
Responses of Water Use Strategies to Seasonal Drought Stress Differed Among Eucalyptus urophylla S.T.Blake × E. grandis Plantations Along with Stand Ages


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site Description
2.2. Experimental Design and Stand Characteristics Measurement
2.3. Sample Collection
- (1)
- Plant xylem samples: On each sampling date, to avoid isotopic enrichment due to stomatal transpiration, healthy non-green and corked branches were selected from the canopy of each standard tree [53]. A branch segment of approximately 0.3–0.5 cm in diameter and 3–5 cm in length were collected on a sunny morning (9:00–12:00), immediately stripped of the phloem, and placed in a brown screw top sampling bottle [52]. The bottle was then sealed with parafilm, quickly placed in a bucket filled with dry ice, and transported to the laboratory and stored in a freezer at −20 °C for freezing before water extraction. A total of 216 plant xylem samples were collected (9 replicates per stand × 4 stands × 6 months).
- (2)
- Leaf samples: On each sampling date (the same as the xylem sampling date), a sample of approximately 20 mature and healthy leaves exposed to full sunlight was collected from the upper canopy of each standard tree on a sunny morning (9:00–12:00) to avoid the canopy effects of leaf δ13C. The leaf samples were placed in a breathable sample bag and returned to the laboratory, where the samples were killed at 105 °C for 20 min, dried at 70 °C for 48 h, then ground at a low temperature in liquid nitrogen, passed through an 80-mesh sieve, sealed, and stored at room temperature. A total of 216 leaf samples were collected (9 replicates per stand × 4 stands × 6 months). The δ13C content of the sampled leaves was measured for long-term WUE analysis.
- (3)
- Soil samples: Soil samples were collected along with the xylem samples on each sampling date. A hole to expose the soil profile was excavated around three standard trees in each plot. Soil samples were collected at depths of 0.1 m, 0.2 m, 0.3 m, 0.4 m, 0.6 m, 0.8 m, 1.0 m, 1.5 m, and 2 m along the soil profile. To avoid the effects of evaporation on the isotopic composition of the soil, approximately 5.0 cm of soil outside of the profile was removed for each layer sampled. Each collected sample was promptly placed into a 12 mL brown sampling bottle with a screw top, sealed with parafilm, immediately placed into a dry ice bucket, and transported to the laboratory [52]. Soil samples were frozen in a freezer at −20 °C prior to moisture extraction. A total of 648 soil samples were collected (9 depth intervals × 3 replicates per stand × 4 stands × 6 months). Meanwhile, the same number of soil samples were collected using ring knives (inner diameter = 5.1 cm, height = 5.0 cm) and sealed bags from each soil layer to determine soil water content (SWC) and physico-chemical properties.
- (4)
- Rainwater samples: All precipitation in the study region falls as rain. We collected rainwater samples using the rainwater sampler recommended by the International Atomic Energy Agency’s Global Network of Isotopes in Precipitation. To minimize the effects of evaporation, rainwater samples were collected immediately after rainfall events. About 2 mL of the sample (or whole sample if <2 mL) was removed from the collection bottle and transferred to a brown sample bottle with a screw cap. The bottle was sealed with parafilm and immediately placed in a portable incubator at 4 °C. The sample was stored in a refrigerator at 4 °C until tested. A total of 31 rainfall events were sampled during the monitoring period.
2.4. Soil Physico-Chemical Properties Analysis
2.5. Meteorological Factors and Soil Water Content Measurement
2.6. Measurement of Stable Isotopic Compositions of Water Samples and Leaf Samples
2.7. Plant Water Source and WUE Identification
2.8. Statistical Analyses
3. Results
3.1. Relative Extractable Water (REW) and Fine Root Distribution
3.2. Shifts in Water Use Sources
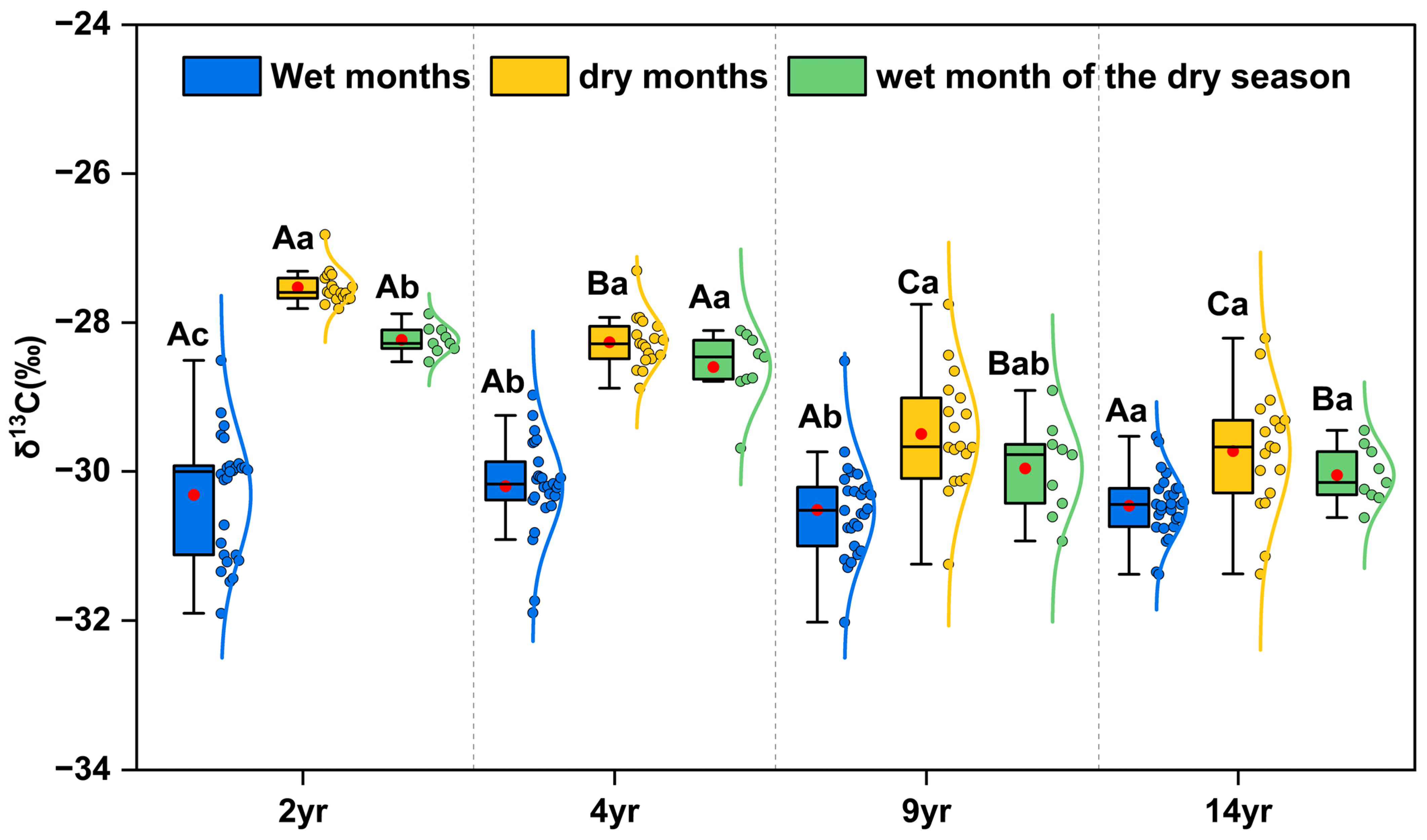
3.3. Leaf δ13C and WUE
3.4. Factors Influencing Water Use Sources and WUE
4. Discussion
4.1. Water Source Used by E. urophylla × E. grandis Plantations at Four Stand Ages
4.2. Seasonal Changes in WUE in E. urophylla × E. grandis Plantations at Four Stand Ages
4.3. Age-Related Water Use Strategies and Implications for Plantation Management
4.4. Limitations and Perspectives for Future Exploration
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicator | Soil Depth (cm) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 0–10 | 10–20 | 20–30 | 30–40 | 40–60 | 60–80 | 80–100 | 100–150 | 150–200 | |
| pH | 5.33 (0.28) | 5.41 (0.27) | 5.45 (0.3) | 5.48 (0.32) | 5.51 (0.29) | 5.56 (0.32) | 5.51 (0.29) | 5.67 (0.27) | 5.71 (0.35) |
| BD (g·m−3) | 0.99 (0.04) | 1.11 (0.03) | 1.13 (0.05) | 1.19 (0.05) | 1.27 (0.03) | 1.26 (0.07) | 1.28 (0.05) | 1.28 (0.08) | 1.27 (0.05) |
| Tpo (%) | 56.94 (2.7) | 56.86 (3.03) | 57.5 (2.11) | 57.9 (1.56) | 55.4 (1.48) | 55.2 (1.75) | 53.2 (3.59) | 52.47 (2.27) | 50.44 (2.93) |
| Cpo (%) | 56.16 (2.31) | 53.44 (2.97) | 55.6 (2.12) | 52 (1.52) | 50.4 (1.30) | 53.1 (1.87) | 50.7 (2.66) | 51.42 (1.94) | 49.26 (2.3) |
| SWHC (%) | 57.46 (5.72) | 59.62 (6.59) | 58.69 (5.04) | 57.87 (3.51) | 55.73 (2.80) | 52.52 (5.01) | 48.36 (7.78) | 43.36 (5.2) | 42.85 (6.49) |
| CWHC (%) | 56.68 (5.00) | 56.08 (6.44) | 55.91 (4.99) | 52.77 (3.52) | 50.7 (2.42) | 50.52 (5.01) | 46.09 (6.15) | 42.49 (4.53) | 41.85 (5.34) |
| FWHC (%) | 56.27 (5.01) | 53.41 (6.60) | 53.42 (5.02) | 52.44 (3.47) | 49.6 (2.37) | 47.57 (4.99) | 45.18 (6.06) | 42.05 (4.47) | 41.48 (5.27) |
| SOM (%) | 2.88 (0.43) | 2.76 (0.66) | 2.13 (0.3) | 1.83 (0.10) | 1.66 (0.32) | 1.09 (0.15) | 1.66 (0.32) | 1.63 (0.26) | 1.65 (0.29) |
| TN (g·kg−1) | 2.51 (0.1) | 2.39 (0.2) | 2.29 (0.88) | 1.95 (0.39) | 1.96 (0.05) | 1.80 (2.05) | 1.96 (0.05) | 1.94 (0.72) | 1.88 (0.38) |
| TP (g·kg−1) | 1.23 (0.24) | 1.21 (0.29) | 1.18 (0.16) | 0.72 (0.1) | 1.03 (0.22) | 0.84 (0.10) | 1.03 (0.22) | 0.95 (0.18) | 0.89 (0.2) |
| TK (g·kg−1) | 2.95 (0.7) | 2.85 (0.93) | 2.92 (0.92) | 3.02 (0.73) | 3.26 (0.59) | 3.24 (1.10) | 3.26 (0.59) | 3.22 (0.76) | 3.25 (0.68) |
| AN (mg·kg−1) | 351.22 (48.13) | 347.74 (41.72) | 292.3 (44.05) | 267.78 (47.92) | 267.31 (51.34) | 267.62 (42.50) | 267.31 (51.33) | 267.43 (48.39) | 266.98 (49.86) |
| AP (mg·kg−1) | 3.4 (2.02) | 3.34 (1.29) | 3.29 (1.32) | 2.79 (1.9) | 3.47 (2.39) | 2.75 (0.76) | 3.47 (2.39) | 2.89 (1.85) | 3.21 (2.12) |
| AK (mg·kg−1) | 31.54 (10.1) | 30.94 (8.57) | 26.81 (12.02) | 24.21 (9.93) | 18.69 (5.86) | 18.44 (6.56) | 18.69 (5.86) | 19.26 (6.09) | 18.77 (5.98) |
| Ages | Layers | Wet Months | Dry Months | Wet Month of the Dry Season | |||
|---|---|---|---|---|---|---|---|
| δ18O (‰) | δD (‰) | δ18O (‰) | δD (‰) | δ18O (‰) | δD (‰) | ||
| 2 yr | Shallow | −8.22 (1.38) a | −57.32 (7.38) a | −7.40 (2.13) a | −55.73 (14.77) a | −6.35 (0.61) a | −50.80 (2.84) a |
| Middle | −10.03 (0.54) b | −67.05 (7.11) b | −10.08 (0.81) b | −70.35 (2.92) b | −8.86 (0.81) b | −63.59 (2.27) b | |
| Deep | −10.58 (0.81) b | −70.20 (4.20) b | −9.62 (1.20) b | −68.82 (7.64) b | −9.34 (0.05) b | −67.54 (0.22) b | |
| 4 yr | Shallow | −9.39 (2.34) a | −64.38 (15.99) a | −6.89 (1.24) a | −51.49 (11.93) a | −7.32 (0.71) a | −50.11 (4.71) a |
| Middle | −10.31 (1.62) ab | −68.10 (7.89) ab | −9.22 (1.34) b | −62.32 (5.04) ab | −9.89 (0.57) b | −64.90 (1.76) b | |
| Deep | −10.75 (0.30) b | −73.72 (2.22) b | −10.8 (0.44) b | −72.42 (1.49) b | −10.43 (0.21) b | −69.20 (3.60) b | |
| 9 yr | Shallow | −7.27 (0.09) a | −56.47 (4.5) a | −9.31 (2.13) a | −63.23 (14.33) a | −7.15 (1.09) a | −55.84 (1.83) a |
| Middle | −9.47 (0.3) b | −76.87 (13.35) b | −11.11 (0.47) b | −73.78 (2.25) b | −10.15 (0.90) b | −68.26 (2.91) b | |
| Deep | −10.96 (0.21) c | −81.79 (0.63) c | −10.09 (0.17) ab | −67.38 (5.09) ab | −10.48 (0.74) b | −67.22 (2.09) b | |
| 14 yr | Shallow | −8.55 (0.75) a | −58.92 (3.24) a | −8.96 (1.10) a | −64.53 (7.74) a | −6.56 (0.17) a | −51.47 (1.97) a |
| Middle | −11.68 (0.66) b | −78.51 (5.58) b | −10.42 (1.00) ab | −69.99 (5.43) ab | −9.20 (1.16) b | −65.94 (1.00) b | |
| Deep | −12.07 (0.36) b | −80.58 (6.03) b | −11.03 (0.74) b | −73.91 (3.28) b | −9.91 (0.69) b | −67.50 (2.51) b | |
Appendix B


References
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Zhou, L.; Foltz, G.R.; Qu, X.; Ying, J.; Tokinaga, H.; Mechoso, C.R.; Li, J.; Gu, X. Hydrological cycle changes under global warming and their effects on multiscale climate variability. Ann. N. Y. Acad. Sci. 2020, 1472, 21–48. [Google Scholar] [CrossRef]
- Wang, X.; Li, Y.; Wang, M.; Li, Y.; Gong, X.; Chen, Y.; Chen, Y.; Cao, W. Changes in daily extreme temperature and precipitation events in mainland China from 1960 to 2016 under global warming. Int. J. Climatol. 2021, 41, 1465–1483. [Google Scholar] [CrossRef]
- Yaduvanshi, A.; Nkemelang, T.; Bendapudi, R.; New, M. Temperature and rainfall extremes change under current and future global warming levels across Indian climate zones. Weather Clim. Extrem. 2021, 31, 100291. [Google Scholar] [CrossRef]
- Han, L.; Zhang, Q.; Zhang, Z.; Jia, J.; Wang, Y.; Huang, T.; Cheng, Y. Drought area, intensity and frequency changes in China under climate warming, 1961–2014. J. Arid Environ. 2021, 193, 104596. [Google Scholar] [CrossRef]
- Dai, Y.; Wang, H.-W.; Shi, Q.-D. Contrasting plant water-use responses to groundwater depth from seedlings to mature trees in the Gurbantunggut Desert. J. Hydrol. 2022, 610, 127986. [Google Scholar] [CrossRef]
- Huang, M.; Zhai, P.; Piao, S. Divergent responses of ecosystem water use efficiency to drought timing over Northern Eurasia. Environ. Res. Lett. 2021, 16, 045016. [Google Scholar] [CrossRef]
- Wang, J.; Fu, B.; Jiao, L.; Lu, N.; Li, J.; Chen, W.; Wang, L. Age-related water use characteristics of Robinia pseudoacacia on the Loess Plateau. Agric. For. Meteorol. 2021, 301–302, 108344. [Google Scholar] [CrossRef]
- Wang, M.; Ding, Z.; Wu, C.; Song, L.; Ma, M.; Yu, P.; Lu, B.; Tang, X. Divergent responses of ecosystem water-use efficiency to extreme seasonal droughts in Southwest China. Sci. Total Environ. 2021, 760, 143427. [Google Scholar] [CrossRef]
- Dai, J.; Zhao, Y.; Seki, K.; Wang, L. Changes in water-use strategies and soil water status of degraded poplar plantations in water-limited areas. Agric. Water Manag. 2024, 296, 108799. [Google Scholar] [CrossRef]
- Yin, D.; Gou, X.; Liu, J.; Zhang, D.; Wang, K.; Yang, H. Increasing deep soil water uptake during drought does not indicate higher drought resistance. J. Hydrol. 2024, 630, 130694. [Google Scholar] [CrossRef]
- Liu, Z.; Yu, X.; Jia, G. Water uptake by coniferous and broad-leaved forest in a rocky mountainous area of northern China. Agric. For. Meteorol. 2019, 265, 381–389. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, L. Plant water use strategy in response to spatial and temporal variation in precipitation patterns in China: A stable isotope analysis. Forests 2018, 9, 123. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, Q.; Wei, Z.; Yu, X.; Jia, G.; Jiang, J. Partitioning tree water usage into storage and transpiration in a mixed forest. For. Ecosyst. 2021, 8, 72. [Google Scholar] [CrossRef]
- Tetzlaff, D.; Buttle, J.; Carey, S.K.; Kohn, M.J.; Laudon, H.; McNamara, J.P.; Smith, A.; Sprenger, M.; Soulsby, C. Stable isotopes of water reveal differences in plant–soil water relationships across northern environments. Hydrol. Process. 2021, 35, e14023. [Google Scholar] [CrossRef]
- Pei, Y.; Huang, L.; Jia, X.; Tang, X.; Zhang, Y.; Pan, Y. Water sources used by artificial Salix psammophila in stands of different ages based on stable isotope analysis in northeastern Mu Us Sandy Land. Catena 2023, 226, 107087. [Google Scholar] [CrossRef]
- Wang, J.; Fu, B.; Lu, N.; Zhang, L. Seasonal variation in water uptake patterns of three plant species based on stable isotopes in the semi-arid Loess Plateau. Sci. Total Environ. 2017, 609, 27–37. [Google Scholar] [CrossRef]
- Chang, E.; Li, P.; Li, Z.; Xiao, L.; Zhao, B.; Su, Y.; Feng, Z. Using water isotopes to analyze water uptake during vegetation succession on abandoned cropland on the Loess Plateau, China. Catena 2019, 181, 104095. [Google Scholar] [CrossRef]
- Ward, D.; Wiegand, K.; Getzin, S. Walter’s two-layer hypothesis revisited: Back roots! Oecologia 2013, 172, 617–630. [Google Scholar] [CrossRef]
- Xu, H.; Li, Y. Water-use strategy of three central Asian desert shrubs and their responses to rain pulse events. Plant Soil 2006, 285, 5–17. [Google Scholar] [CrossRef]
- Xu, Q.; Li, H.; Chen, J.; Cheng, X.; Liu, S.; An, S. Water use patterns of three species in subalpine forest, Southwest China: The deuterium isotope approach. Ecohydrology 2011, 4, 236–244. [Google Scholar] [CrossRef]
- Magh, R.-K.; Eiferle, C.; Burzlaff, T.; Dannenmann, M.; Rennenberg, H.; Dubbert, M. Competition for water rather than facilitation in mixed beech-fir forests after drying-wetting cycle. J. Hydrol. 2020, 587, 124944. [Google Scholar] [CrossRef]
- Tiemuerbieke, B.; Min, X.-J.; Zang, Y.-X.; Xing, P.; Ma, J.-Y.; Sun, W. Water use patterns of co-occurring C3 and C4 shrubs in the Gurbantonggut desert in northwestern China. Sci. Total Environ. 2018, 634, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Kray, J.A.; Cooper, D.J.; Sanderson, J.S. Groundwater use by native plants in response to changes in precipitation in an intermountain basin. J. Arid Environ. 2012, 83, 25–34. [Google Scholar] [CrossRef]
- Gao, Y.; He, L.; Jia, Z.; Li, Q.; Dai, J. Effects of precipitation on water use characteristics of Caragana intermedia plantations with different stand ages in alpine sandy land. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2021, 32, 1935–1942. [Google Scholar] [CrossRef]
- Song, L.; Zhu, J.; Li, M.; Zhang, J.; Li, D. Water use strategies of natural Pinus sylvestris var. Mongolica trees of different ages in Hulunbuir Sandy Land of Inner Mongolia, China, based on stable isotope analysis. Trees 2018, 32, 1001–1011. [Google Scholar] [CrossRef]
- Goldsmith, G.R.; Muñoz-Villers, L.E.; Holwerda, F.; McDonnell, J.J.; Asbjornsen, H.; Dawson, T.E. Stable isotopes reveal linkages among ecohydrological processes in a seasonally dry tropical montane cloud forest. Ecohydrology 2012, 5, 779–790. [Google Scholar] [CrossRef]
- Mininni, A.N.; Tuzio, A.C.; Brugnoli, E.; Dichio, B.; Sofo, A. Carbon isotope discrimination and water use efficiency in interspecific Prunus hybrids subjected to drought stress. Plant Physiol. Biochem. 2022, 175, 33–43. [Google Scholar] [CrossRef]
- Mokhtar, A.; He, H.; Alsafadi, K.; Mohammed, S.; He, W.; Li, Y.; Zhao, H.; Abdullahi, N.M.; Gyasi-Agyei, Y. Ecosystem water use efficiency response to drought over southwest China. Ecohydrology 2021, 15, e2317. [Google Scholar] [CrossRef]
- Sun, S.; Xiang, W.; Ouyang, S.; Hu, Y.; Peng, C. Balancing water yield and water use efficiency between planted and natural forests: A global analysis. Glob. Change Biol. 2024, 30, e17561. [Google Scholar] [CrossRef]
- Gong, X.Y.; Ma, W.T.; Yu, Y.Z.; Fang, K.; Yang, Y.; Tcherkez, G.; Adams, M.A. Overestimated gains in water-use efficiency by global forests. Glob. Change Biol. 2022, 28, 4923–4934. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ma, F.-Y.; Hu, T.-X.; Zhao, K.-G.; Gao, T.-P.; Zhao, H.-X.; Ning, T.-Y. Using stable isotopes to quantify water uptake from different soil layers and water use efficiency of wheat under long-term tillage and straw return practices. Agric. Water Manag. 2020, 229, 105933. [Google Scholar] [CrossRef]
- Hai, X.; Li, J.; Li, J.; Liu, Y.; Dong, L.; Wang, X.; Lv, W.; Hu, Z.; Shangguan, Z.; Deng, L. Variations in plant water use efficiency response to manipulated precipitation in a temperate grassland. Front. Plant Sci. 2022, 13, 881282. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Jia, X.; Zha, T.; Bourque, C.P.-A.; Tian, Y.; Bai, Y.; Liu, P.; Yang, R.; Li, C.; Li, C.; et al. Ecosystem water use efficiency in a young plantation in Northern China and its relationship to drought. Agric. For. Meteorol. 2019, 275, 1–10. [Google Scholar] [CrossRef]
- Song, L.; Zhu, J.; Zhang, J.; Zhang, T.; Wang, K.; Wang, G.; Liu, J. Effect of Drought and Topographic Position on Depth of Soil Water Extraction of Pinus sylvestris L. var. mongolica Litv. Trees in a Semiarid Sandy Region, Northeast China. Forests 2019, 10, 370. [Google Scholar] [CrossRef]
- Cao, M.; Wu, C.; Liu, J.; Jiang, Y. Increasing leaf δ13C values of woody plants in response to water stress induced by tunnel excavation in a karst trough valley: Implication for improving water-use efficiency. J. Hydrol. 2020, 586, 124895. [Google Scholar] [CrossRef]
- Vickers, D.; Thomas, C.K.; Pettijohn, C.; Martin, J.G.; Law, B. Five years of carbon fluxes and inherent water-use efficiency at two semi-arid pine forests with different disturbance histories. Tellus B Chem. Phys. Meteorol. 2012, 64, 17159. [Google Scholar] [CrossRef]
- Reichstein, M.; Tenhunen, J.D.; Roupsard, O.; Ourcival, J.-M.; Rambal, S.; Miglietta, F.; Peressotti, A.; Pecchiari, M.; Tirone, G.; Valentini, R. Severe drought effects on ecosystem CO2 and H2O fluxes at three Mediterranean evergreen sites: Revision of current hypotheses? Glob. Change Biol. 2002, 8, 999–1017. [Google Scholar] [CrossRef]
- Schulze, E.-D.; Mooney, H.A.; Sala, O.; Jobbagy, E.; Buchmann, N.; Bauer, G.; Canadell, J.; Jackson, R.; Loreti, J.; Oesterheld, M. Rooting depth, water availability, and vegetation cover along an aridity gradient in Patagonia. Oecologia 1996, 108, 503–511. [Google Scholar] [CrossRef]
- Yanghua, Y.; Wei, Z.; Xinping, Z.; Bin, Y. Stoichiometric characteristics in Zanthoxylum planispinum var. dintanensis plantation of different ages. Agron. J. 2021, 113, 685–695. [Google Scholar] [CrossRef]
- Jassal, R.S.; Black, T.A.; Spittlehouse, D.L.; Brümmer, C.; Nesic, Z. Evapotranspiration and water use efficiency in different-aged Pacific Northwest Douglas-fir stands. Agric. For. Meteorol. 2009, 149, 1168–1178. [Google Scholar] [CrossRef]
- Skubel, R.; Arain, M.A.; Peichl, M.; Brodeur, J.J.; Khomik, M.; Thorne, R.; Trant, J.; Kula, M. Age effects on the water-use efficiency and water-use dynamics of temperate pine plantation forests. Hydrol. Process. 2015, 29, 4100–4113. [Google Scholar] [CrossRef]
- Liu, L.; Jia, Z.; Zhu, Y.; Li, H.; Yang, D.; Wei, D.; Zhao, X. Water Use Strategy of Salix cheilophila Stands with Different Ages in Gonghe Basin, Qinghai Province. For. Res. 2012, 25, 597–603. [Google Scholar] [CrossRef]
- Heilman, K.A.; Trouet, V.M.; Belmecheri, S.; Pederson, N.; Berke, M.A.; McLachlan, J.S. Increased water use efficiency leads to decreased precipitation sensitivity of tree growth, but is offset by high temperatures. Oecologia 2021, 197, 1095–1110. [Google Scholar] [CrossRef]
- Arnold, R.; Xie, Y.; Luo, J.; Wang, H.; Midgley, S. A tale of two genera: Exotic Eucalyptus and Acacia species in China. 1. Domestication and research. Int. For. Rev. 2020, 22, 1–18. [Google Scholar] [CrossRef]
- Campoe, O.C.; Alvares, C.A.; Carneiro, R.L.; Binkley, D.; Ryan, M.G.; Hubbard, R.M.; Stahl, J.; Moreira, G.; Moraes, L.F.; Stape, J.L. Climate and genotype influences on carbon fluxes and partitioning in Eucalyptus plantations. For. Ecol. Manag. 2020, 475, 118445. [Google Scholar] [CrossRef]
- Wang, Z.; Du, A.; Xu, Y.; Zhu, W.; Zhang, J. Factors Limiting the Growth of Eucalyptus and the Characteristics of Growth and Water Use under Water and Fertilizer Management in the Dry Season of Leizhou Peninsula, China. Agronomy 2019, 9, 590. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, P.; Ouyang, L.; Zhu, L.; Ni, G.; Schäfer, K.V. Whole-plant water hydraulic integrity to predict drought-induced Eucalyptus urophylla mortality under drought stress. For. Ecol. Manag. 2020, 468, 118179. [Google Scholar] [CrossRef]
- Hechter, U.; Little, K.; Chan, J.; Crous, J.; da Costa, D. Factors affecting eucalypt survival in South African plantation forestry. South. For. A J. For. Sci. 2022, 84, 253–270. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World reference base for soil resource 2006. In World Soil Resources Reports No. 103, 2nd ed.; FAO: Rome, Italy, 2006. [Google Scholar]
- Xu, Y.; Du, A.; Wang, Z.; Zhu, W.; Li, C.; Wu, L. Effects of different rotation periods of Eucalyptus plantations on soil physiochemical properties, enzyme activities, microbial biomass and microbial community structure and diversity. For. Ecol. Manag. 2020, 456, 117683. [Google Scholar] [CrossRef]
- Pei, Y.; Huang, L.; Zhang, Y.; Pan, Y. Water use pattern and transpiration of Mongolian pine plantations in relation to stand age on northern Loess Plateau of China. Agric. For. Meteorol. 2023, 330, 109320. [Google Scholar] [CrossRef]
- Ding, Y.; Nie, Y.; Chen, H.; Wang, K.; Querejeta, J.I. Water uptake depth is coordinated with leaf water potential, water-use efficiency and drought vulnerability in karst vegetation. New Phytol. 2021, 229, 1339–1353. [Google Scholar] [CrossRef]
- Wang, D.; Chen, J.; Tang, Z.; Zhang, Y. Effects of Soil Physical Properties on Soil Infiltration in Forest Ecosystems of Southeast China. Forests 2024, 15, 1470. [Google Scholar] [CrossRef]
- Walkley, A. An examination of methods for determining organic carbon and nitrogen in Soils1. (With one text-figure.). J. Agric. Sci. 1935, 25, 598–609. [Google Scholar] [CrossRef]
- Wu, L.; Li, Z.; Li, J.; Khan, M.A.; Huang, W.; Zhang, Z.; Lin, W. Assessment of shifts in microbial community structure and catabolic diversity in response to Rehmannia glutinosa monoculture. Appl. Soil Ecol. 2013, 67, 1–9. [Google Scholar] [CrossRef]
- Rukun, L. Analytical Methods for Soil and Agricultural Chemistry; Chinese Agriculture Science and Technology Press: Beijing, China, 1999; pp. 159–160. [Google Scholar]
- Liu, H.; Wang, Y.; Tang, M. Arbuscular mycorrhizal fungi diversity associated with two halophytes Lycium barbarum L. and Elaeagnus angustifolia L. in Ningxia, China. Arch. Agron. Soil Sci. 2017, 63, 796–806. [Google Scholar] [CrossRef]
- Tu, J.; Wang, B.; McGrouther, K.; Wang, H.; Ma, T.; Qiao, J.; Wu, L. Soil quality assessment under different Paulownia fortunei plantations in mid-subtropical China. J. Soils Sediments 2017, 17, 2371–2382. [Google Scholar] [CrossRef]
- Granier, A. Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements. Tree Physiol. 1987, 3, 309–320. [Google Scholar] [CrossRef]
- Gao, X.; Sun, S.; Meng, P.; Cai, J.; Pei, S.; Huang, H.; Zhang, J. Carbon fluxes and water-use efficiency in a Pinus tabuliformis plantation in Northeast China and their relationship to drought. Sci. Total Environ. 2024, 946, 174258. [Google Scholar] [CrossRef]
- Jia, G.; Liu, Z.; Chen, L.; Yu, X. Distinguish water utilization strategies of trees growing on earth-rocky mountainous area with transpiration and water isotopes. Ecol. Evol. 2017, 7, 10640–10651. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, L. Insights into the isotopic mismatch between bulk soil water and Salix matsudana Koidz trunk water from root water stable isotope measurements. Hydrol. Earth Syst. Sci. 2021, 25, 3975–3989. [Google Scholar] [CrossRef]
- Gupta, P.; Noone, D.; Galewsky, J.; Sweeney, C.; Vaughn, B.H. Demonstration of high-precision continuous measurements of water vapor isotopologues in laboratory and remote field deployments using wavelength-scanned cavity ring-down spectroscopy (WS-CRDS) technology. Rapid Commun. Mass Spectrom. Int. J. Devoted Rapid Dissem. Up-to-the-Minute Res. Mass Spectrom. 2009, 23, 2534–2542. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, X.; Huang, Z.; Bao, Y.; Jiang, J.; Liu, Z. Quercus acutissima exhibits more adaptable water uptake patterns in response to seasonal changes compared to Pinus massoniana. For. Ecosyst. 2025, 12, 100255. [Google Scholar] [CrossRef]
- Parnell, A.C.; Phillips, D.L.; Bearhop, S.; Semmens, B.X.; Ward, E.J.; Moore, J.W.; Jackson, A.L.; Grey, J.; Kelly, D.J.; Inger, R. Bayesian stable isotope mixing models. Environmetrics 2013, 24, 387–399. [Google Scholar] [CrossRef]
- Ding, Y.L.; Chen, H.S.; Nie, Y.P.; Wang, S.; Zhang, H.L.; Wang, K.L. Water use strategy of Eucalyptus urophylla × E. grandis on karst hillslope based on isotope analysis. J. Appl. Ecol. 2016, 27, 2729–2736. [Google Scholar] [CrossRef]
- Ward, E.J.; Semmens, B.X.; Schindler, D.E. Including source uncertainty and prior information in the analysis of stable isotope mixing models. Environ. Sci. Technol. 2010, 44, 4645–4650. [Google Scholar] [CrossRef]
- Hector, A.; Bagchi, R. Biodiversity and ecosystem multifunctionality. Nature 2007, 448, 188–190. [Google Scholar] [CrossRef]
- Isbell, F.; Calcagno, V.; Hector, A.; Connolly, J.; Harpole, W.S.; Reich, P.B.; Scherer-Lorenzen, M.; Schmid, B.; Tilman, D.; Van Ruijven, J. High plant diversity is needed to maintain ecosystem services. Nature 2011, 477, 199–202. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Huo, G.; Zhao, X.; Gao, X.; Wang, S.; Pan, Y. Seasonal water use patterns of rainfed jujube trees in stands of different ages under semiarid Plantations in China. Agric. Ecosyst. Environ. 2018, 265, 392–401. [Google Scholar] [CrossRef]
- Yang, Q.; Fan, J.; Xing, Y.; Tong, B.; Luo, Z. Water use strategies for three dominant tree species in pure plantations of the semi-arid Chinese Loess Plateau. J. Hydrol. 2025, 654, 132844. [Google Scholar] [CrossRef]
- Dawson, T.E.; Pate, J.S. Seasonal water uptake and movement in root systems of Australian phraeatophytic plants of dimorphic root morphology: A stable isotope investigation. Oecologia 1996, 107, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Nie, Y.; Wang, K. Spatio-temporal heterogeneity of water and plant adaptation mechanisms in karst regions: A review. Shengtai Xuebao Acta Ecol. Sin. 2013, 33, 317–326. [Google Scholar] [CrossRef]
- Schenk, H.J.; Jackson, R.B. The global biogeography of roots. Ecol. Monogr. 2002, 72, 311–328. [Google Scholar] [CrossRef]
- Amin, A.; Zuecco, G.; Geris, J.; Schwendenmann, L.; McDonnell, J.J.; Borga, M.; Penna, D. Depth distribution of soil water sourced by plants at the global scale: A new direct inference approach. Ecohydrology 2020, 13, e2177. [Google Scholar] [CrossRef]
- Wu, J.; Zeng, H.; Zhao, F.; Chen, C.; Singh, A.K.; Jiang, X.; Yang, B.; Liu, W. Plant hydrological niches become narrow but stable as the complexity of interspecific competition increases. Agric. For. Meteorol. 2022, 320, 108953. [Google Scholar] [CrossRef]
- February, E.C.; Higgins, S.I. The distribution of tree and grass roots in savannas in relation to soil nitrogen and water. S. Afr. J. Bot. 2010, 76, 517–523. [Google Scholar] [CrossRef]
- Fitter, A. Characteristics and functions of root systems. In Plant Roots; CRC Press: Boca Raton, FL, USA, 2002; pp. 49–78. [Google Scholar]
- Oliveira, R.; Bezerra, L.; Davidson, E.; Pinto, F.; Klink, C.; Nepstad, D.; Moreira, A. Deep root function in soil water dynamics in cerrado savannas of central Brazil. Funct. Ecol. 2005, 19, 574–581. [Google Scholar] [CrossRef]
- Schwinning, S.; Davis, K.; Richardson, L.; Ehleringer, J.R. Deuterium enriched irrigation indicates different forms of rain use in shrub/grass species of the Colorado Plateau. Oecologia 2002, 130, 345–355. [Google Scholar] [CrossRef]
- Schwinning, S.; Starr, B.I.; Ehleringer, J.R. Summer and winter drought in a cold desert ecosystem (Colorado Plateau) part I: Effects on soil water and plant water uptake. J. Arid Environ. 2005, 60, 547–566. [Google Scholar] [CrossRef]
- Kwon, H.; Law, B.E.; Thomas, C.K.; Johnson, B.G. The influence of hydrological variability on inherent water use efficiency in forests of contrasting composition, age, and precipitation regimes in the Pacific Northwest. Agric. For. Meteorol. 2018, 249, 488–500. [Google Scholar] [CrossRef]
- Pirasteh-Anosheh, H.; Saed-Moucheshi, A.; Pakniyat, H.; Pessarakli, M. Stomatal Responses to Drought Stress; John Wiley & Sons: Hoboken, NJ, USA, 2016; Volume 1, pp. 24–40. [Google Scholar]
- Wu, J.; Serbin, S.P.; Ely, K.S.; Wolfe, B.T.; Dickman, L.T.; Grossiord, C.; Michaletz, S.T.; Collins, A.D.; Detto, M.; McDowell, N.G. The response of stomatal conductance to seasonal drought in tropical forests. Glob. Change Biol. 2020, 26, 823–839. [Google Scholar] [CrossRef]
- Landsberg, J.; Waring, R. Water relations in tree physiology: Where to from here? Tree Physiol. 2017, 37, 18–32. [Google Scholar] [CrossRef]
- Law, B.; Williams, M.; Anthoni, P.; Baldocchi, D.; Unsworth, M. Measuring and modelling seasonal variation of carbon dioxide and water vapour exchange of a Pinus ponderosa forest subject to soil water deficit. Glob. Change Biol. 2000, 6, 613–630. [Google Scholar] [CrossRef]
- Martorell, S.; Diaz-Espejo, A.; Medrano, H.; Ball, M.C.; Choat, B. Rapid hydraulic recovery in Eucalyptus pauciflora after drought: Linkages between stem hydraulics and leaf gas exchange. Plant Cell Environ. 2014, 37, 617–626. [Google Scholar] [CrossRef]
- Brodersen, C.R.; McElrone, A.J.; Choat, B.; Matthews, M.A.; Shackel, K.A. The dynamics of embolism repair in xylem: In vivo visualizations using high-resolution computed tomography. Plant Physiol. 2010, 154, 1088–1095. [Google Scholar] [CrossRef]
- Salleo, S.; Trifilò, P.; Esposito, S.; Nardini, A.; Gullo, M.A.L. Starch-to-sugar conversion in wood parenchyma of field-growing Laurus nobilis plants: A component of the signal pathway for embolism repair? Funct. Plant Biol. 2009, 36, 815–825. [Google Scholar] [CrossRef]
- Alsina, M.; De Herralde, F.; Aranda, X.; Save, R.; Biel, C. Water relations and vulnerability to embolism are not related: Experiments with eight grapevine cultivars. Vitis Geilweilerhof 2007, 46, 1–7. [Google Scholar] [CrossRef]
- Pockman, W.T.; Sperry, J.S. Vulnerability to xylem cavitation and the distribution of Sonoran desert vegetation. Am. J. Bot. 2000, 87, 1287–1299. [Google Scholar] [CrossRef]
- Mukarram, M.; Choudhary, S.; Kurjak, D.; Petek, A.; Khan, M.M.A. Drought: Sensing, signalling, effects and tolerance in higher plants. Physiol. Plant. 2021, 172, 1291–1300. [Google Scholar] [CrossRef]
- Germon, A.; Cardinael, R.; Prieto, I.; Mao, Z.; Kim, J.; Stokes, A.; Dupraz, C.; Laclau, J.-P.; Jourdan, C. Unexpected phenology and lifespan of shallow and deep fine roots of walnut trees grown in a silvoarable Mediterranean agroforestry system. Plant Soil 2016, 401, 409–426. [Google Scholar] [CrossRef]
- Yu, L.; Gao, X.; Zhao, X. Global synthesis of the impact of droughts on crops’ water-use efficiency (WUE): Towards both high WUE and productivity. Agric. Syst. 2020, 177, 102723. [Google Scholar] [CrossRef]
- Di, N.; Yang, S.; Liu, Y.; Fan, Y.; Duan, J.; Nadezhdina, N.; Li, X.; Xi, B. Soil-moisture-dependent nocturnal water use strategy and its responses to meteorological factors in a seasonal-arid poplar plantation. Agric. Water Manag. 2022, 274, 107984. [Google Scholar] [CrossRef]
- Pu, X.; Lyu, L. Disentangling the impact of photosynthesis and stomatal conductance on rising water-use efficiency at different altitudes on the Tibetan plateau. Agric. For. Meteorol. 2023, 341, 109659. [Google Scholar] [CrossRef]
- Zhao, J.; Xu, T.; Xiao, J.; Liu, S.; Mao, K.; Song, L.; Yao, Y.; He, X.; Feng, H. Responses of water use efficiency to drought in southwest China. Remote Sens. 2020, 12, 199. [Google Scholar] [CrossRef]
- James, J.; Page-Dumroese, D.; Busse, M.; Palik, B.; Zhang, J.; Eaton, B.; Slesak, R.; Tirocke, J.; Kwon, H. Effects of forest harvesting and biomass removal on soil carbon and nitrogen: Two complementary meta-analyses. For. Ecol. Manag. 2021, 485, 118935. [Google Scholar] [CrossRef]
- Špulák, O.; Kacálek, D. How different approaches to logging residues handling affected retention of nutrients at poor-soil Scots pine site after clear-cutting? A case study. J. For. Sci. 2020, 66, 461–470. [Google Scholar] [CrossRef]
- Nan, W.; Ta, F.; Meng, X.; Dong, Z.; Xiao, N. Effects of age and density of Pinus sylvestris var. mongolica on soil moisture in the semiarid Mu Us Dunefield, northern China. For. Ecol. Manag. 2020, 473, 118313. [Google Scholar] [CrossRef]
- Gendvilas, V.; Neyland, M.; Rocha-Sepúlveda, M.F.; Downes, G.M.; Hunt, M.; Jacobs, A.; Williams, D.; Vega, M.; O’Reilly-Wapstra, J. Effects of thinning on the longitudinal and radial variation in wood properties of Eucalyptus nitens. Forestry 2022, 95, 504–517. [Google Scholar] [CrossRef]
- Alcorn, P.J.; Forrester, D.I.; Thomas, D.S.; James, R.; Smith, R.G.B.; Nicotra, A.B.; Bauhus, J. Changes in whole-tree water use following live-crown pruning in young plantation-grown Eucalyptus pilularis and Eucalyptus cloeziana. Forests 2013, 4, 106–121. [Google Scholar] [CrossRef]
- Buyinza, J.; Muthuri, C.W.; Denton, M.D.; Nuberg, I.K. Impact of tree pruning on water use in tree-coffee systems on smallholder farms in Eastern Uganda. Agrofor. Syst. 2023, 97, 953–964. [Google Scholar] [CrossRef]
- Crockford, R.; Richardson, D. Partitioning of rainfall in a eucalypt forest and pine plantation in southeastern Australia: IV The relationship of interception and canopy storage capacity, the interception of these forests, and the effect on interception of thinning the pine plantation. Hydrol. Process. 1990, 4, 169–188. [Google Scholar] [CrossRef]
- Hua, L.; Yu, F.; Qiu, Q.; He, Q.; Su, Y.; Liu, X.; Li, J. Dry-season irrigation further promotes the growth of Eucalyptus urophylla × E. grandis plantations under the conventional fertilization. New For. 2023, 54, 1085–1102. [Google Scholar] [CrossRef]
- Yu, F.; Truong, T.V.; He, Q.; Hua, L.; Su, Y.; Li, J. Dry season irrigation promotes leaf growth in Eucalyptus urophylla × E. grandis under fertilization. Forests 2019, 10, 67. [Google Scholar] [CrossRef]
- Dodd, I.C. Abscisic acid and stomatal closure: A hydraulic conductance conundrum? New Phytol. 2013, 197, 6–8. [Google Scholar] [CrossRef] [PubMed]
- Verslues, P.E. ABA and cytokinins: Challenge and opportunity for plant stress research. Plant Mol. Biol. 2016, 91, 629–640. [Google Scholar] [CrossRef]
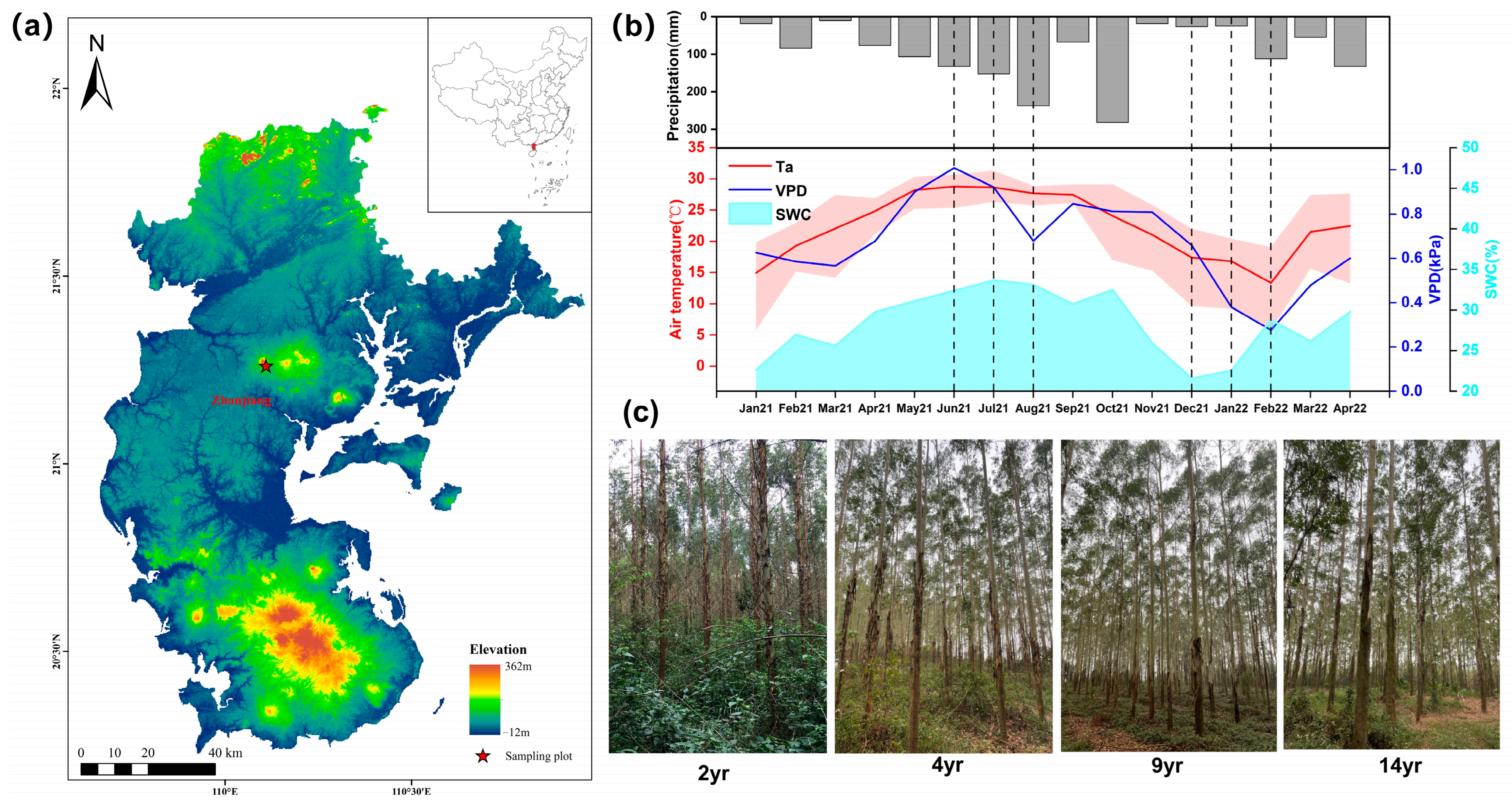






| Stand Age (Years) | Stand Density (Trees·ha−1) | Mean DBH (cm) | Mean Height (m) | Leaf Area Index (LAI) | Crown Breadth (m) | Mean Annual Increment (m3 cha−1·yr) |
|---|---|---|---|---|---|---|
| 2 | 1666 | 9.1 (1.44) | 11.5 (0.64) | 1.24 (0.14) | 2.3 (0.81) | 22.50 (10.51) |
| 4 | 1666 | 12.8 (1.02) | 16.4 (0.72) | 1.79 (0.21) | 3.1 (1.15) | 24.78 (9.13) |
| 9 | 1500 | 19.4 (1.61) | 19.7 (1.02) | 2.08 (0.14) | 4.8 (1.36) | 26.68 (6.45) |
| 14 | 1300 | 24.7 (2.04) | 27.1 (1.36) | 2.48 (0.31) | 5.6 (1.74) | 27.05 (4.63) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Xu, Y.; Zhu, W.; Huang, R.; Du, A.; Cao, H.; Xiang, W. Responses of Water Use Strategies to Seasonal Drought Stress Differed Among Eucalyptus urophylla S.T.Blake × E. grandis Plantations Along with Stand Ages. Forests 2025, 16, 962. https://doi.org/10.3390/f16060962
Wang Z, Xu Y, Zhu W, Huang R, Du A, Cao H, Xiang W. Responses of Water Use Strategies to Seasonal Drought Stress Differed Among Eucalyptus urophylla S.T.Blake × E. grandis Plantations Along with Stand Ages. Forests. 2025; 16(6):962. https://doi.org/10.3390/f16060962
Chicago/Turabian StyleWang, Zhichao, Yuxing Xu, Wankuan Zhu, Runxia Huang, Apeng Du, Haoyang Cao, and Wenhua Xiang. 2025. "Responses of Water Use Strategies to Seasonal Drought Stress Differed Among Eucalyptus urophylla S.T.Blake × E. grandis Plantations Along with Stand Ages" Forests 16, no. 6: 962. https://doi.org/10.3390/f16060962
APA StyleWang, Z., Xu, Y., Zhu, W., Huang, R., Du, A., Cao, H., & Xiang, W. (2025). Responses of Water Use Strategies to Seasonal Drought Stress Differed Among Eucalyptus urophylla S.T.Blake × E. grandis Plantations Along with Stand Ages. Forests, 16(6), 962. https://doi.org/10.3390/f16060962