
Response Mechanism of Carbon Fluxes in Restored and Natural Mangrove Ecosystems Under the Effects of Storm Surges


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Data Collection and Preprocessing
2.3. Storm Surge Impacts on Carbon Exchange
2.4. Statistical Analysis
3. Results
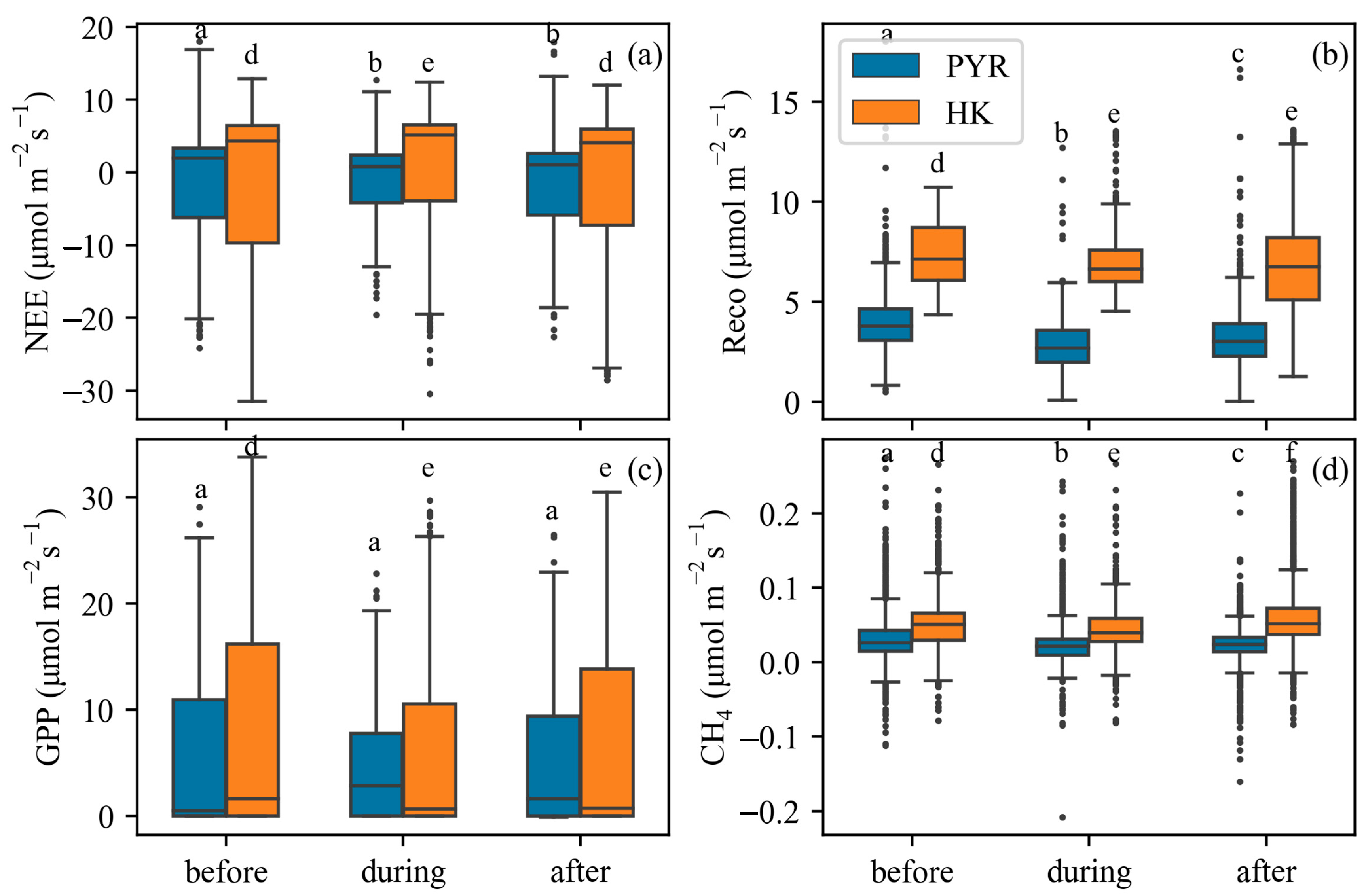
3.1. Temporal Variations of Carbon Fluxes Before and After Storm Surges
3.2. Storm Surges on Carbon Fluxes and Meteorological Variables
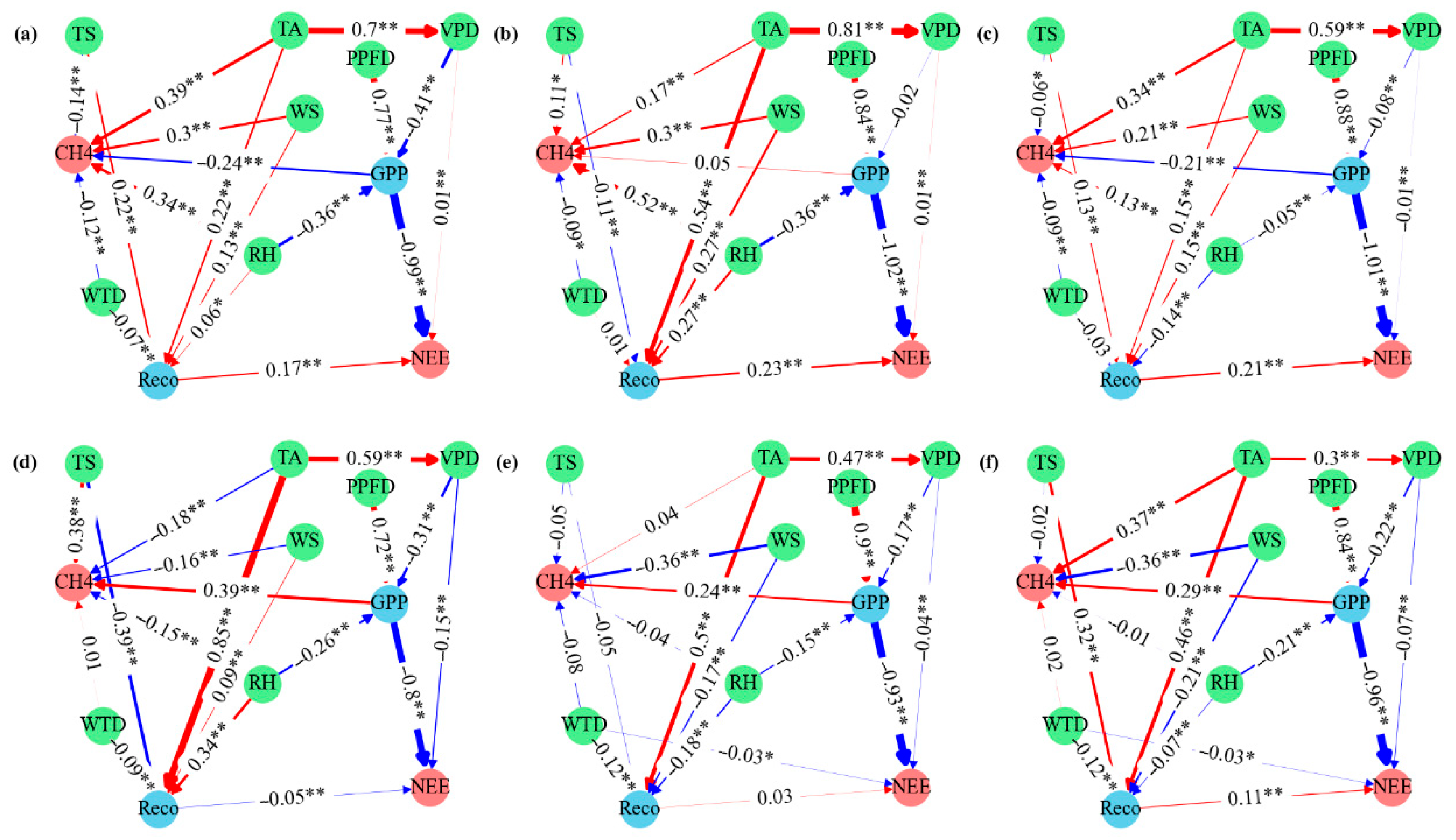
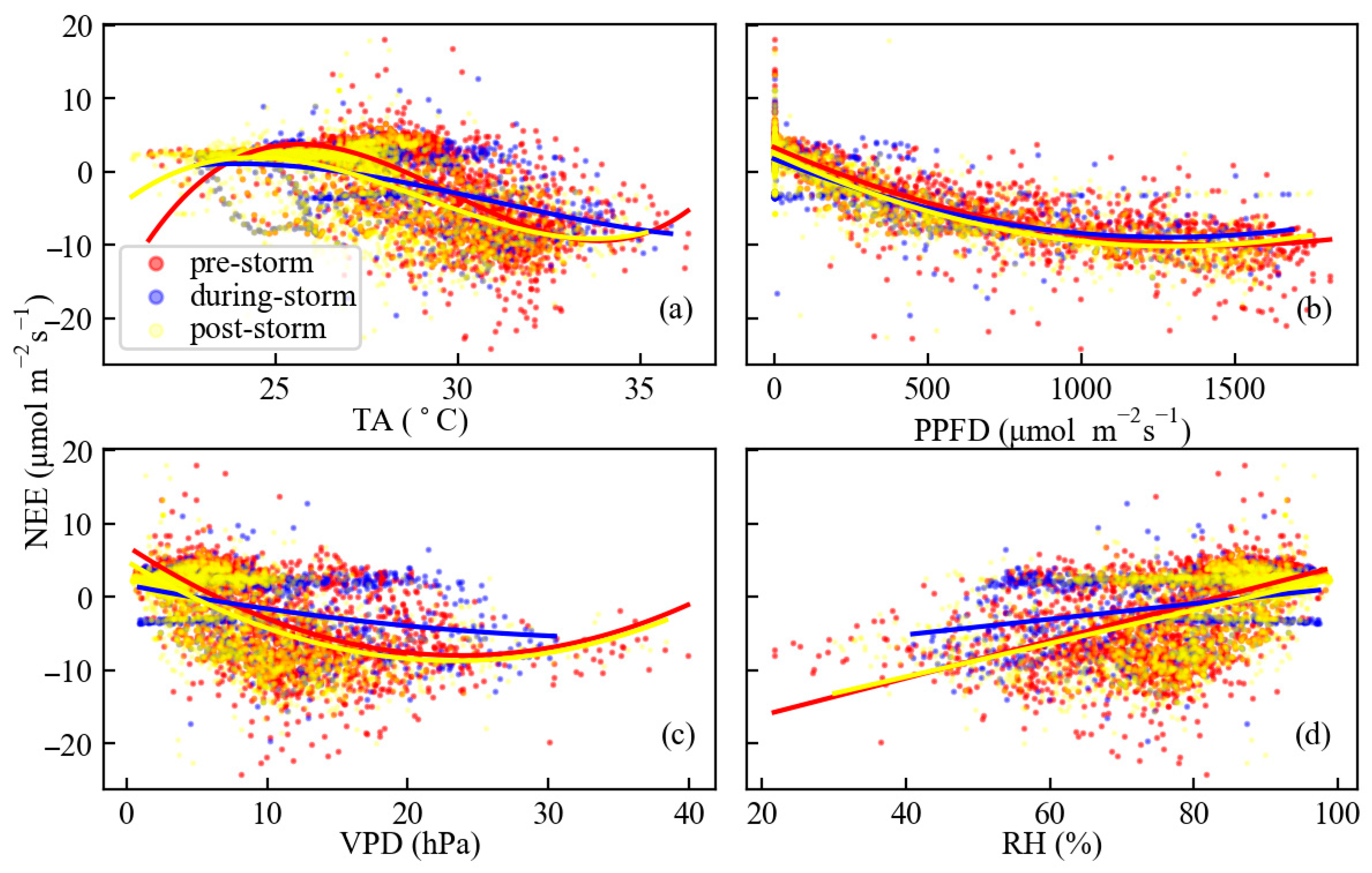
3.3. Relationships Between Carbon Fluxes and Meteorological Variables
4. Discussion
4.1. Impact of Storm Surges on Carbon Fluxes (NEE, GPP, Reco, and CH4)
4.2. Environmental Drivers of Carbon Flux Variability
4.3. Site-Specific Responses to Storm Intensity
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D.; Kurnianto, S.; Stidham, M.; Kanninen, M. Mangroves among the most carbon-rich forests in the tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Liu, J.G.; Zhou, Y.L.; Valach, A.; Shortt, R.; Kasak, K.; Rey-Sanchez, C.; Hemes, K.S.; Baldocchi, D.; Lai, D.Y.F. Methane emissions reduce the radiative cooling effect of a subtropical estuarine mangrove wetland by half. Glob. Change Biol. 2020, 26, 4998–5016. [Google Scholar] [CrossRef] [PubMed]
- Intergovernmental Panel on Climate Change. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Ji, T.; Li, G.S.; Liu, Y.X.Y.; Liu, R.; Zhu, Y.Q. Spatiotemporal features of storm surge activity and its response to climate change in the southeastern coastal area of China in the past 60 years. J. Geophys. Res. Atmos. 2021, 126, e2020JD033234. [Google Scholar] [CrossRef]
- Lin, G.M.; He, Y.N.; Lu, J.G.; Chen, H.; Feng, J.X. Seasonal variations in soil physicochemical properties and microbial community structure influenced by Spartina alterniflora invasion and Kandelia obovata restoration. Sci. Total Environ. 2021, 797, 149213. [Google Scholar] [CrossRef] [PubMed]
- Baldocchi, D.D. How eddy covariance flux measurements have contributed to our understanding of Global Change Biology. Glob. Change Biol. 2019, 26, 242–260. [Google Scholar] [CrossRef]
- Alongi, D.M. Carbon sequestration in mangrove forests. Carbon Manag. 2012, 3, 313–322. [Google Scholar] [CrossRef]
- Alongi, D.M. Mangrove forests: Resilience, protection from tsunamis, and responses to global climate change. Estuar. Coast. Shelf Sci. 2008, 76, 1–13. [Google Scholar] [CrossRef]
- Alvarado-Barrientos, M.S.; López-Adame, H.; Lazcano-Hernández, H.E.; Arellano-Verdejo, J.; Hernández-Arana, H.A. Ecosystem-atmosphere exchange of CO2, water, and energy in a basin mangrove of the northeastern coast of the Yucatan Peninsula. J. Geophys. Res. Biogeosci. 2021, 126, e2020JG005811. [Google Scholar] [CrossRef]
- Zhu, X.D.; Sun, C.Y.; Qin, Z.C. Drought-induced salinity enhancement weakens mangrove greenhouse gas cycling. J. Geophys. Res. Biogeosci. 2021, 126, e2021JG006416. [Google Scholar] [CrossRef]
- Zhao, X.D.; Wang, C.L.; Li, T.T.; Zhang, C.Y.; Fan, X.W.; Zhang, Q.; Chen, X.Y.; Zou, X.K.; Shen, C.; Tang, Y.; et al. Net CO2 and CH4 emissions from restored mangrove wetland: New insights based on a case study in estuary of the Pearl River, China. Sci. Total Environ. 2021, 811, 151619. [Google Scholar] [CrossRef]
- Chen, Y.J.; Zheng, D.Z.; Liao, B.W.; Zheng, S.F.; Zan, Q.J.; Song, X.Y. Researches on typhoon damage to mangroves and preventive measures. For. Res. 2000, 13, 524–529. (In Chinese) [Google Scholar]
- Wang, Y.S.; Gu, J.D. Ecological responses, adaptation and mechanisms of mangrove wetland ecosystem to global climate change and anthropogenic activities. Int. Biodeterior. Biodegrad. 2021, 162, 105248. [Google Scholar] [CrossRef]
- Xu, Z.; Li, X.L.; Tian, P.P.; Huang, Y.T.; Zhu, Q.S.; Zou, H.M.; Huang, Y.; Zhang, Z.; Zhang, S.M.; Chen, M.N.; et al. Global warming potentials of CO2 uptake, CH4 emissions, and albedo changes in a restored mangrove ecosystem. J. Geophys. Res. Biogeosci. 2024, 129, e2023JG007924. [Google Scholar] [CrossRef]
- Liu, J.G.; Schäfer, K.V.R.; Lai, D.Y.F. Biosphere-atmosphere exchange of CO2 and CH4 in mangrove forests and salt marshes. In Carbon Mineralization in Coastal Wetlands; Ouyang, X., Lee, S.Y., Lai, D.Y.F., Marchand, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; Volume 2, pp. 93–132. [Google Scholar]
- Lai, D.Y.F.; Liu, J.G. (2016–2018) FLUXNET-CH4 HK-MPM Mai Po Mangrove, Dataset. Available online: https://fluxnet.org/doi/FLUXNET-CH4/HK-MPM (accessed on 3 July 2025).
- Wutzler, T.; Lucas-Moffat, A.; Migliavacca, M.; Knauer, J.; Sickel, K.; Šigut, L.; Menzer, O.; Reichstein, M. Basic and extensible post-processing of eddy covariance flux data with REddyProc. Biogeosciences 2018, 15, 5015–5030. [Google Scholar] [CrossRef]
- Liu, J.G.; Lai, D.Y.F. Subtropical mangrove wetland is a stronger carbon dioxide sink in the dry than wet seasons. Agric. For. Meteorol. 2019, 278, 107644. [Google Scholar] [CrossRef]
- Chen, H.; Lu, W.; Yan, G.; Yang, S.; Lin, G. Typhoons exert significant but differential impacts on net ecosystem carbon exchange of subtropical mangrove forests in China. Biogeosciences 2014, 11, 5323–5333. [Google Scholar] [CrossRef]
- Baldocchi, D.D. Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems: Past, present and future. Glob. Change Biol. 2003, 9, 479–492. [Google Scholar] [CrossRef]
- Dang, C.; Morrissey, E.M.; Neubauer, S.C.; Franklin, R.B. Novel microbial community composition and carbon biogeochemistry emerge over time following saltwater intrusion in wetlands. Glob. Change Biol. 2018, 25, 549–561. [Google Scholar] [CrossRef]
- Smith, R.S.; Beauvais, J.; Byers, J.E. Traits of resident saltmarsh plants promote retention of range-expanding mangroves under specific tidal regimes. Estuaries Coasts 2021, 45, 1422–1433. [Google Scholar] [CrossRef]
- Reed, D.; Chavez, S.; Castañeda-Moya, E.; Oberbauer, S.F.; Troxler, T.; Malone, S. Resilience to hurricanes is high in mangrove blue carbon forests. Glob. Change Biol. 2025, 31, e7024. [Google Scholar] [CrossRef]
- Krauss, K.W.; Osland, M.J. Tropical cyclones and the organization of mangrove forests: A review. Ann. Bot. 2020, 125, 213–234. [Google Scholar] [CrossRef]
- Peereman, J.; Hogan, J.A.; Lin, T. Disturbance frequency, intensity and forest structure modulate cyclone-induced changes in mangrove forest canopy cover. Glob. Ecol. Biogeogr. 2022, 31, 37–50. [Google Scholar] [CrossRef]
- Lagomasino, D.; Fatoyinbo, T.; Castañeda-Moya, E.; Cook, B.D.; Montesano, P.M.; Neigh, C.S.R.; Corp, L.A.; Ott, L.E.; Chavez, S.; Morton, D.C. Storm surge and ponding explain mangrove dieback in southwest Florida following Hurricane Irma. Nat. Commun. 2021, 12, 4003. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Lagomasino, D.; Charles, S.P.; Castañeda-Moya, E.; Cook, B.D.; Redwine, J.; Fatoyinbo, L. Quantifying mangrove canopy regrowth and recovery after Hurricane Irma with large-scale repeat airborne lidar in the Florida Everglades. Int. J. Appl. Earth Obs. Geoinf. 2022, 114, 103031. [Google Scholar] [CrossRef]
- Quebbeman, A.W.; Menge, D.N.L.; Arellano, G.; Hall, J.; Wood, T.E.; Zimmerman, J.K.; Uriarte, M. A severe hurricane increases carbon dioxide and methane fluxes and triples nitrous oxide emissions in a tropical forest. Ecosystems 2022, 25, 1754–1766. [Google Scholar] [CrossRef]
- Li, X.; Tang, C.Y.; Cao, Y.J.; Li, X. Carbon, nitrogen and sulfur isotopic features and the associated geochemical processes in a coastal aquifer system of the Pearl River Delta, China. J. Hydrol. 2019, 575, 986–998. [Google Scholar] [CrossRef]
- Jaffé, R.; Elismé, T.; Cabrera, A.C. Organic geochemistry of seasonally flooded rain forest soils: Molecular composition and early diagenesis of lipid components. Org. Geochem. 1996, 25, 9–17. [Google Scholar] [CrossRef]
- Li, X.Y.; Cheng, X.Y.; Cheng, K.K.; Cai, Z.H.; Feng, S.Y.; Zhou, J. The influence of tide-brought nutrients on microbial carbon metabolic profiles of mangrove sediments. Sci. Total Environ. 2024, 906, 167732. [Google Scholar] [CrossRef]
- Craig, H.; Antwis, R.E.; Cordero, I.; Ashworth, D.; Robinson, C.H.; Osborne, T.Z.; Bardgett, R.D.; Rowntree, J.K.; Simpson, L.T. Nitrogen addition alters composition, diversity, and functioning of microbial communities in mangrove soils: An incubation experiment. Soil Biol. Biochem. 2021, 153, 108076. [Google Scholar] [CrossRef]
- Wei, P.P.; Lei, A.P.; Zhou, H.C.; Hu, Z.L.; Wong, Y.S.; Tam, N.F.Y.; Lu, Q. Comparison of microbial community structure and function in sediment between natural regenerated and original mangrove forests in a National Nature Mangrove Reserve, South China. Mar. Pollut. Bull. 2021, 163, 111955. [Google Scholar] [CrossRef]
- Yang, Y.H.; Shi, Y.; Sun, W.J.; Chang, J.F.; Zhu, J.X.; Chen, L.Y.; Wang, X.; Guo, Y.P.; Zhang, H.T.; Yu, L.F.; et al. Terrestrial carbon sinks in China and around the world and their contribution to carbon neutrality. Sci. China Life Sci. 2022, 65, 861–895. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.Y.; Jiang, H.L.; Zhang, C.H.; Xu, J.Y.; Xu, X.; Zhang, T.; Huang, Y.Q.; Xu, X.Q.; Zhao, Y.R.; Hu, Y.H.; et al. Light saturation and temperature jointly dominate the diurnal variation of net ecosystem exchange in grassland ecosystems. Ecol. Indic. 2024, 168, 112737. [Google Scholar] [CrossRef]
- Barr, J.G.; Fuentes, J.D.; Engel, V.; Zieman, J.C. Physiological responses of red mangroves to the climate in the Florida Everglades. J. Geophys. Res. Biogeosci. 2009, 114, G02008. [Google Scholar] [CrossRef]
- Cheeseman, J.M.; Lovelock, C.E. Photosynthetic characteristics of dwarf and fringe Rhizophora mangle L. in a Belizean mangrove. Plant Cell Environ. 2004, 27, 769–780. [Google Scholar] [CrossRef]
- Yao, H.M.; Montagna, P.A.; Wetz, M.S.; Staryk, C.J.; Hu, X.P. Subtropical estuarine carbon budget under various hydrologic extremes and implications on the lateral carbon exchange from tidal wetlands. Water Res. 2022, 217, 118436. [Google Scholar] [CrossRef]
- Barr, J.G.; Engel, V.; Fuentes, J.D.; Zieman, J.C.; O’Halloran, T.L.; Smith, T.J.; Anderson, G.H. Controls on mangrove forest-atmosphere carbon dioxide exchanges in western Everglades National Park. J. Geophys. Res. Biogeosci. 2010, 115, G02020. [Google Scholar] [CrossRef]
- Li, Q.; Lu, W.; Chen, H.; Luo, Y.; Lin, G. Differential responses of net ecosystem exchange of carbon dioxide to light and temperature between spring and neap tides in subtropical mangrove forests. Sci. World J. 2014, 2014, 943697. [Google Scholar] [CrossRef]
- Nuria, G.; Nicholas, J.D.; Evan, H.D.; Elena, B.; Elizabeth, H.B.; Jed, S.; Carl, J.B. Seasonal controls of CO2 and CH4 dynamics in a temporarily flooded subtropical wetland. J. Geophys. Res. Biogeosci. 2020, 125, e2019JG005257. [Google Scholar] [CrossRef]
- Yu, H.; Organ, H.; Xu, D.; Coffin, R. Tidal control and mangrove dieback impact on methane emissions from a subtropical mangrove estuary. Limnol. Oceanogr. 2023, 68, 753–766. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Bernal, B.; Nahlik, A.M.; Mander, Ü.; Zhang, L.; Anderson, C.J.; Jørgensen, S.E.; Brix, H. Wetlands, carbon, and climate change. Landsc. Ecol. 2013, 28, 583–597. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviations | Parameters Description | Units |
|---|---|---|
| NEE | Net ecosystem CO2 exchange | μmol m−2 s−1 |
| GPP | Gross primary productivity | μmol m−2 s−1 |
| Reco | Ecosystem respiration | μmol m−2 s−1 |
| CH4 | Flux of methane | μmol m−2 s−1 |
| VPD | Vapor pressure deficit | hPa |
| RH | Relative humidity | % |
| TA | Air temperature | °C |
| WS | Wind speed | m/s |
| PPFD | Photosynthetic photon flux density | μmol m−2 s−1 |
| TS | Soil temperature | °C |
| WTD | Water table depth | m |
| International Number | Name | Time (yyyy–mm–dd hh:mm) | Wind Speed (m/s) | Min_Distance (km) | Duration (Hour) |
|---|---|---|---|---|---|
| PYR site | |||||
| 2004 | Hagupit | 2020-08-04 02:00 | 38 | 57.68 | 108 |
| 2106 | In-Fa | 2021-07-25 14:00 | 38 | 311.86 | 234 |
| 2114 | Chanthu | 2021-09-13 01:00 | 50 | 242.23 | 261 |
| 2212 | Uifa | 2022-09-14 14:00 | 48 | 249.03 | 202 |
| HK site | |||||
| 1604 | Nida | 2016-08-02 06:00 | 33 | 11.5 | 84 |
| 1622 | Haima | 2016-10-21 13:00 | 42 | 114.89 | 162 |
| 1702 | Merbok | 2017-06-12 22:00 | 25 | 44.07 | 45 |
| 1713 | Hato | 2017-08-23 10:00 | 48 | 77.85 | 111 |
| 1714 | Pakhar | 2017-08-27 08:00 | 33 | 99.08 | 78 |
| 1822 | Mangkhut | 2018-09-16 14:00 | 48 | 123.52 | 237 |
| 1823 | Barijat | 2018-09-12 17:00 | 23 | 166.12 | 78 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, H.; Zhu, J.; Tian, Z.; Chen, Z.; Xue, Z.; Li, W. Response Mechanism of Carbon Fluxes in Restored and Natural Mangrove Ecosystems Under the Effects of Storm Surges. Forests 2025, 16, 1115. https://doi.org/10.3390/f16071115
Zou H, Zhu J, Tian Z, Chen Z, Xue Z, Li W. Response Mechanism of Carbon Fluxes in Restored and Natural Mangrove Ecosystems Under the Effects of Storm Surges. Forests. 2025; 16(7):1115. https://doi.org/10.3390/f16071115
Chicago/Turabian StyleZou, Huimin, Jianhua Zhu, Zhen Tian, Zhulin Chen, Zhiyong Xue, and Weiwei Li. 2025. "Response Mechanism of Carbon Fluxes in Restored and Natural Mangrove Ecosystems Under the Effects of Storm Surges" Forests 16, no. 7: 1115. https://doi.org/10.3390/f16071115
APA StyleZou, H., Zhu, J., Tian, Z., Chen, Z., Xue, Z., & Li, W. (2025). Response Mechanism of Carbon Fluxes in Restored and Natural Mangrove Ecosystems Under the Effects of Storm Surges. Forests, 16(7), 1115. https://doi.org/10.3390/f16071115