

Climate Vulnerability Analysis of Marginal Populations of Yew (Taxus baccata L.): The Case of the Iberian Peninsula
 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Species and Study Area
2.2. Distribution and Climatic Data
2.3. Individual Growth Data of Natural Populations of Taxus baccata
2.4. Ecological Niche and Climate Vulnerability Analysis
2.4.1. Climate Suitability Modeling
2.4.2. Using the Individual Performance of Natural Taxus baccata Populations to Validate the Output of the ENMs
2.5. Climate Vulnerability Analysis
3. Results
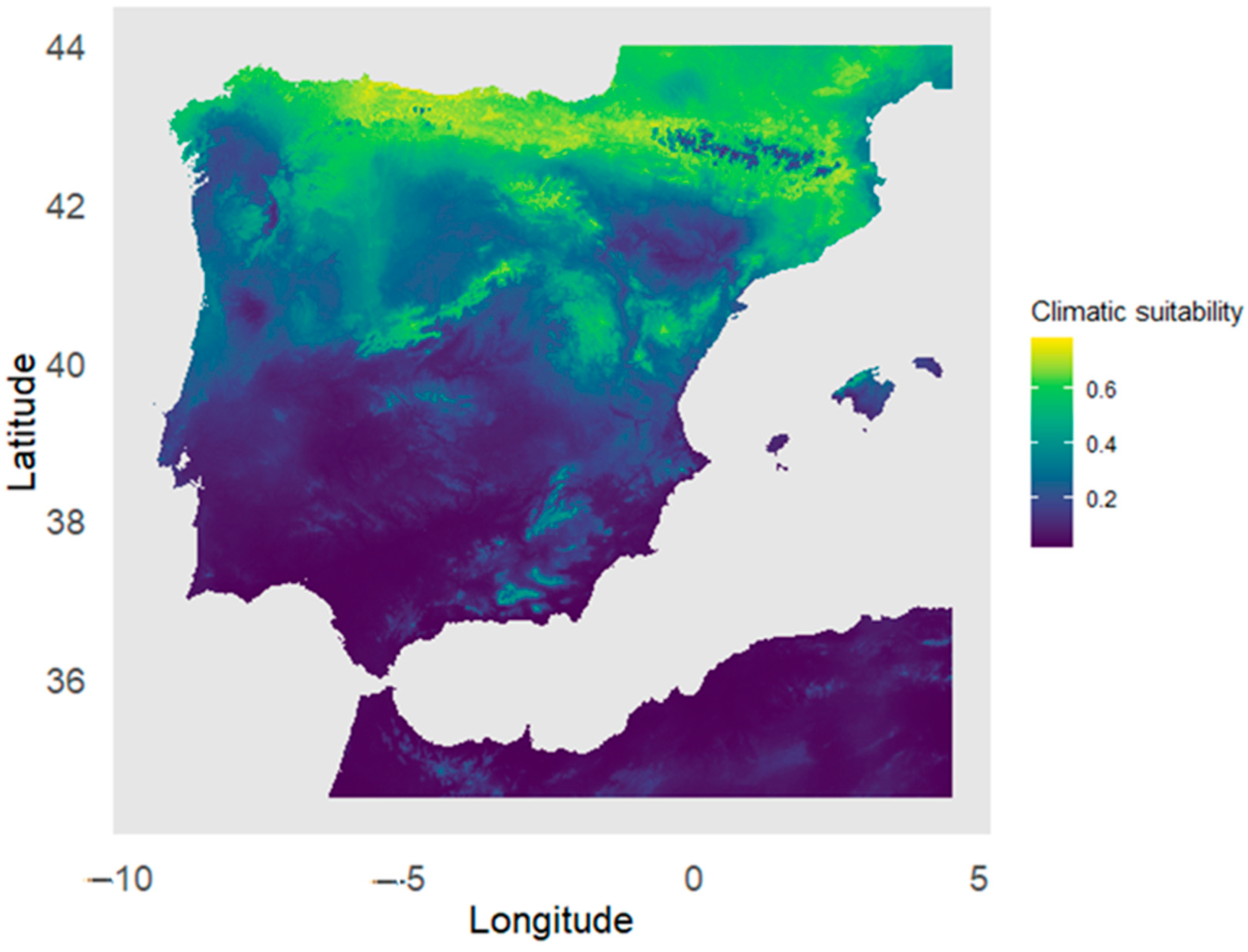
3.1. Climatic Suitability Analysis of Taxus baccata Habitat in the Iberian Peninsula
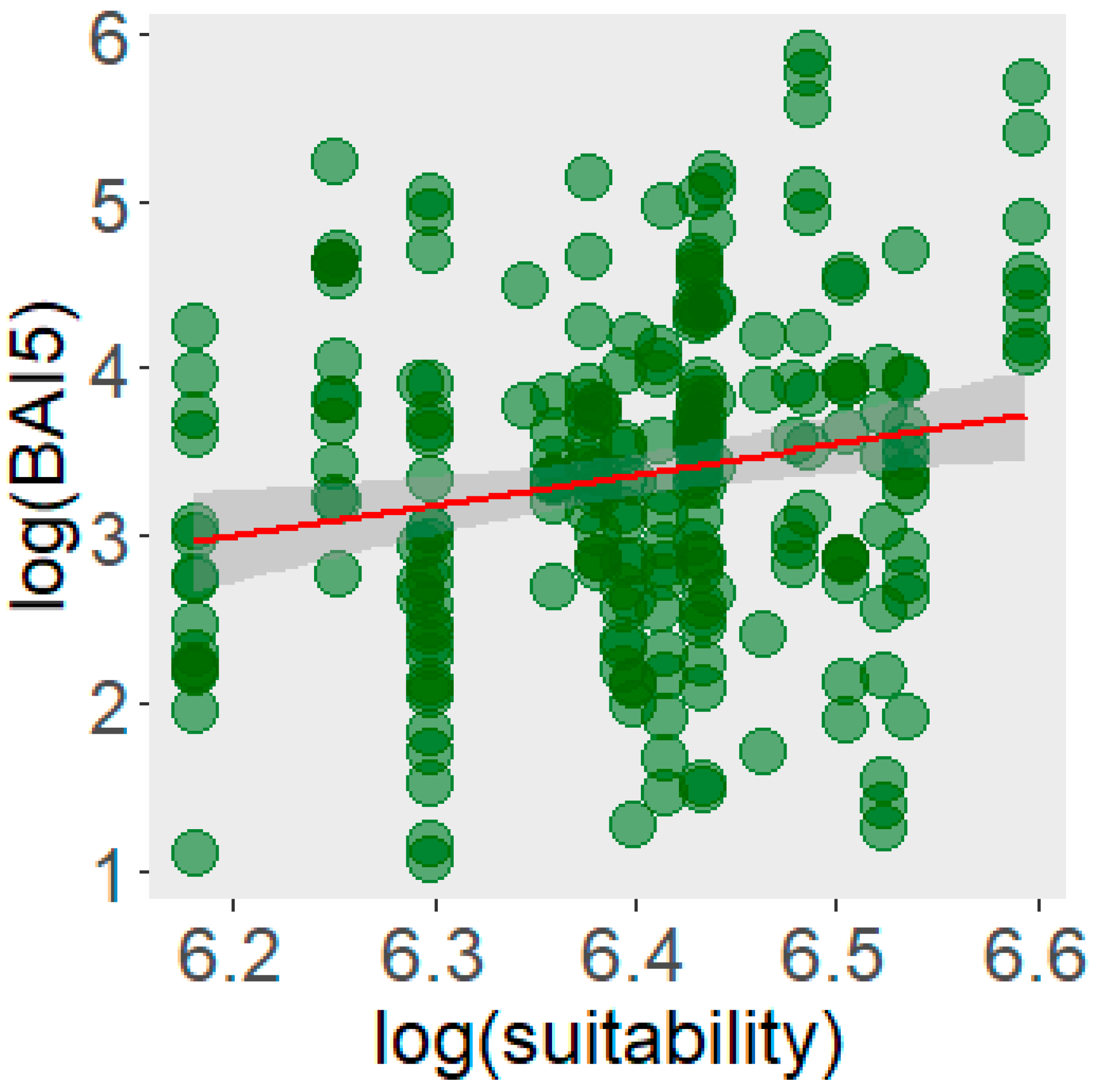
3.2. Predicted Climatic Habitat Suitability and Individual Performance of Natural Populations of Taxus baccata
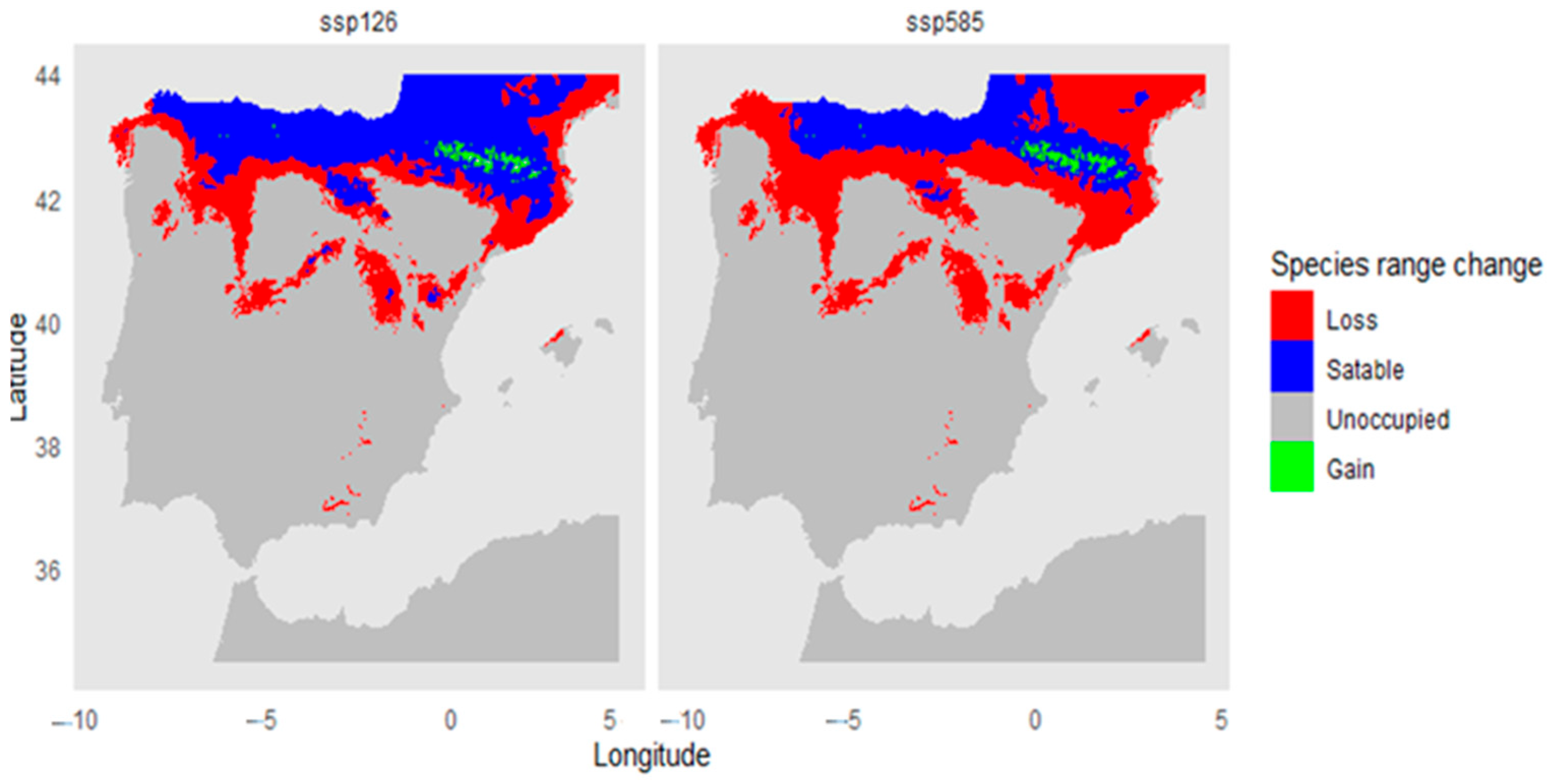
3.3. Climatic Vulnerability Analysis of Taxus baccata in the Iberian Peninsula
4. Discussion
4.1. Climatic Habitat Suitability and Regional Future Trends
4.2. Relationship Between Individual Growth and Habitat Suitability, and Regional Trends Under Future Scenarios
4.3. Management Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abeli, T.; Ghitti, M.; Sacchi, R. Does Ecological Marginality Reflect Physiological Marginality in Plants? Plant Biosyst. 2020, 154, 149–157. [Google Scholar] [CrossRef]
- Koç, D.; Svenning, J.-C.; Avci, M. Climate Change Impacts on the Potential Distribution of Taxus baccata L. in the Eastern Mediterranean and the Bolkar Mountains (Turkey) from Last Glacial Maximum to the Future. Eurasian J. For. Sci. 2018, 6, 69–82. [Google Scholar] [CrossRef]
- Iszkuło, G. Success and Failure of Endangered Tree Species: Low Temperatures and Low Light Availability Affect Survival and Growth of European Yew (Taxus baccata L.) Seedlings. Pol. J. Ecol. 2010, 58, 259–271. [Google Scholar]
- Alavi, S.J.; Ahmadi, K.; Hosseini, S.M.; Tabari, M.; Nouri, Z. The Response of English Yew (Taxus baccata L.) to Climate Change in the Caspian Hyrcanian Mixed Forest Ecoregion. Reg. Env. Change 2019, 19, 1495–1506. [Google Scholar] [CrossRef]
- Hernández-Lambraño, R.E.; de la Cruz, D.R.; Agudo, J.Á.S. Effects of the Climate Change on Peripheral Populations of Hydrophytes: A Sensitivity Analysis for European Plant Species Based on Climate Preferences. Sustainability 2021, 13, 3147. [Google Scholar] [CrossRef]
- Vilà-Cabrera, A.; Premoli, A.C.; Jump, A.S. Refining Predictions of Population Decline at Species’ Rear Edges. Glob. Change Biol. 2019, 25, 1549–1560. [Google Scholar] [CrossRef]
- Hampe, A.; Petit, R.J. Conserving Biodiversity under Climate Change: The Rear Edge Matters. Ecol. Lett. 2005, 8, 461–467. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; de Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction Risk from Climate Change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Urban, M.C. Accelerating Extinction Risk from Climate Change. Science 2015, 348, 571–573. [Google Scholar] [CrossRef]
- Stanton, J.C.; Shoemaker, K.T.; Pearson, R.G.; Akçakaya, H.R. Warning Times for Species Extinctions Due to Climate Change. Glob. Change Biol. 2015, 21, 1066–1077. [Google Scholar] [CrossRef]
- IUCN Red List. IUCN Red List. IUCN Red List of Threatened Species: Taxus baccata. In IUCN Red List of Threatened Species; IUCN Red List: Cambridge, UK, 2010. [Google Scholar]
- Decreto 63/2007, de 14 de Junio, por el Que se Crean el Catálogo de Flora Protegida de Castilla y León y la Figura de Protección Denominada Microrreserva de Flora. Available online: https://bocyl.jcyl.es/boletines/2007/06/20/pdf/BOCYL-D-20062007-3.pdf (accessed on 29 May 2025).
- Uzquiano, P.; Allué, E.; Antolín, F.; Burjachs, F.; Picornel, L.; Piqué, R.; Zapata, L. All about Yew: On the Trail of Taxus baccata in Southwest Europe by Means of Integrated Palaeobotanical and Archaeobotanical Studies. Veget. Hist. Archaeobot. 2015, 24, 229–247. [Google Scholar] [CrossRef]
- Thomas, P.; Garcia-Marti, X. Response of European Yews to Climate Change: A Review. For. Syst. 2015, 24, eR01. [Google Scholar] [CrossRef]
- Iszkuło, G.; Didukh, Y.; Giertych, M.J.; Jasińska, A.K.; Sobierajska, K.; Szmyt, J. Weak Competitive Ability May Explain Decline of Taxus baccata. Ann. For. Sci. 2012, 69, 705–712. [Google Scholar] [CrossRef]
- Song, Y.; Sass-Klaassen, U.; Sterck, F.; Goudzwaard, L.; Akhmetzyanov, L.; Poorter, L. Growth of 19 Conifer Species Is Highly Sensitive to Winter Warming, Spring Frost and Summer Drought. Ann. Bot. 2021, 128, 545–557. [Google Scholar] [CrossRef] [PubMed]
- Linares, J.C. Shifting Limiting Factors for Population Dynamics and Conservation Status of the Endangered English Yew (Taxus baccata L., Taxaceae). For. Ecol. Manag. 2013, 291, 119–127. [Google Scholar] [CrossRef]
- Thuiller, W.; Georges, D.; Engler, R. Biomod2: Ensemble Platform for Species Distribution Modelling; The R Foundation: Vienna, Austria, 2014; Volume 2. [Google Scholar]
- Hernández-Lambraño, R.E.; González-Moreno, P.; Sánchez-Agudo, J.Á.; Hernández-Lambraño, R.E.; González-Moreno, P.; Sánchez-Agudo, J.Á. Towards the Top: Niche Expansion of Taraxacum Officinale and Ulex Europaeus in Mountain Regions of South America. Austral Ecol. 2017, 42, 577–589. [Google Scholar] [CrossRef]
- Franklin, J. Mapping Species Distributions: Spatial Inference and Prediction; Cambridge University Press: Cambridge, UK, 2010; ISBN 1-139-48529-6. [Google Scholar]
- Soberon, J.; Peterson, A.T. Interpretation of Models of Fundamental Ecological Niches and Species’ Distributional Areas. Biodivers. Inform. 2005, 2, 1–10. [Google Scholar] [CrossRef]
- Peterson, K.; Bode, M. Using Ensemble Modeling to Predict the Impacts of Assisted Migration on Recipient Ecosystems. Conserv. Biol. 2021, 35, 678–687. [Google Scholar] [CrossRef]
- Sanchez-Martinez, P.; Marcer, A.; Mayol, M.; Riba, M. Shaping the Niche of Taxus baccata, a Modelling Exercise Using Biologically Meaningful Information. For. Ecol. Manag. 2021, 501, 119688. [Google Scholar] [CrossRef]
- Maroso, F.; Vera, M.; Ferreiro, J.; Mayol, M.; Riba, M.; Ramil-Rego, P.; Martínez, P.; Bouza, C. Genetic Diversity and Structure of Taxus baccata from the Cantabrian-Atlantic Area in Northern Spain: A Guide for Conservation and Management Actions. For. Ecol. Manag. 2021, 482, 118844. [Google Scholar] [CrossRef]
- Casals, P.; Camprodon, J.; Caritat, A.; Rios, A.I.; Guixé, D.; Garcia-Marti, X.; Martín-Alcón, S.; Coll, L. Forest Structure of Mediterranean Yew (Taxus baccata L.) Populations and Neighbor Effects on Juvenile Yew Performance in the NE Iberian Peninsula. For. Syst. 2015, 24, e042. [Google Scholar] [CrossRef]
- Karger, D.N.; Conrad, O.; Böhner, J.; Kawohl, T.; Kreft, H.; Soria-Auza, R.W.; Zimmermann, N.E.; Linder, H.P.; Kessler, M. Climatologies at High Resolution for the Earth’s Land Surface Areas. Sci. Data 2017, 4, 170122. [Google Scholar] [CrossRef]
- Hernández-Lambraño, R.E.; Parra, J.L.; Román, J.F.C.; Sánchez-Agudo, J.Á. Less Suitable Climatic Conditions and Pests Increase Tree Defoliation in Spanish Iberian Peninsula Forests. For. Ecol. Manag. 2024, 566, 122048. [Google Scholar] [CrossRef]
- Gent, P.R.; Danabasoglu, G.; Donner, L.J.; Holland, M.M.; Hunke, E.C.; Jayne, S.R.; Lawrence, D.M.; Neale, R.B.; Rasch, P.J.; Vertenstein, M.; et al. The Community Climate System Model Version 4. J. Clim. 2011, 24, 4973–4991. [Google Scholar] [CrossRef]
- The HadGEM2 Development Team; Martin, G.M.; Bellouin, N.; Collins, W.J.; Culverwell, I.D.; Halloran, P.R.; Hardiman, S.C.; Hinton, T.J.; Jones, C.D.; McDonald, R.E.; et al. The HadGEM2 Family of Met Office Unified Model Climate Configurations. Geosci. Model. Dev. 2011, 4, 723–757. [Google Scholar] [CrossRef]
- Albuquerque, F.; Benito, B.; Rodriguez, M.Á.M.; Gray, C. Potential Changes in the Distribution of Carnegiea Gigantea under Future Scenarios. PeerJ 2018, 6, e5623. [Google Scholar] [CrossRef] [PubMed]
- Araújo, M.B.; New, M. Ensemble Forecasting of Species Distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2019. [Google Scholar]
- Hernández-Lambraño, R.E.; Carbonell, R.; Sánchez-Agudo, J. Making the Most of Scarce Data: Mapping Distribution Range and Variation in Population Abundance of a Threatened Narrow-Range Endemic Plant. J. Nat. Conserv. 2020, 57, 125889. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Package Lme4: Linear Mixed-Effects Models Using Eigen and S4; CRAN: Vienna, Austria, 2014; Volume 67. [Google Scholar]
- Jiménez-Valverde, A.; Lobo, J.M.; Hortal, J. Not as Good as They Seem: The Importance of Concepts in Species Distribution Modelling. Divers. Distrib. 2008, 14, 885–890. [Google Scholar] [CrossRef]
- Osorio-Olvera, L.; Soberón, J.; Falconi, M. On Population Abundance and Niche Structure. Ecography 2019, 42, 1415–1425. [Google Scholar] [CrossRef]
- Oldfather, M.F.; Kling, M.M.; Sheth, S.N.; Emery, N.C.; Ackerly, D.D. Range Edges in Heterogeneous Landscapes: Integrating Geographic Scale and Climate Complexity into Range Dynamics. Glob. Change Biol. 2020, 26, 1055–1067. [Google Scholar] [CrossRef] [PubMed]
- García, D.; Ramón Obeso, J. Facilitation by Herbivore-Mediated Nurse Plants in a Threatened Tree, Taxus baccata: Local Effects and Landscape Level Consistency. Ecography 2003, 26, 739–750. [Google Scholar] [CrossRef]
- Sanz, R.; Pulido, F.; Nogués-Bravo, D. Predicting Mechanisms across Scales: Amplified Effects of Abiotic Constraints on the Recruitment of Yew Taxus baccata. Ecography 2009, 32, 993–1000. [Google Scholar] [CrossRef]
- Peterson, A.T.; Papeş, M.; Soberón, J. Rethinking Receiver Operating Characteristic Analysis Applications in Ecological Niche Modeling. Ecol. Model. 2008, 213, 63–72. [Google Scholar] [CrossRef]
- Ahmadi, K.; Alavi, S.J.; Amiri, G.Z.; Hosseini, S.M.; Serra-Diaz, J.M.; Svenning, J.-C. The Potential Impact of Future Climate on the Distribution of European Yew (Taxus baccata L.) in the Hyrcanian Forest Region (Iran). Int. J. Biometeorol. 2020, 64, 1451–1462. [Google Scholar] [CrossRef]
- Dubreuil, M.; Riba, M.; González-Martínez, S.C.; Vendramin, G.G.; Sebastiani, F.; Mayol, M. Genetic Effects of Chronic Habitat Fragmentation Revisited: Strong Genetic Structure in a Temperate Tree, Taxus baccata (Taxaceae), with Great Dispersal Capability. Am. J. Bot. 2010, 97, 303–310. [Google Scholar] [CrossRef]
- González-Martínez, S.C.; Dubreuil, M.; Riba, M.; Vendramin, G.G.; Sebastiani, F.; Mayol, M. Spatial Genetic Structure of Taxus baccata L. in the Western Mediterranean Basin: Past and Present Limits to Gene Movement over a Broad Geographic Scale. Mol. Phylogenet. Evol. 2010, 55, 805–815. [Google Scholar] [CrossRef]
- Allen, C.; Macalady, A.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.; Hogg, E.H.; et al. A Global Overview of Drought and Heat-Induced Tree Mortality Reveals Emerging Climate Change Risks for Forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Hammond, W.M.; Williams, A.P.; Abatzoglou, J.T.; Adams, H.D.; Klein, T.; López, R.; Sáenz-Romero, C.; Hartmann, H.; Breshears, D.D.; Allen, C.D. Global Field Observations of Tree Die-off Reveal Hotter-Drought Fingerprint for Earth’s Forests. Nat. Commun. 2022, 13, 1761. [Google Scholar] [CrossRef]
- Selwood, K.E.; Zimmer, H.C. Refuges for Biodiversity Conservation: A Review of the Evidence. Biol. Conserv. 2020, 245, 108502. [Google Scholar] [CrossRef]
- Ahmadi, K.; Jalil Alavi, S.; Zahedi Amiri, G.; Mohsen Hosseini, S.; Serra-Diaz, J.M.; Svenning, J.-C. Patterns of Density and Structure of Natural Populations of Taxus baccata in the Hyrcanian Forests of Iran. Nord. J. Bot. 2020, 38. [Google Scholar] [CrossRef]
- Boisvert-Marsh, L.; Périé, C.; de Blois, S. Shifting with Climate? Evidence for Recent Changes in Tree Species Distribution at High Latitudes. Ecosphere 2014, 5, art83. [Google Scholar] [CrossRef]
- Rew, L.J.; McDougall, K.L.; Alexander, J.M.; Daehler, C.C.; Essl, F.; Haider, S.; Kueffer, C.; Lenoir, J.; Milbau, A.; Nuñez, M.A.; et al. Moving up and over: Redistribution of Plants in Alpine, Arctic, and Antarctic Ecosystems under Global Change. Arct. Antarct. Alp. Res. 2020, 52, 651–665. [Google Scholar] [CrossRef]
- Lavabre, J.E.; García, D. Geographic Consistency in the Seed Dispersal Patterns of Taxus baccata L. in the Iberian Peninsula. For. Syst. 2015, 24, e040. [Google Scholar] [CrossRef]
- Chybicki, I.J.; Oleksa, A. Seed and Pollen Gene Dispersal in Taxus baccata, a Dioecious Conifer in the Face of Strong Population Fragmentation. Ann. Bot. 2018, 122, 409–421. [Google Scholar] [CrossRef] [PubMed]
- Pautasso, M. Geographical Genetics and the Conservation of Forest Trees. Perspect. Plant Ecol. Evol. Syst. 2009, 11, 157–189. [Google Scholar] [CrossRef]
- Smith, W.P.; Zollner, P.A. Sustainable Management of Wildlife Habitat and Risk of Extinction. Biol. Conserv. 2005, 125, 287–295. [Google Scholar] [CrossRef]
- Mahmoodi, S.; Ahmadi, K.; Heydari, M.; Karami, O.; Esmailzadeh, O.; Heung, B. Elevational Shift of Endangered European Yew under Climate Change in Hyrcanian Mountain Forests: Rethinking Conservation-Restoration Strategies and Management. For. Ecol. Manag. 2023, 529, 120693. [Google Scholar] [CrossRef]
- Piovesan, G.; Presutti Saba, E.; Biondi, F.; Alessandrini, A.; Di Filippo, A.; Schirone, B. Population Ecology of Yew (Taxus baccata L.) in the Central Apennines: Spatial Patterns and Their Relevance for Conservation Strategies. Plant Ecol. 2009, 205, 23–46. [Google Scholar] [CrossRef]
- Stanturf, J.A.; Ivetic, V.; Dumroese, R.K. Framing Recent Advances in Assisted Migration of Trees: A Special Issue. For. Ecol. Manag. 2024, 551, 121552. [Google Scholar] [CrossRef]
- Calvia, G.; Casula, P.; Farris, E.; Fenu, G.; Fantini, S.; Bacchetta, G. Shrub Cover and Soil Moisture Affect Taxus baccata L. Regeneration at Its Southern Range. Plants 2023, 12, 1819. [Google Scholar] [CrossRef] [PubMed]
- Perry, A.; Aravanopoulos, F.A.; Budde, K.B.; Hansen, O.K.; Rellstab, C.; Schroeder, H.; Curtu, A.L. Resilient Forests for the Future. Tree Genet. Genomes 2024, 20, 17. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Models | Climatic Variables | |||
|---|---|---|---|---|
| bio01 | bio04 | bio12 | bio15 | |
| GBM | 0.11 ± 0.04 (−) | 0.05 ± 0.05 (−) | 0.24 ± 0.03 (=) | 0.38 ± 0.10 (=) |
| GLM | 0.10 ± 0.03 (−) | 0.07 ± 0.03 (−) | 0.19 ± 0.05 (=) | 0.35 ± 0.03 (=) |
| MAXENT | 0.07 ± 0.02 (−) | 0.07 ± 0.01 (−) | 0.21 ± 0.03 (Ω) | 0.43 ± 0.03 (−) |
| RF | 0.41 ± 0.04 (−) | 0.39 ± 0.06 (−) | 0.40 ± 0.05 (+) | 0.60 ± 0.0 (−) |
| EM | 0.08 ± 0.00 (−) | 0.06 ± 0.00 (−) | 0.19 ± 0.00 (Ω) | 0.39 ± 0.00 (−) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz Román, J.F.; Hernández-Lambraño, R.E.; Rodríguez-de la Cruz, D.; Sánchez-Agudo, J.Á. Climate Vulnerability Analysis of Marginal Populations of Yew (Taxus baccata L.): The Case of the Iberian Peninsula. Forests 2025, 16, 931. https://doi.org/10.3390/f16060931
Cruz Román JF, Hernández-Lambraño RE, Rodríguez-de la Cruz D, Sánchez-Agudo JÁ. Climate Vulnerability Analysis of Marginal Populations of Yew (Taxus baccata L.): The Case of the Iberian Peninsula. Forests. 2025; 16(6):931. https://doi.org/10.3390/f16060931
Chicago/Turabian StyleCruz Román, Jhony Fernando, Ricardo Enrique Hernández-Lambraño, David Rodríguez-de la Cruz, and José Ángel Sánchez-Agudo. 2025. "Climate Vulnerability Analysis of Marginal Populations of Yew (Taxus baccata L.): The Case of the Iberian Peninsula" Forests 16, no. 6: 931. https://doi.org/10.3390/f16060931
APA StyleCruz Román, J. F., Hernández-Lambraño, R. E., Rodríguez-de la Cruz, D., & Sánchez-Agudo, J. Á. (2025). Climate Vulnerability Analysis of Marginal Populations of Yew (Taxus baccata L.): The Case of the Iberian Peninsula. Forests, 16(6), 931. https://doi.org/10.3390/f16060931