


Planting Trees as a Nature-Based Solution to Mitigate Climate Change: Opportunities, Limits, and Trade-Offs



Abstract
1. Introduction
2. Methods
3. Atmospheric [CO2] and Potential Sequestration by Trees and Forests
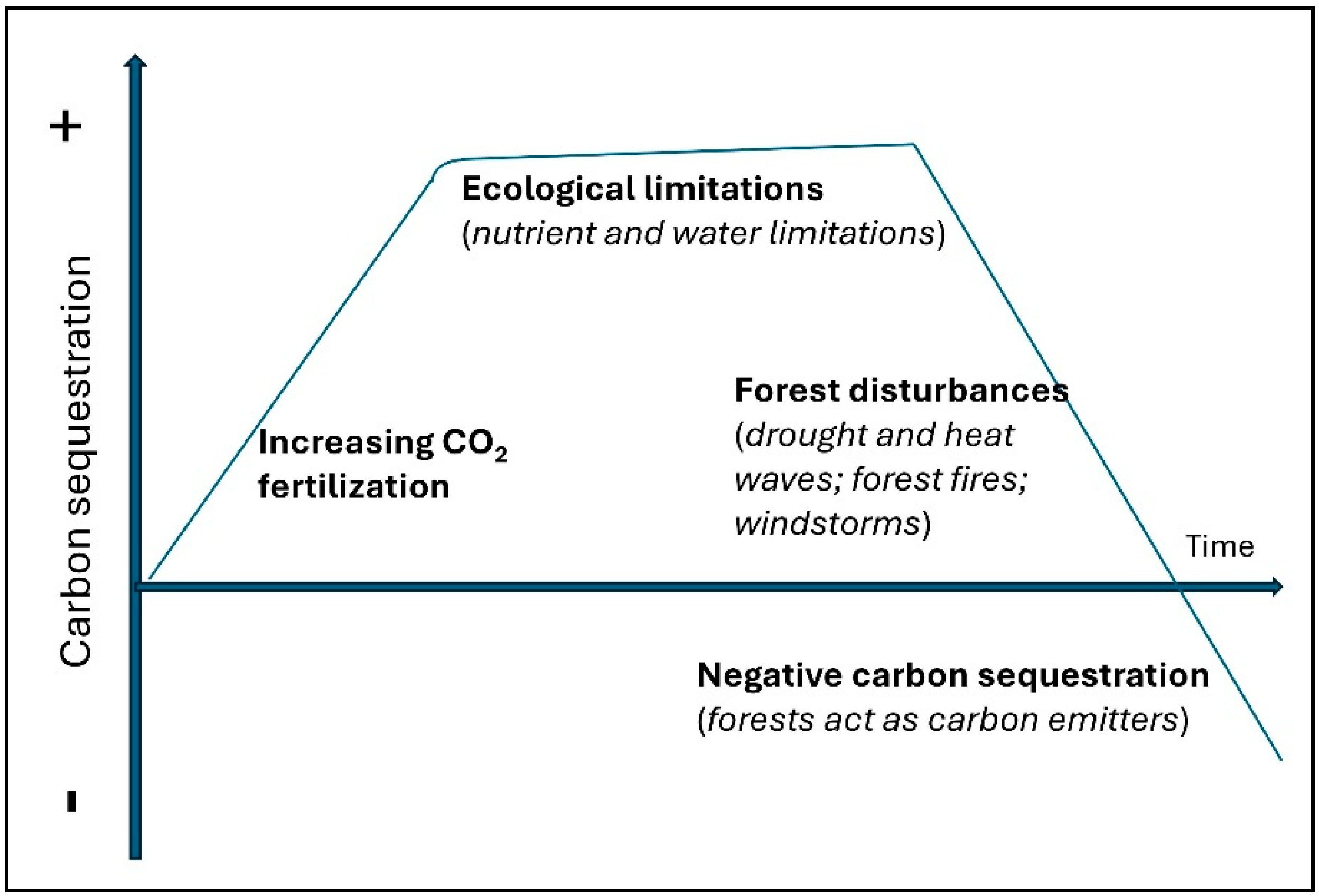
4. Trade-Offs and Limitations
4.1. Growth and Persistence
4.2. How Trees Interact with Climate
4.3. Stimulation and Limitation of Tree Growth Induced by Climate Change Factors
4.4. Forest Disturbances
5. Plantation and Forest: Guidelines for Climate-Smart Management
5.1. Tree Species Selection for “Ad Hoc” Plantations
5.2. Fast-Growing Plantations
5.3. Managed Forests
5.4. Unmanaged Forests
6. Discussion
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- IPCC. Global Warming of 1.5 °C. In An IPCC Special Report on the Impacts of Global Warming of 1.5 °C Above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Masson-Delmotte, V.P., Zhai, H.-O., Pörtner, D., Roberts, J., Skea, P.R., Shukla, A., Pirani, W., Moufouma-Okia, C., Péan, R., Pidcock, S., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2018; 616p. [Google Scholar] [CrossRef]
- Lewis, S.L.; Wheeler, C.E.; Mitchard, E.T.A.; Koch, A. Regenerate natural forests to store carbon. Nature 2019, 568, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.W.; Thawale, P.R.; Sharma, J.K.; Gautam, R.K.; Kundargi, G.P.; Juwarkar, A.A. Chapter 3—Carbon Sequestration in Terrestrial Ecosystems. In Hydrogen Production and Remediation of Carbon and Pollutants; Lichtfouse, E., Schwarzbauer, J., Eds.; Environmental Chemistry for a Sustainable World 6; Springer International Publishing: Cham, Switzerland, 2015; pp. 99–130. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A Large and Persistent Carbon Sink in the World’s Forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef]
- FAO. State of the World Forests; Forest-Sector Innovations Towards a More Sustainable Future; Food and Agriculture Organization of the United Nations Rome: Rome, Italy, 2024; p. 122. [Google Scholar]
- Goymer, P. A trillion trees. Nat. Ecol. Evol. 2018, 2, 208–209. [Google Scholar] [CrossRef]
- Available online: https://environment.ec.europa.eu/strategy/biodiversity-strategy-2030_en (accessed on 10 April 2025).
- Available online: https://climate.ec.europa.eu/eu-action/carbon-removals-and-carbon-farming_en (accessed on 10 April 2025).
- Harde, H. Understanding Increasing Atmospheric CO2. Sci. Clim. Chang. 2023, 3, 46–67. [Google Scholar] [CrossRef]
- Available online: https://www.climate.gov/news-features/understanding-climate/climate-change-atmospheric-carbon-dioxide (accessed on 10 April 2025).
- Bastin, J.-F.; Finegold, Y.; Garcia, C.; Mollicone, D.; Rezende, M.; Routh, D.; Zohner, C.M.; Crowther, T.W. The global tree restoration potential. Science 2019, 365, 76–79. [Google Scholar] [CrossRef]
- Palmero-Iniesta, M.; Pino, J.; Pesque, L.; Espelta, J.M. Recent forest area increase in Europe: Expanding and regenerating forests differ in their regional patterns, drivers and productivity trends. Eur. J. For. Res. 2021, 140, 793–805. [Google Scholar] [CrossRef]
- Francini, S.; Chirici, G.; Chiesi, L.; Costa, P.; Caldarelli, G.; Mancuso, S. Global spatial assessment of potential for new peri-urban forests to combat climate change. Nat. Cities 2014, 1, 286–294. [Google Scholar] [CrossRef]
- Veldman, J.W.; Overbeck, G.E.; Negreiros, D.; Mahy, G.; Le Stradic, S.; Fernandes, G.W.; Durigan, G.; Buisson, E.; Putz, F.E.; Bond, W.J. Where Tree Planting and Forest Expansion are Bad for Biodiversity and Ecosystem Services. BioScience 2015, 65, 1011–1018. [Google Scholar] [CrossRef]
- Payn, T.; Carnus, J.-M.; Freer-Smith, P.; Kimberley, M.; Kollert, W.; Liu, S.; Orazio, C.; Rodriguez, L.; Silva, L.N.; Wingfield, M.J. Changes in planted forests and future global implications. For. Ecol. Manag. 2015, 352, 57–67. [Google Scholar] [CrossRef]
- Walker, W.S.; Gorelik, S.R.; Cook-Patton, S.C.; Baccini, A.; Farina, M.K.; Solvik, K.K.; Ellis, P.W.; Sanderman, J.; Houghton, R.A.; Leavitt, S.M.; et al. The global potential for increased storage of carbon on land. Proc. Natl. Acad. Sci. USA 2022, 119, e2111312119. [Google Scholar] [CrossRef]
- Mo, L.; Zohner, C.M.; Reich, P.B.; Liang, J.; de Miguel, S.; Nabuurs, G.-J.; Renner, S.S.; van den Hoogen, J.; Araza, A.; Herold, M.; et al. Integrated global assessment of the natural forest carbon potential. Nature 2013, 624, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Bazzaz, F.A. The physiological ecology of plant succession. Annu. Rev. Ecol. Syst. 1979, 10, 351–371. [Google Scholar] [CrossRef]
- Büntgen, U.; Krusic, P.J.; Piermattei, A.; Coomes, D.A.; Esper, J.; Myglan, V.S.; Kirdyanov, A.V.; Camarero, J.J.; Crivellaro, A.; Körner, C. Limited capacity of tree growth to mitigate the global greenhouse effect under predicted warming. Nat. Commun. 2019, 10, 2171. [Google Scholar] [CrossRef]
- Wasternak, C. A plant’s balance of growth and defense—Revisited. New Phytol. 2017, 215, 1291–1294. [Google Scholar] [CrossRef]
- He, Z.; Webster, S.; He, S.Y. Growth–defense trade-offs in plants. Curr. Biol. 2022, 32, R634–R639. [Google Scholar] [CrossRef]
- Issartel, J.; Coiffard, C. Extreme longevity in trees: Live slow, die old? Oecologia 2011, 165, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Brienen, R.J.W.; Fan, C.; Hao, M.; Zhao, X.; Zhang, C. Tree Lifespans in a Warming World: Unravelling the Universal Trade-Off Between Growth and Lifespan in Temperate Forests. Glob. Change Biol. 2025, 31, e70023. [Google Scholar] [CrossRef]
- Bonan, G.B. Forests and Climate Change: Forcings, Feedbacks, and the Climate Benefits of Forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef]
- Winbourne, J.B.; Jones, T.S.; Garvey, S.M.; Harrison, J.L.; Wang, L.; Li, D.; Templer, P.H.; Hutyra, L.R. Tree Transpiration and Urban Temperatures: Current Understanding; Implications, and Future Research Directions. BioScience 2020, 70, 576–588. [Google Scholar] [CrossRef]
- Matteucci, G.; Vanclay, J.; Martin-Vide, J. Do forests areas influence rainfall regime? In Water for Forest and People in the Mediterranean Region—A Challenging Balance; Birot, Y., Gracia, C., Palahí, M., Eds.; What Science Can Tell Us 1; European Forest Institute: Joensuu, Finland, 2011; pp. 32–36. ISBN 978-952-5453-80-5. [Google Scholar]
- Verheyen, K.; Gillerot, L.; Blondeel, H.; De Frenne, P.; De Pauw, K.; Depauw, L.; Lorer, E.; Sanczuk, P.; Schreel, J.; Vanneste, T.; et al. Forest canopies as nature-based solutions to mitigate global change effects on people and nature. J. Ecol. 2024, 112, 2451–2461. [Google Scholar] [CrossRef]
- De Frenne, P.; Graae, B.J.; Brunet, J.; Shevtsova, A.; De Schrijver, A.; Chabrerie, O.; Cousins, S.A.O.; Decocq, G.; Diekmann, M.; Hermy, M.; et al. The response of forest plant regeneration to temperature variation along a latitudinal gradient. Ann. Bot. 2012, 109, 1037–1046. [Google Scholar] [CrossRef]
- Lintunen, J.; Rautiainen, A.; Uusivuori, J. Which Is more Important, Carbon or Albedo? Optimizing Harvest Rotations for Timber and Climate Benefits in a Changing Climate. Am. J. Agric. Econ. 2022, 104, 134–160. [Google Scholar] [CrossRef]
- Hasler, N.; Williams, C.A.; Denney, V.C.; Ellis, P.W.; Shrestha, S.; Terasaki Hart, D.E.; Wolff, N.H.; Yeo, S.; Crowther, T.; Werden, L.K.; et al. Accounting for albedo change to identify climate-positive tree cover restoration. Nat. Commun. 2024, 15, 2275. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Staudt, M. BVOCs and global change. Trends Plant Sci. 2010, 15, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Popkin, G. How much can forests fight climate change? Nature 2019, 565, 280–282. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.G.; Bergeron, Y.; Denneler, B.; Berninger, F.; Tardif, J. Response of Forest Trees to Increased Atmospheric CO2. Crit. Rev. Plant Sci. 2007, 26, 265–283. [Google Scholar] [CrossRef]
- Keenan, T.F.; Luo, X.; Stocker, B.D.; De Kauwe, M.G.; Medlyn, B.E.; Prentice, I.C.; Smith, N.G.; Terrer, C.; Wang, H.; Zhang, Y.; et al. A constraint on historic growth in global photosynthesis due to rising CO2. Nat. Clim. Chang. 2023, 13, 1376–1381. [Google Scholar] [CrossRef]
- Bugmann, H.; Bigler, C. Will the CO2 fertilization effect in forests be offset by reduced tree longevity? Oecologia 2011, 165, 533–544. [Google Scholar] [CrossRef]
- Kirschbaum, M.U.F. Direct and Indirect Climate Change Effects on Photosynthesis and Transpiration. Plant Biol. 2004, 6, 242–253. [Google Scholar] [CrossRef]
- Linderholm, H.W. Growing season changes in the last century. Agric. For. Meteorol. 2006, 137, 1–14. [Google Scholar] [CrossRef]
- Duffy, K.A.; Schwalm, C.R.; Arcus, V.L.; Koch, G.W.; Liang, L.L.; Schipper, L.A. How close are we to the temperature tipping point of the terrestrial biosphere? Sci. Adv. 2021, 7, eaay1052. [Google Scholar] [CrossRef] [PubMed]
- Dusenge, M.E.; Duarte, A.G.; Way, D.A. Plant carbon metabolism and climate change: Elevated CO2 and temperature impacts on photosynthesis, photorespiration and respiration. New Phytol. 2019, 221, 32–49. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, A.; Sanders, T.G.M.; Bolte, A.; Bussotti, F.; Dirnböck, T.; Johnson, J.; Peñuelas, J.; Pollastrini, M.; Prescher, A.-K.; Sardans, J.; et al. Responses of forest ecosystems in Europe to decreasing nitrogen deposition. Environ. Pollut. 2019, 244, 980–994. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, Y.; Ju, W.; Chen, J.M.; Ciais, P.; Cescatti, A.; Sardans, J.; Janssens, I.A.; Wu, M.; Berry, J.A.; et al. Recent global decline of CO2 fertilization effects on vegetation photosynthesis. Science 2020, 370, 1295–1300. [Google Scholar] [CrossRef] [PubMed]
- Beedlow, P.A.; Tingey, D.T.; Phillips, D.L.; Hogsett, W.E.; Olszyk, D.M. Rising atmospheric CO2 and carbon sequestration in forests. Front. Ecol. Environ. 2004, 2, 315–322. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, W.; Forzieri, G.; Cescatti, A. Transition from positive to negative indirect CO2 effects on the vegetation carbon uptake. Nat. Commun. 2024, 15, 1500. [Google Scholar] [CrossRef]
- Braun, S.; Thomas, V.F.D.; Quiring, R.; Flückiger, W. Does nitrogen deposition increase forest production? The role of phosphorus. Environ. Pollut. 2010, 158, 2043–2052. [Google Scholar] [CrossRef]
- Jonard, M.; Fürst, A.; Verstraeten, A.; Thimonier, A.; Timmermann, V.; Potočić, N.; Waldner, P.; Benham, S.; Hansen, K.; Merilä, P.; et al. Tree mineral nutrition is deteriorating in Europe. Glob. Change Biol. 2015, 21, 418–430. [Google Scholar] [CrossRef]
- Laffitte, B.; Seyler, B.C.; Wang, W.; Li, P.; Du, J.; Tang, Y. Declining tree growth rates despite increasing water-use efficiency under elevated CO2 reveals a possible global overestimation of CO2 fertilization effect. Heliyon 2022, 8, e11219. [Google Scholar] [CrossRef]
- McDowell, N.G.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J. Compound climate events increase tree drought mortality across European forests. Sci. Total Environ. 2022, 816, 151604. [Google Scholar] [CrossRef] [PubMed]
- FAO. Global Forest Resource Assessment. Terms and Definitions; Paper 118; FAO: Rome, Italy, 2020; 32p. [Google Scholar]
- Seidl, R.; Schelhaas, M.J.; Rammer, W.; Verkerk, P.J. Increasing forest disturbances in Europe and their impact on carbon storage. Nat. Clim. Chang. 2014, 4, 806–810. [Google Scholar] [CrossRef]
- Senf, C.; Buras, A.; Zang, C.S.; Rammig, A.; Seidl, R. Excess forest mortality is consistently linked to drought across Europe. Nat. Commun. 2020, 11, 6200. [Google Scholar] [CrossRef] [PubMed]
- Patacca, M.; Lindner, M.; Lucas-Borja, M.E.; Cordonnier, T.; Fidej, G.; Gardiner, B.; Hauf, Y.; Jasinevičius, G.; Labonne, S.; Linkevičius, E.; et al. Significant increase in natural disturbance impacts on European forests since 1950. Glob. Change Biol. 2023, 29, 1359–1376. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Hicke, J.A.; Fisher, R.A.; Allen, C.D.; Aukema, J.; Bentz, B.; Hood, S.; Lichstein, J.W.; Macalady, A.K.; McDowell, N.; et al. Tree mortality from drought, insects, and their interactions in a changing climate. New Phytol. 2015, 208, 674–683. [Google Scholar] [CrossRef]
- Wessely, J.; Essl, F.; Fiedler, K.; Gattringer, A.; Hülber, B.; Ignateva, O.; Moser, D.; Rammer, W.; Dullinger, W.; Seidl, R. A climate-induced tree species bottleneck for forest management in Europe. Nat. Ecol. Evol. 2024, 8, 1109–1117. [Google Scholar] [CrossRef]
- Streit, K.; Brang, P.; Frei, E.R. The Swiss common garden network: Testing assisted migration of tree species in Europe. Front. For. Glob. Chang. 2024, 7, 1396798. [Google Scholar] [CrossRef]
- Koralewski, T.E.; Wang, H.H.; Grant, W.E.; Byram, T.D. Plants on the move: Assisted migration of forest trees in the face of climate change. For. Ecol. Manag. 2015, 344, 30–37. [Google Scholar] [CrossRef]
- Bantis, F.; Graap, J.; Früchtenicht, E.; Bussotti, F.; Radoglou, K.; Brüggemann, W. Field Performances of Mediterranean Oaks in Replicate Common Gardens for Future Reforestation Under Climate Change in Central and Southern Europe: First Results from a Four-Year Study. Forests 2021, 12, 678. [Google Scholar] [CrossRef]
- MacKenzie, W.H.; Mahony, C.R. An ecological approach to climate change-informed tree species selection for reforestation. For. Ecol. Manag. 2021, 481, 118705. [Google Scholar] [CrossRef]
- Forster, E.J.; Healey, J.R.; Dymond, C.; Styles, D. Commercial afforestation can deliver effective climate change mitigation under multiple decarbonisation pathways. Nat. Commun. 2021, 12, 831. [Google Scholar] [CrossRef] [PubMed]
- Bukoski, J.J.; Cook-Patton, S.C.; Melikov, C.; Ban, H.; Chen, J.L.; Goldman, E.D.; Harris, N.L.; Potts, M.D. Rates and drivers of aboveground carbon accumulation in global monoculture plantation forests. Nat. Commun. 2022, 13, 4206. [Google Scholar] [CrossRef] [PubMed]
- Surya Prabha, A.C.; Velumani, R.; Senthivelu, M.; Pragadeesh, S. Carbon Sequestration in Plantations and Agriculture Systems: A Review. Environ. Ecol. 2022, 40, 893–898. [Google Scholar]
- Teskey, R.O.; Saveyn, A.; Steppe, K.; McGuire, M.A. Origin, fate and significance of CO2 in tree stems. New Phytol. 2008, 177, 17–32. [Google Scholar] [CrossRef]
- Noormets, A.; Epron, D.; Domec, J.C.; McNulty, S.G.; Fox, T.; Sun, G.; King, J.S. Effects of forest management on productivity and carbon sequestration: A review and hypothesis. For. Ecol. Manag. 2015, 355, 124–140. [Google Scholar] [CrossRef]
- Brang, P.; Spathelf, P.; Bo Larsen, J.; Bauhus, J.; Boncčìna, A.; Chauvin, C.; Drössler, L.; García-Güemes, C.; Heiri, C.; Kerr, G.; et al. Suitability of close-to-nature silviculture for adapting temperate European forests to climate change. Forestry 2014, 87, 492–503. [Google Scholar] [CrossRef]
- Vilà-Cabrera, A.; Coll, L.; Martínez-Vilalta, J.; Retana, J. Forest management for adaptation to climate change in the Mediterranean basin: A synthesis of evidence. For. Ecol. Manag. 2018, 407, 16–22. [Google Scholar] [CrossRef]
- Walentowski, H.; Falk, W.; Mette, T.; Kunz, J.; Bräuning, A.; Meinardus, C.; Zang, C.; Sutcliffe, L.; Leuschner, C. Assessing future suitability of tree species under climate change by multiple methods: A case study in southern Germany. Ann. For. Res. 2017, 60, 101–126. [Google Scholar] [CrossRef]
- Kunz, J.; Löffler, G.; Bauhus, J. Minor European broadleaved tree species are more drought-tolerant than Fagus sylvatica but not more tolerant than Quercus petraea. For. Ecol. Manag. 2018, 414, 15–27. [Google Scholar] [CrossRef]
- Luyssaert, S.; Schulze, E.D.; Börner, A.; Knohl, A.; Hessenmöller, D.; Law, B.E.; Ciais, P.; Grace, J. Old-growth forests as global carbon sinks. Nature 2008, 455, 213–215. [Google Scholar] [CrossRef]
- Knohl, A.; Schulze, E.D.; Kolle, O.; Buchmann, N. Large carbon uptake by an unmanaged 250-year-old deciduous forest in Central Germany. Agric. For. Meteorol. 2003, 118, 151–167. [Google Scholar] [CrossRef]
- Roebroek Caspar, T.J.; Duveiller, G.; Seneviratne, S.I.; Davin, E.L.; Cescatti, A. Releasing global forests from human management: How much more carbon could be stored? Science 2023, 380, 749–753. [Google Scholar] [CrossRef]
- Holl, K.D.; Brancalion, P.H.S. Tree planting is not a simple solution. Science 2020, 368, 580–581. [Google Scholar] [CrossRef] [PubMed]
- Scherer-Lorenzen, M. The functional role of biodiversity in the context of global change. In Forests and Global Change; Coomes, D.D., Burslem, D.F.R.P., Simonson, W.D., Eds.; Cambridge University Press: Cambridge, UK; British Ecological Society: London, UK, 2014; pp. 195–237. [Google Scholar]
- Liang, J.; Crowther, T.W.; Picard, N.; Wiser, S.; Zhou, M.; Alberti, G.; Schulze, E.-D.; McGuire, A.D.; Bozzato, F.; Pretzsch, H.; et al. Positive biodiversity-productivity relationship predominant in global forests. Science 2016, 354, aaf8957. [Google Scholar] [CrossRef]
- Beugnon, R.; Ladouceur, E.; Sünnemann, M.; Cesarz, S.; Eisenhauer, N. Diverse forests are cool: Promoting diverse forests to mitigate carbon emissions and climate change. J. Sustain. Agric. Environ. 2022, 1, 5–8. [Google Scholar] [CrossRef]
- Morin, X.; Toigo, M.; Fahse, L.; Guillemot, J.; Cailleret, M.; Bertrand, R.; Cateau, E.; de Coligny, F.; García-Valdés, R.; Ratcliffe, S.; et al. More species, more trees: The role of tree packing in promoting forest productivity. J. Ecol. 2025, 113, 371–386. [Google Scholar] [CrossRef]
- Di Sacco, A.; Hardwick, K.A.; Blakesley, D.; Brancalion, P.H.S.; Breman, E.; Rebola, L.C.; Chomba, S.; Dixon, K.; Elliott, S.; Ruyonga, G.; et al. Ten golden rules for reforestation to optimize carbon sequestration, biodiversity recovery and livelihood benefits. Glob. Change Biol. 2021, 27, 1328–1348. [Google Scholar] [CrossRef]
- Nuñez, M.A.; Davis, K.T.; Dimarco, R.D.; Peltzer, D.A.; Paritsis, J.; Maxwell, B.D.; Pauchard, A. Should tree invasions be used in treeless ecosystems to mitigate climate change? Front. Ecol. Environ. 2021, 19, 334–341. [Google Scholar] [CrossRef]
- Seddon, N.; Smith, A.; Smith, P.; Key, I.; Chausson, A.; Girardin, C.; House, J.; Srivastava, S.; Turner, B. Getting the message right on nature-based solutions to climate change. Glob. Change Biol. 2012, 27, 1518–1546. [Google Scholar] [CrossRef]
- Brancalion, P.H.S.; Holl, K.D. Guidance for successful tree planting initiatives. J. Appl. Ecol. 2020, 57, 2349–2361. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Area | Plants | Soil | Total | GtC | |
|---|---|---|---|---|---|
| Biome | M Km2 | (GtC) | (GtC) | (GtC) | M Km−2 |
| Tropical forests | 17.6 | 212 | 216 | 428 | 24.32 |
| Temperate forests | 10.4 | 59 | 100 | 159 | 15.29 |
| Boreal forests | 13.7 | 88 | 471 | 559 | 40.8 |
| Tropical savannas | 22.5 | 66 | 264 | 330 | 14.67 |
| Temperate grasslands | 12.5 | 9 | 295 | 304 | 24.32 |
| Deserts and semideserts | 45.5 | 8 | 191 | 199 | 4.37 |
| Tundra | 9.5 | 6 | 121 | 127 | 13.37 |
| Wetlands | 3.5 | 15 | 225 | 240 | 68.57 |
| Croplands | 16 | 3 | 128 | 131 | 8.19 |
| Total | 151.2 | 466 | 2011 | 2477 | 16.38 |
| Objective | Management Options |
|---|---|
| Ad hoc tree plantations | |
| Establishing stable and resilient forests for the future | Consider site compatibilities |
| Select tree species suitable for the future climate | |
| Promote persistence rather than fast growth | |
| Fostering biodiversity | Utilize native species |
| Avoid invasive alien species | |
| Commercial tree plantations | |
| Maximizing the persistence of carbon over time | Promote carbon storage in woody products |
| Managed forest | |
| Increasing carbon storage | Promote the complexity of the aboveground structure |
| Enhance the capacity of soil to stock carbon | |
| Promote carbon storage in woody products | |
| Fostering biodiversity | Close-to-Nature Silviculture |
| Foster minor and accessory species | |
| Reducing the impacts of forest disturbances | Plan and execute appropriate forestry roads |
| Monitor health conditions | |
| Reducing the impacts of forestry interventions | Reduce the use of fuel in forestry works |
| Unmanaged forests and natural ecosystems | |
| Fostering protection and conservation | Avoid forestation in intact ecosystems |
| Monitoring impacts and health conditions | |
| Conservative management | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bussotti, F.; Pollastrini, M. Planting Trees as a Nature-Based Solution to Mitigate Climate Change: Opportunities, Limits, and Trade-Offs. Forests 2025, 16, 810. https://doi.org/10.3390/f16050810
Bussotti F, Pollastrini M. Planting Trees as a Nature-Based Solution to Mitigate Climate Change: Opportunities, Limits, and Trade-Offs. Forests. 2025; 16(5):810. https://doi.org/10.3390/f16050810
Chicago/Turabian StyleBussotti, Filippo, and Martina Pollastrini. 2025. "Planting Trees as a Nature-Based Solution to Mitigate Climate Change: Opportunities, Limits, and Trade-Offs" Forests 16, no. 5: 810. https://doi.org/10.3390/f16050810
APA StyleBussotti, F., & Pollastrini, M. (2025). Planting Trees as a Nature-Based Solution to Mitigate Climate Change: Opportunities, Limits, and Trade-Offs. Forests, 16(5), 810. https://doi.org/10.3390/f16050810