACAAGCAA | [36] |
| ASPV-R | CGCCGAAGTTCACAGCCTGAGTACC | |
| ASGV-F | CATATGTTCACTGAGGCAAAAGCTG | [37] |
| ASGV-R | CGATCCAGAAACCCATCAAAGACTT | |
| ApMV-F | ATCCGAGTGAACAGTCTATCCCTC | [38] |
| ApMV-R | GTAACTCACTCGTTATCACGTAC | |
| ApNMV-F | ATGGTGTGCAATCGCTGTCA | [38] |
| ApNMV-R | CATCGACCATAAGGATATCA |
| Population | ACLSV | ASPV | Local Pearson Correlation ACLSV and ASPV | |||
|---|---|---|---|---|---|---|
| Moran’s I | p-Value | Moran’s I | p-Value Resampling | LPCC | p-Value | |
| Ketpentau | 0.05733 | 0.33759 | 0.01667 | 0.64741 | 0.42594 | 0.00204 |
| Sumbe | 0.08766 | 0.17728 | −0.07727 | 0.47776 | 0.18831 | 0.19032 |
| Tauturgen | 0.51212 | 3.59 * 10−12 | 0.21653 | 0.00207 | 0.27501 | 0.04215 |
| Sievers Apple Reserve | −0.00259 | 0.18259 | 0.07938 | 0.15637 | 0.21501 | 0.07603 |
| Taraz | 0.09320 | 0.06714 | 0.09644 | 0.05897 | −0.32938 | 0.00068 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerimbek, N.; Khusnitdinova, M.; Taskuzhina, A.; Kapytina, A.; Pozharskiy, A.; Sagitov, A.; Gritsenko, D. Mapping the Distribution of Viruses in Wild Apple Populations in the Southeast Region of Kazakhstan. Forests 2025, 16, 1119. https://doi.org/10.3390/f16071119
Kerimbek N, Khusnitdinova M, Taskuzhina A, Kapytina A, Pozharskiy A, Sagitov A, Gritsenko D. Mapping the Distribution of Viruses in Wild Apple Populations in the Southeast Region of Kazakhstan. Forests. 2025; 16(7):1119. https://doi.org/10.3390/f16071119
Chicago/Turabian StyleKerimbek, Nazym, Marina Khusnitdinova, Aisha Taskuzhina, Anastasiya Kapytina, Alexandr Pozharskiy, Abay Sagitov, and Dilyara Gritsenko. 2025. "Mapping the Distribution of Viruses in Wild Apple Populations in the Southeast Region of Kazakhstan" Forests 16, no. 7: 1119. https://doi.org/10.3390/f16071119
APA StyleKerimbek, N., Khusnitdinova, M., Taskuzhina, A., Kapytina, A., Pozharskiy, A., Sagitov, A., & Gritsenko, D. (2025). Mapping the Distribution of Viruses in Wild Apple Populations in the Southeast Region of Kazakhstan. Forests, 16(7), 1119. https://doi.org/10.3390/f16071119