
Forest Soil Fungal Diversity in Stands of Norway Spruce (Picea abies (L.) Karst.) of Different Ages

 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites and Sampling
2.2. Environmental Variables
2.3. DNA Extraction and High-Throughput Sequencing
2.4. Bioinformatics and Statistical Analyses
3. Results
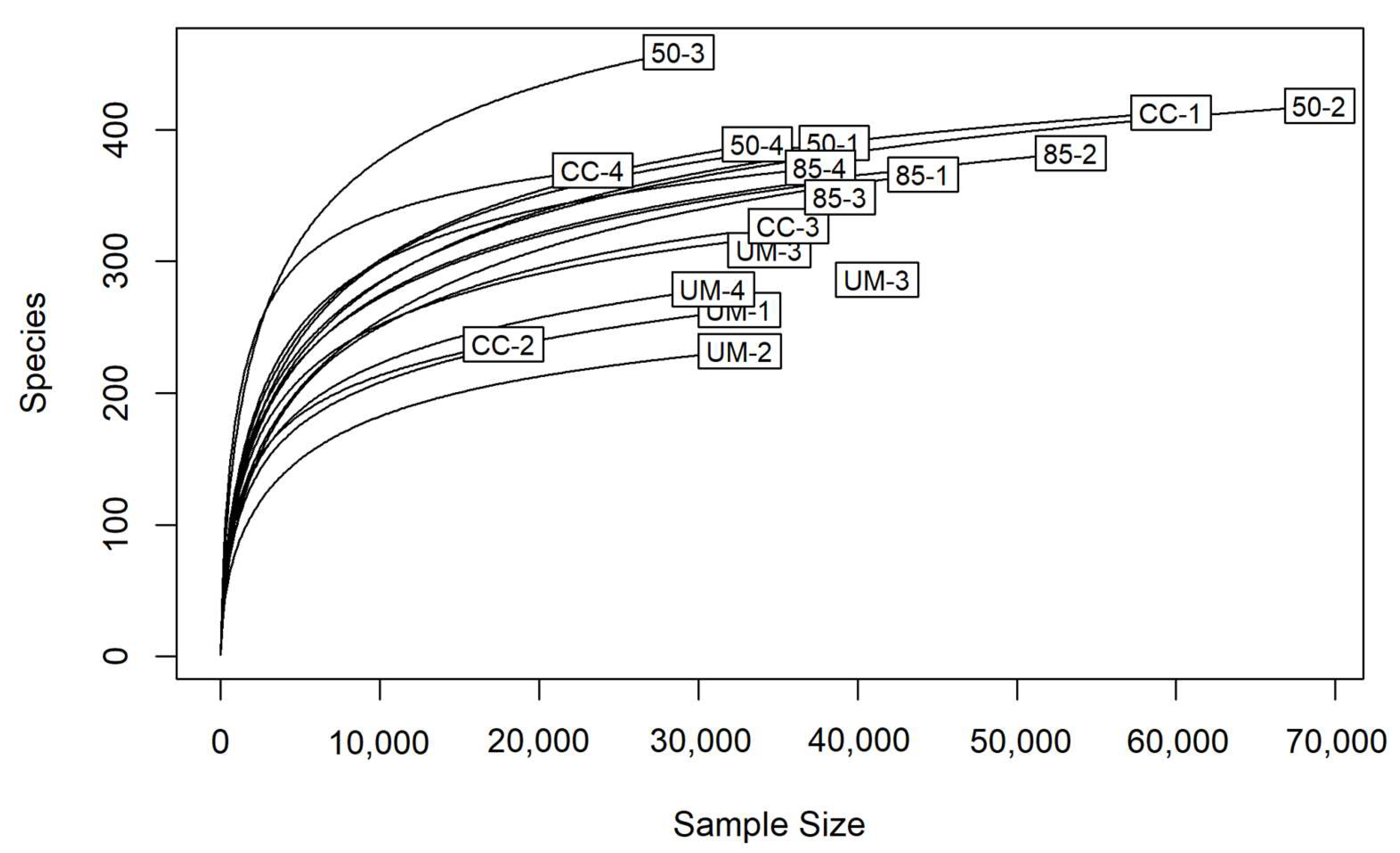
3.1. Sequence Analysis
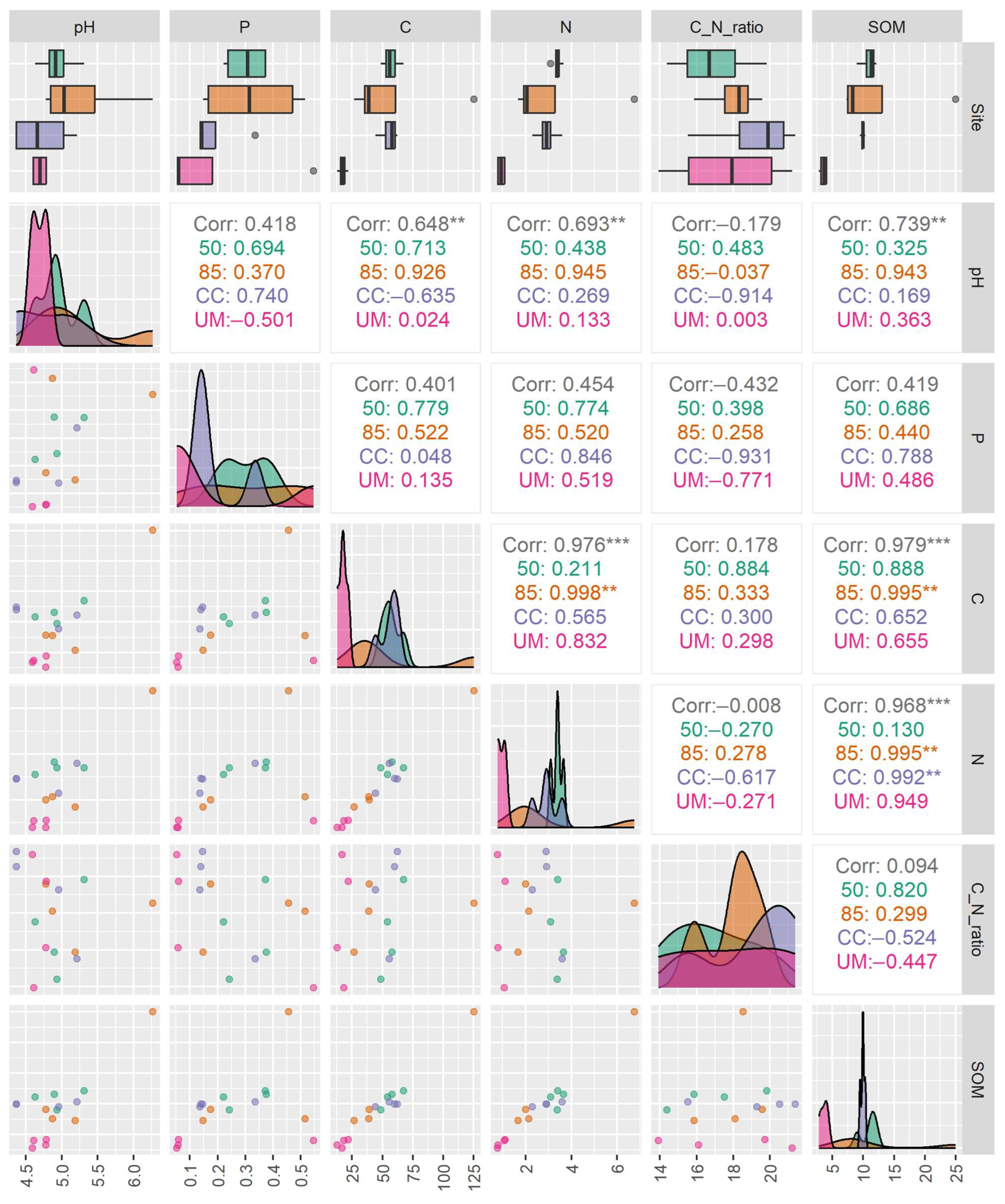
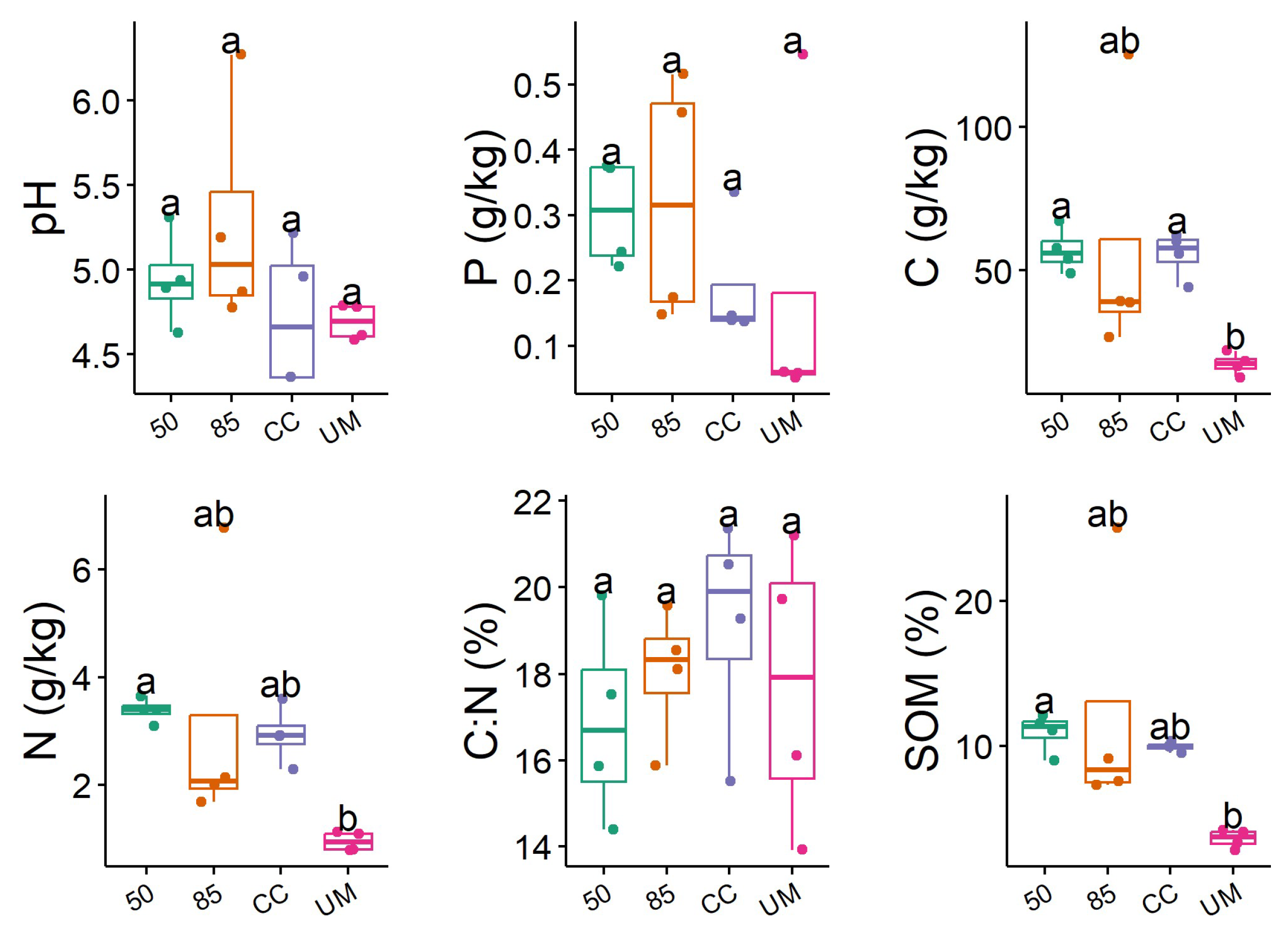
3.2. Soil Chemical Characteristics
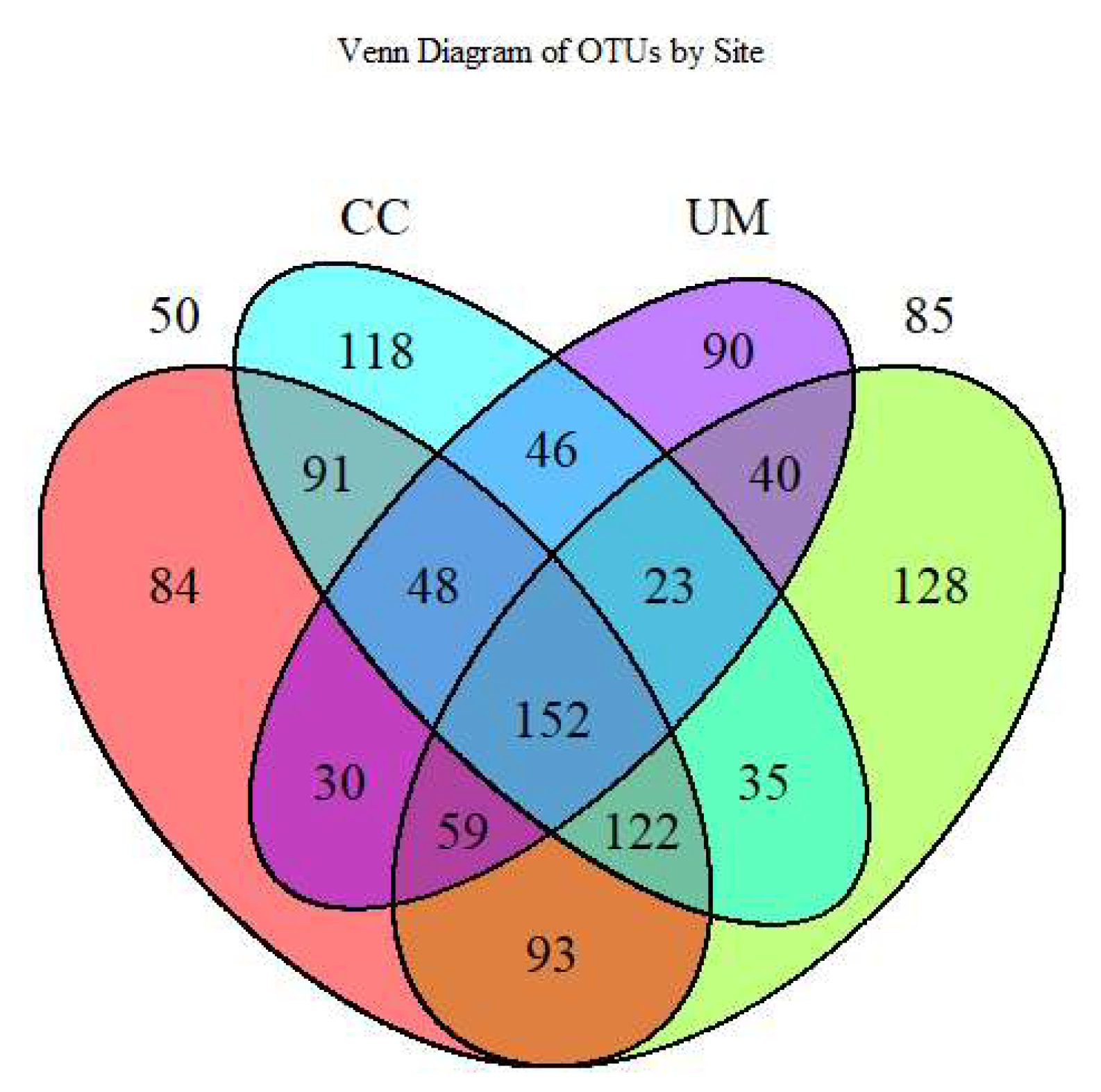
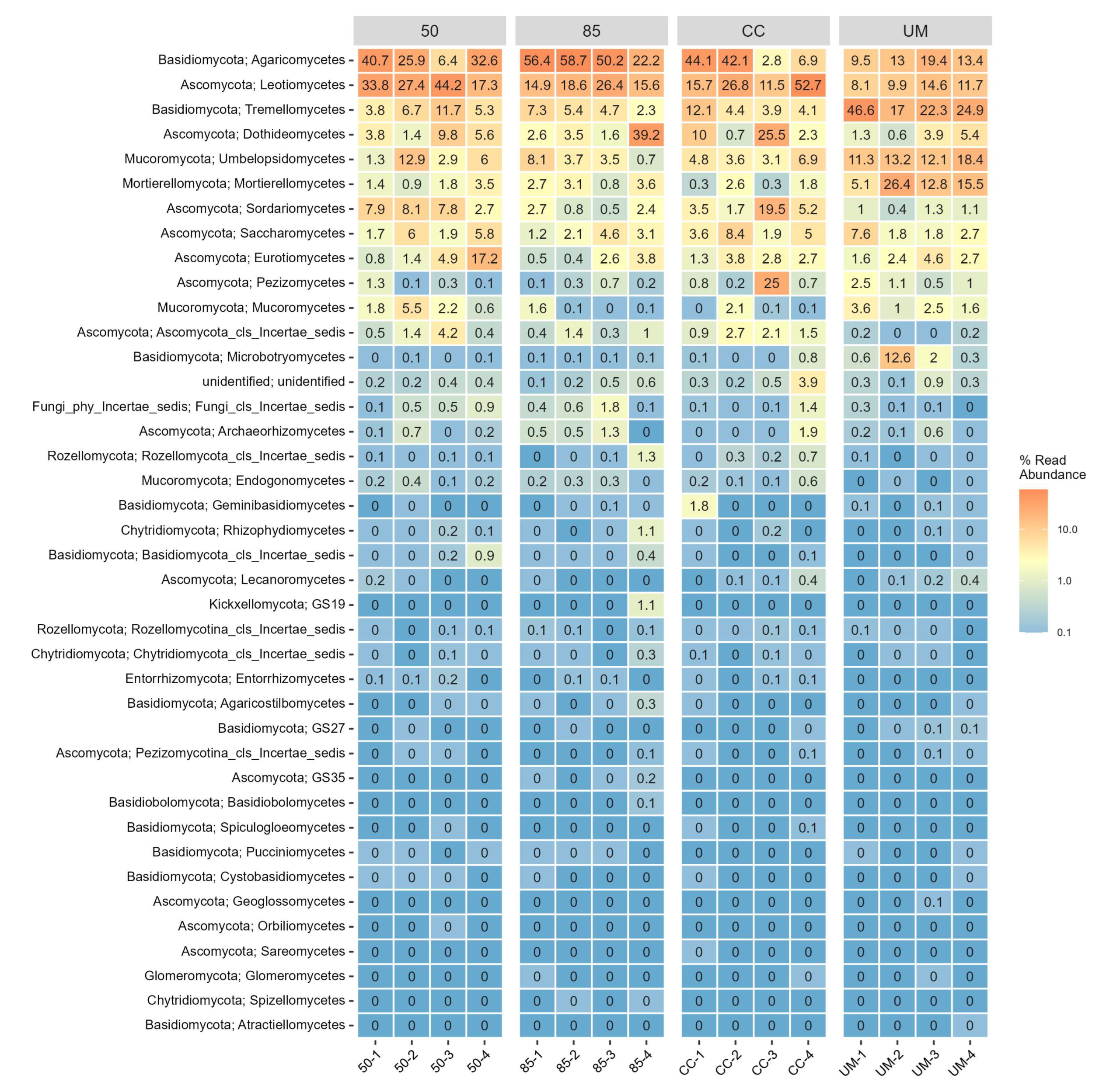
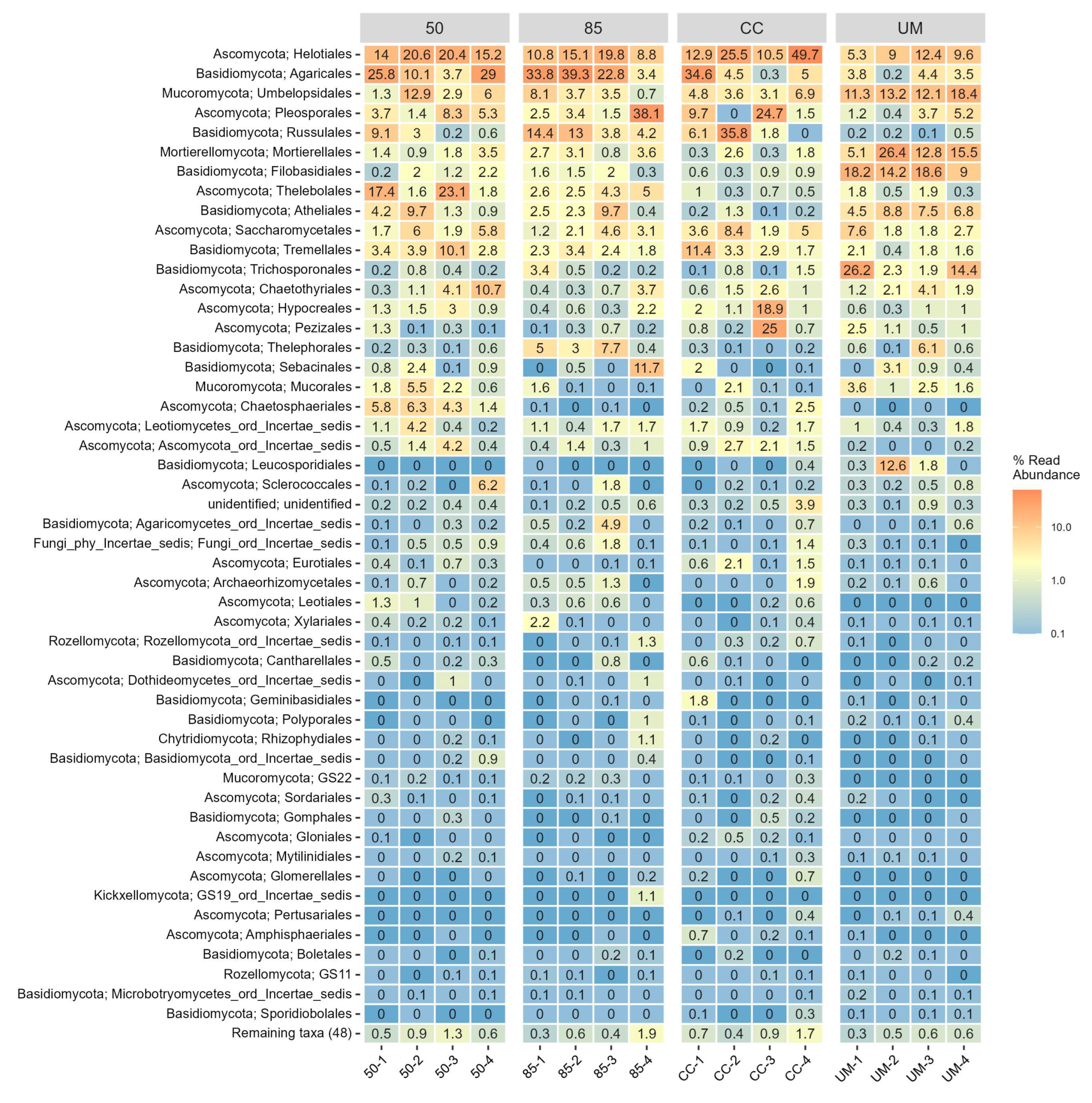
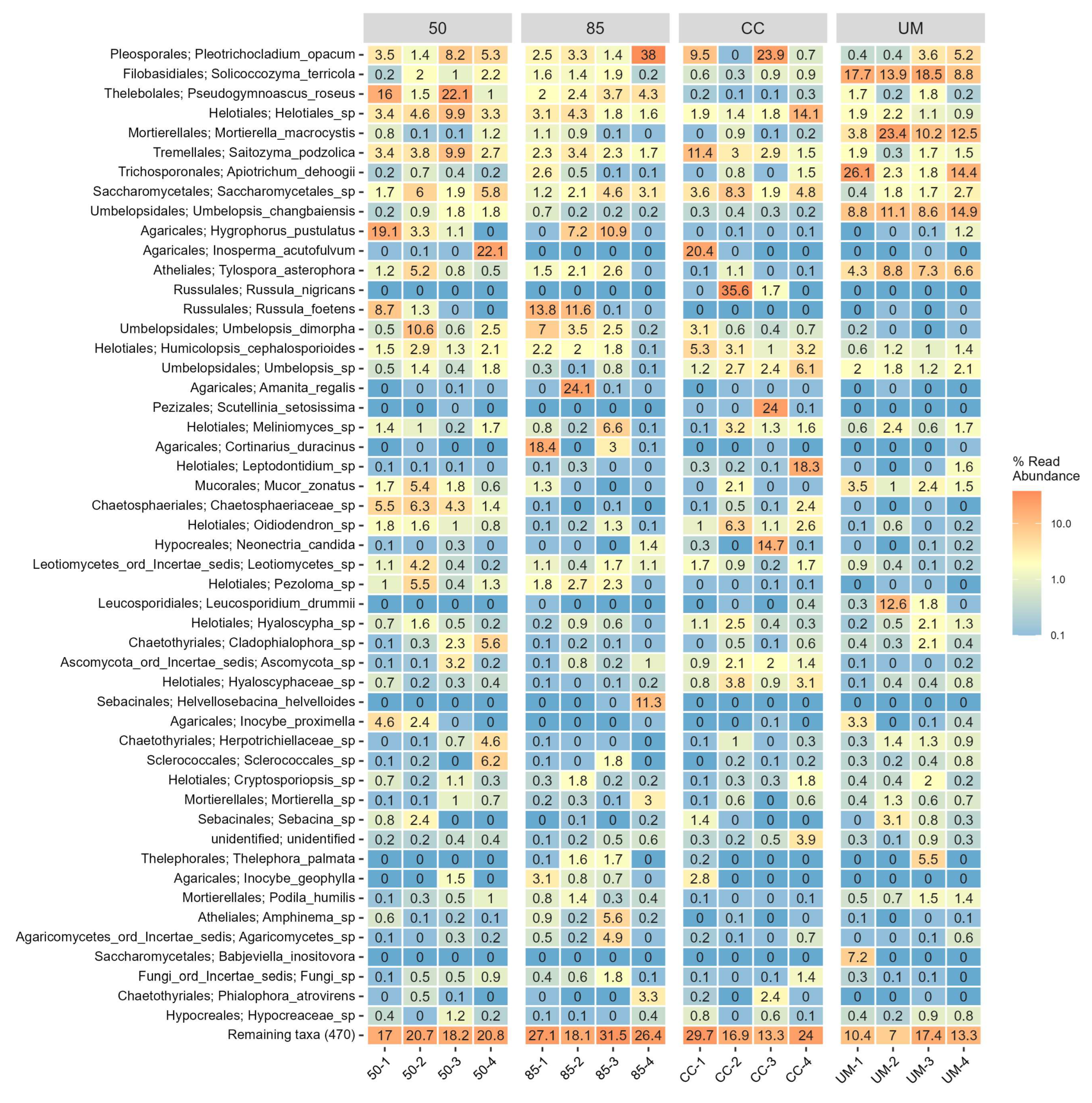
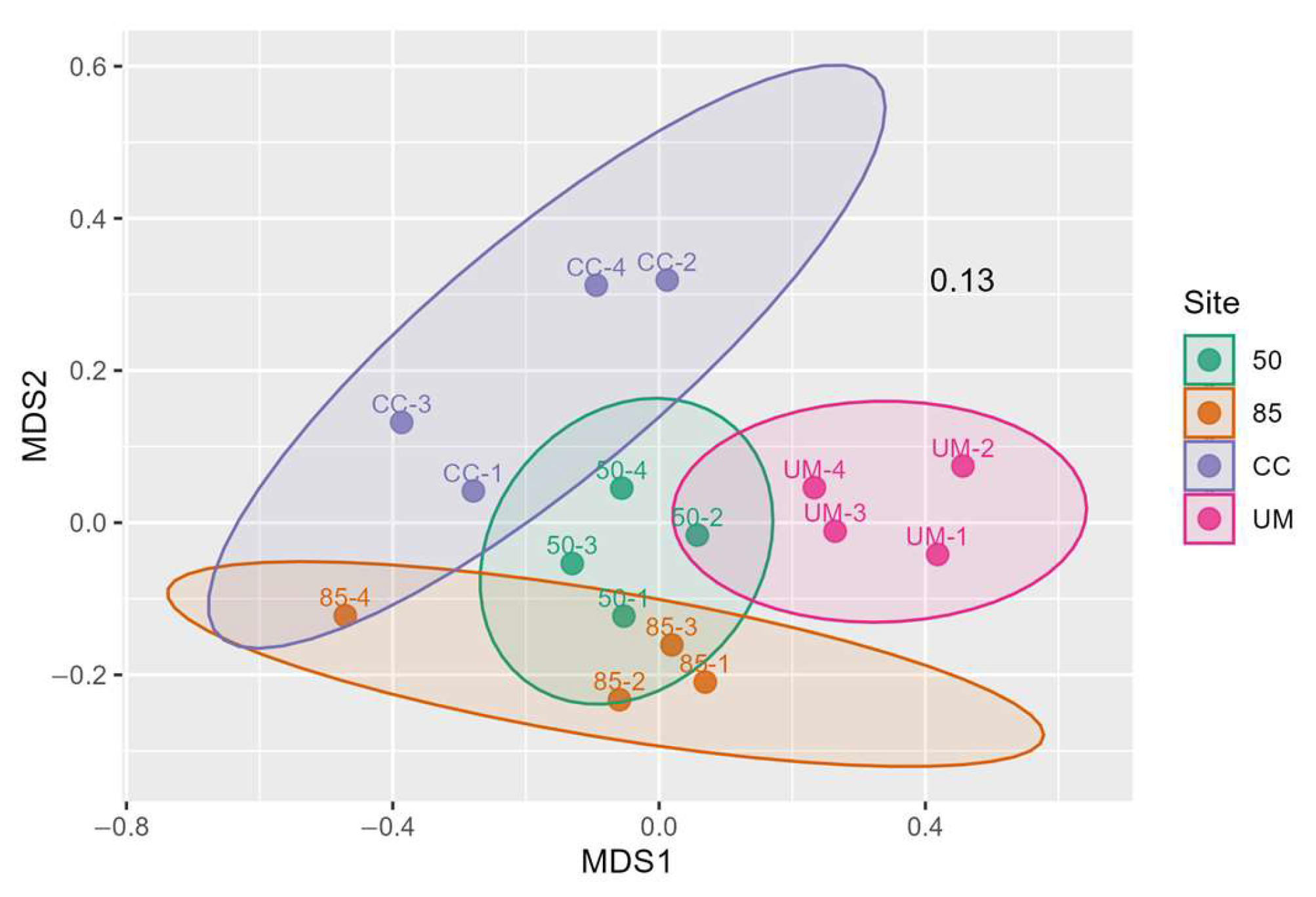
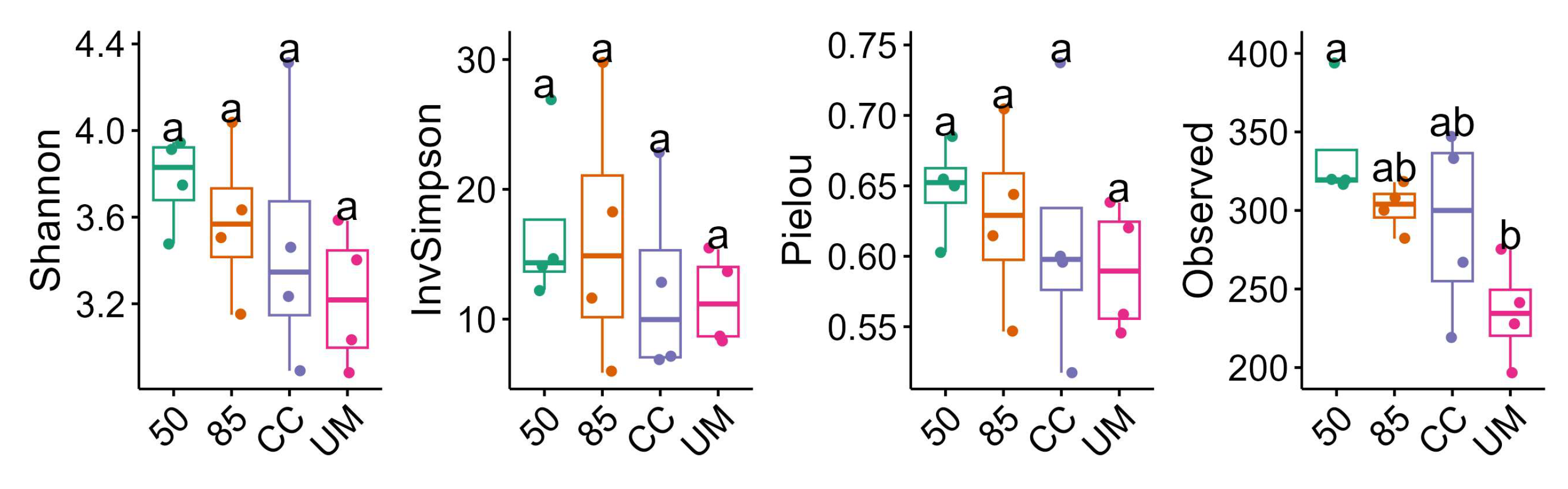
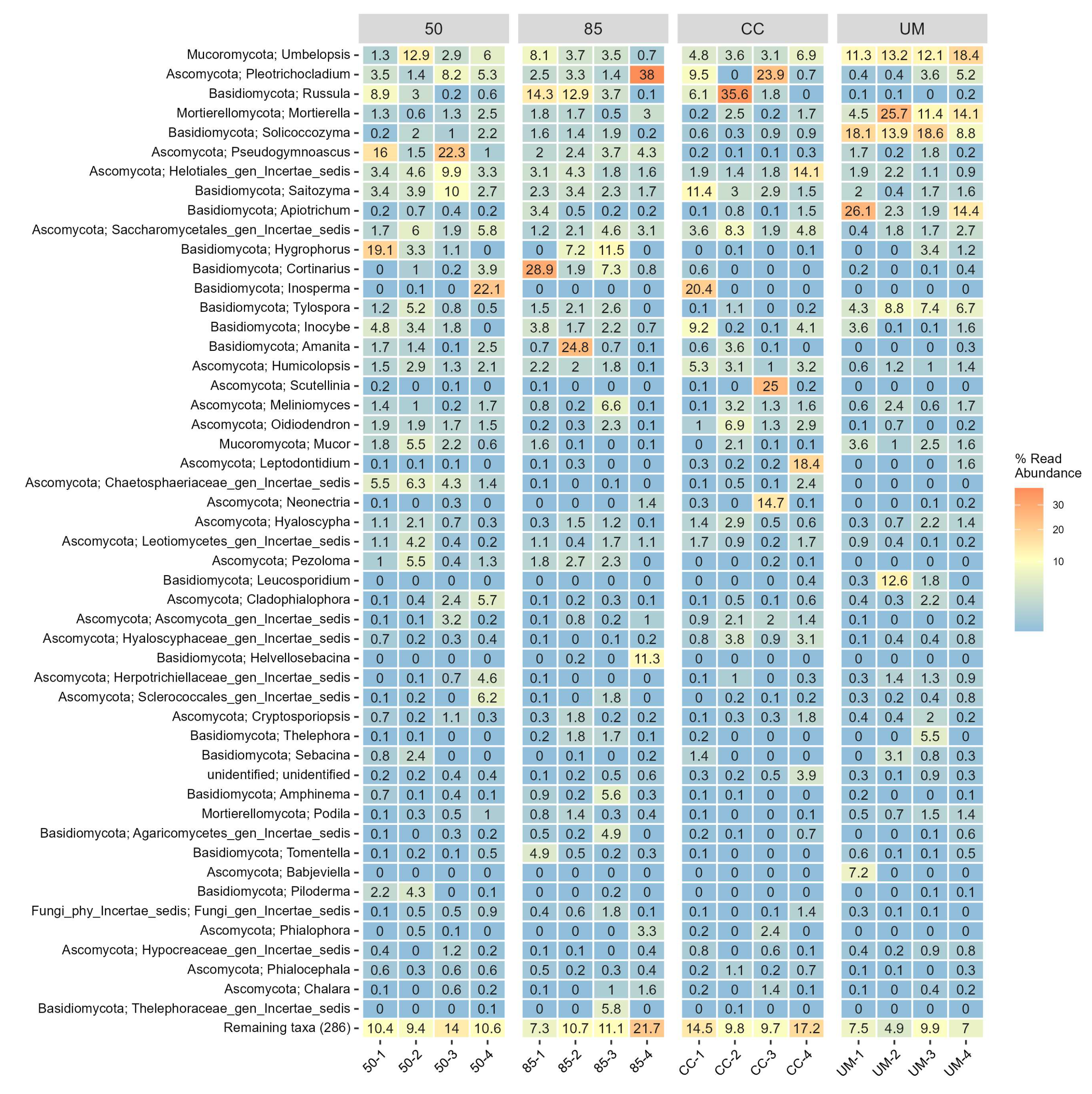
3.3. Taxonomic Assignments and Diversity of Fungal Communities
3.4. Relationships Between Fungal Communities and Environmental Variables
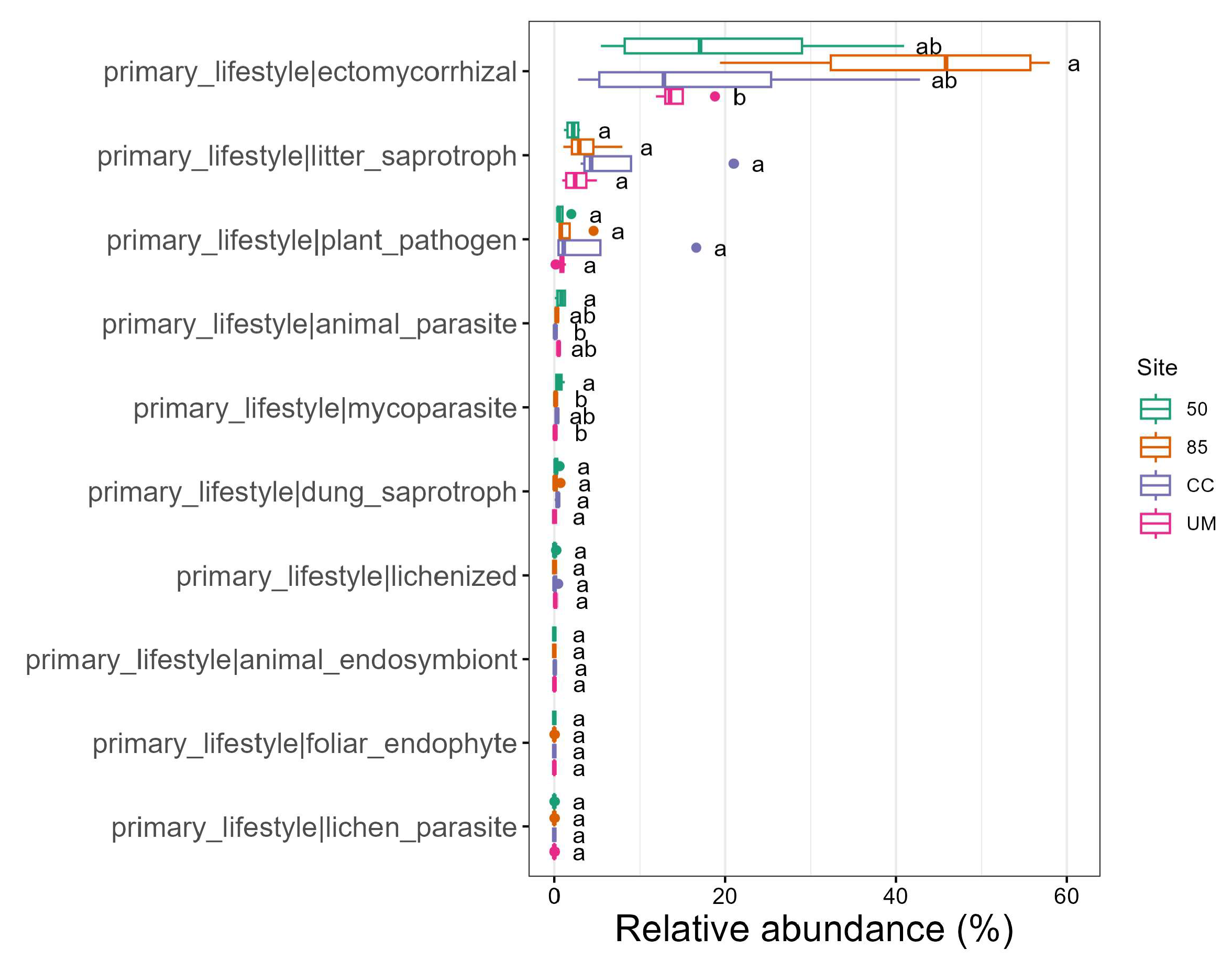
3.5. Ecological Traits of Fungi
4. Discussion
4.1. Diversity of Fungal Communities
4.2. The Influence of Forest Management
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A
References
- Dighton, J. Fungi in Ecosystem Processes; CRC Press: Boca Raton, FL, USA, 2018; p. 434. [Google Scholar]
- Li, X.; Qu, Z.; Zhang, Y.; Ge, Y.; Sun, H. Soil fungal community and potential function in different forest ecosystems. Diversity 2022, 14, 520. [Google Scholar] [CrossRef]
- Heilmann-Clausen, J.; Barron, E.S.; Boddy, L.; Dahlberg, A.; Griffith, G.W.; Nordén, J.; Ovaskainen, O.; Perini, C.; Senn-Irlet, B.; Halme, P. A fungal perspective on conservation biology. Conserv. Biol. 2015, 29, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Bowd, E.J.; Banks, S.C.; Bissett, A.; May, T.W.; Lindenmayer, D.B. Disturbance alters the forest soil microbiome. Mol. Ecol. 2022, 31, 419–447. [Google Scholar] [CrossRef]
- Maron, J.L.; Marler, M.; Klironomos, J.N.; Cleveland, C.C. Soil fungal pathogens and the relationship between plant diversity and productivity. Ecol. Lett. 2011, 14, 36–41. [Google Scholar] [CrossRef]
- Frąc, M.; Hannula, S.E.; Bełka, M.; Jędryczka, M. Fungal biodiversity and their role in soil health. Front. Microbiol. 2018, 9, 707. [Google Scholar] [CrossRef] [PubMed]
- López-Bucio, J.; Pelagio-Flores, R.; Herrera-Estrell, A. Trichoderma as biostimulant: Exploiting the multilevel properties of a plant beneficial fungus. Sci. Hortic. 2015, 196, 109–123. [Google Scholar] [CrossRef]
- Rouphael, Y.; Franken, P.; Schneider, C.; Schwarz, D.; Giovannetti, M.; Agnolucci, M.; De Pascale, S.; Bonini, P.; Colla, G. Arbuscular mycorrhizal fungi act as biostimulants in horticultural crops. Sci. Hortic. 2015, 196, 91–108. [Google Scholar] [CrossRef]
- Shi, L.; Dossa, G.G.; Paudel, E.; Zang, H.; Xu, J.; Harrison, R.D. Changes in fungal communities across a forest disturbance gradient. Appl. Environ. Microbiol. 2019, 85, e00080-19. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Lücking, R. Fungal Diversity Revisited: 2.2 to 3.8 Million Species. Microbiol. Spectr. 2017, 5, 10.1128. [Google Scholar] [CrossRef]
- Goldmann, K.; Schöning, I.; Buscot, F.; Wubet, T. Forest management type influences diversity and community composition of soil fungi across temperate forest ecosystems. Front. Microbiol. 2015, 6, 1300. [Google Scholar] [CrossRef]
- Paillet, Y.; Berges, L.; Hjalten, J.; Odor, P.; Avon, C.; Bernhardt-Romermann, M.; Bijlsma, R.J.; De Bruyn, L.U.C.; Fuhr, M.; Grandin, U.L.F.; et al. Biodiversity differences between managed and unmanaged forests: Meta-analysis of species richness in Europe. Conserv. Biol. 2010, 24, 101–112. [Google Scholar] [CrossRef]
- Foote, J.A.; Boutton, T.W.; Scott, D.A. Soil C and N storage and microbial biomass in US southern pine forests: Influence of forest management. For. Ecol. Manag. 2015, 355, 48–57. [Google Scholar] [CrossRef]
- James, J.; Harrison, R. The effect of harvest on forest soil carbon: A meta-analysis. Forests 2016, 7, 308. [Google Scholar] [CrossRef]
- Ramirez, K.S.; Craine, J.M.; Fierer, N. Consistent effects of nitrogen amendments on soil microbial communities and processes across biomes. Glob. Chang. Biol. 2012, 18, 1918–1927. [Google Scholar] [CrossRef]
- Colombo, F.; Macdonald, C.A.; Jeffries, T.C.; Powell, J.R.; Singh, B.K. Impact of forest management practices on soil bacterial diversity and consequences for soil processes. Soil. Biol. Biochem. 2016, 94, 200–210. [Google Scholar] [CrossRef]
- Chen, J.; Chazdon, R.L.; Swenson, N.G.; Xu, H.; Luo, T. Drivers of soil microbial community assembly during recovery from selective logging and clear-cutting. J. Appl. Ecol. 2021, 58, 2231–2242. [Google Scholar] [CrossRef]
- Sun, S.; Li, S.; Avera, B.N.; Strahm, B.D.; Badgley, B.D. Soil bacterial and fungal communities show distinct recovery patterns during forest ecosystem restoration. Appl. Environ. Microbiol. 2017, 83, e00966-17. [Google Scholar] [CrossRef]
- Baldrian, P. The known and the unknown in soil microbial ecology. FEMS Microbiol. Ecol. 2019, 95, fiz005. [Google Scholar] [CrossRef]
- Küffer, N.; Senn-Irlet, B. Influence of forest management on the species richness and composition of wood-inhabiting basidiomycetes in Swiss forests. Biodivers. Conserv. 2005, 14, 2419–2435. [Google Scholar] [CrossRef]
- Brukas, V.; Weber, N. Forest management after the economic transition—At the crossroads between German and Scandinavian traditions. For. Policy Econ. 2009, 11, 586–592. [Google Scholar] [CrossRef]
- Brukas, V.; Felton, A.; Lindbladh, M.; Sallnäs, O. Linking forest management, policy and biodiversity indicators–A comparison of Lithuania and Southern Sweden. For. Ecol. Manag. 2013, 291, 181–189. [Google Scholar] [CrossRef]
- Levers, C.; Verkerk, P.J.; Müller, D.; Verburg, P.H.; Butsic, V.; Leitão, P.J.; Lindner, M.; Kuemmerle, T. Drivers of forest harvesting intensity patterns in Europe. For. Ecol. Manag. 2014, 315, 160–172. [Google Scholar] [CrossRef]
- Tērauds, A.; Brūmelis, G.; Nikodemus, O. Seventy-year changes in tree species composition and tree ages in state-owned forests in Latvia. Scand. J. For. Res. 2011, 26, 446–456. [Google Scholar] [CrossRef]
- Lim, Y.W.; Kim, B.K.; Kim, C.; Jung, H.S.; Kim, B.S.; Lee, J.H.; Chun, J. Assessment of soil fungal communities using pyrosequencing. J. Microbiol. 2010, 48, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Allison, S.D.; Treseder, K.K. Climate change feedbacks to microbial decomposition in boreal soils. Fungal Ecol. 2011, 4, 362–374. [Google Scholar] [CrossRef]
- Dutta, H.; Dutta, A. The microbial aspect of climate change. Energy Ecol. Environ. 2016, 1, 209–232. [Google Scholar] [CrossRef]
- Shi, L.L.; Mortimer, P.E.; Ferry Slik, J.W.; Zou, X.M.; Xu, J.; Feng, W.T.; Qiao, L. Variation in forest soil fungal diversity along a latitudinal gradient. Fungal Divers. 2014, 64, 305–315. [Google Scholar] [CrossRef]
- Linares, J. Evaluating the Effectiveness of the Internal Transcribed Sequences (ITS) as DNA Barcodes to Estimate Fungal Diversity. Master’s Thesis, Nova Southeastern University, Fort Lauderdale, FL, USA, 2023. [Google Scholar]
- Leckie, S.E. Methods of microbial community profiling and their application to forest soils. For. Ecol. Manag. 2005, 220, 88–106. [Google Scholar] [CrossRef]
- The Climate of Latvia. Latvian Environment, Geology and Meteorology Centre. 2024. Available online: https://videscentrs.lvgmc.lv/lapas/latvijas-klimats (accessed on 6 December 2024).
- National Forest Inventory. Latvian State Forest Research Institute Silava. 2023. Available online: https://www.silava.lv/petnieciba/nacionalais-meza-monitorings (accessed on 15 December 2024).
- State Forest Register. State Forest Service. 2024. Available online: https://data.gov.lv/dati/eng/dataset/meza-valsts-registra-meza-dati (accessed on 3 December 2024).
- Bušs, K. Forest ecosystem classification in Latvia. Proc. Latv. Acad. Sci. Sect. B Nat. Exact. Appl. Sci. 1997, 51, 204–218. [Google Scholar]
- Anslan, S.; Bahram, M.; Hiiesalu, I.; Tedersoo, L. PipeCraft: Flexible open-source toolkit for bioinformatics analysis of custom high-throughput amplicon sequencing data. Mol. Ecol. Resour. 2017, 17, e234–e240. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Bengtsson-Palme, J.; Ryberg, M.; Hartmann, M.; Branco, S.; Wang, Z.; Godhe, A.; De Wit, P.; Sanchez-Garcia, M.; Ebersberger, I.; Dousa, F.; et al. Improved software detection and extraction of ITS1 and ITS2 from ribosomal ITS sequences of fungi and other eukaryotes for analysis of environmental sequencing data. Methods Ecol. Evol. 2013, 4, 914–919. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Abarenkov, K.; Zirk, A.; Piirmann, T.; Pöhönen, R.; Ivanov, F.; Nilsson, R.H.; Kõljalg, U. UNITE General FASTA Release for Fungi; UNITE Community: London, UK, 2024. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024; Available online: http://www.R-project.org/ (accessed on 3 December 2024).
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.P.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Package ‘Vegan’. Community Ecology Package, Version, 2(9). Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 3 December 2024).
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Cui, Y.; Li, X.; Yao, M. Microeco: An R package for data mining in microbial community ecology. FEMS Microbiol. Ecol. 2020, 97, fiaa255. [Google Scholar] [CrossRef]
- Andersen, K.S.; Kirkegaard, R.H.; Karst, S.M.; Albertsen, M. Ampvis2: An R package to analyse and visualise 16S rRNA amplicon data. bioRxiv 2018. [Google Scholar] [CrossRef]
- Põlme, S.; Abarenkov, K.; Nilsson, R.H.; Lindahl, B.D.; Clemmensen, K.E.; Kauserud, H.; Nguyen, N.; Kjøslev, G.; Adojaa, K.; Vizzini, A.; et al. FungalTraits: A User-Friendly Traits Database of Fungi and Fungus-Like Stramenopiles. Fungal Divers. 2021, 105, 1–16. [Google Scholar] [CrossRef]
- Rähn, E.; Tedersoo, L.; Adamson, K.; Drenkhan, T.; Sibul, I.; Lutter, R.; Anslan, S.; Pritsch, K.; Drenkhan, R. Rapid shift of soil fungal community compositions after clear-cutting in hemiboreal coniferous forests. For. Ecol. Manag. 2023, 544, 121211. [Google Scholar] [CrossRef]
- Kujawska, M.B.; Rudawska, M.; Wilgan, R.; Leski, T. Similarities and differences among soil fungal assemblages in managed forests and formerly managed forest reserves. Forests 2021, 12, 353. [Google Scholar] [CrossRef]
- Hui, N.; Jumpponen, A.; Niskanen, T.; Liimatainen, K.; Jones, K.L.; Koivula, T.; Romantschuk, M.; Strömmer, R. EcM fungal community structure, but not diversity, altered in a Pb-contaminated shooting range in a boreal coniferous forest site in Southern Finland. FEMS Microbiol. Ecol. 2011, 76, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Klavina, D.; Tedersoo, L.; Agan, A.; Adamson, K.; Bitenieks, K.; Gaitnieks, T.; Drenkhan, R. Soil fungal communities in young Norway spruce-dominant stands: Footprints of former land use and selective thinning. Eur. J. For. Res. 2022, 141, 503–516. [Google Scholar] [CrossRef]
- Ampoorter, E.; Barbaro, L.; Jactel, H.; Baeten, L.; Boberg, J.; Carnol, M.; Castagneyrol, B.; Charbonnier, Y.; Dawud, S.M.; Deconchat, M.; et al. Tree diversity is key for promoting the diversity and abundance of forest-associated taxa in Europe. Oikos 2020, 129, 133–146. [Google Scholar] [CrossRef]
- Tedorsoo, L.; Anslan, S.; Bahram, M.; Drenkhan, R.; Pritsch, K.; Buegger, F.; Padari, A.; Hagh-Doust, N.; Mikryukov, V.; Gohar, D.; et al. Regional-Scale In-Depth Analysis of Soil Fungal Diversity Reveals Strong pH and Plant Species Effects in Northern Europe. Front. Microbiol. 2020, 11, 1953. [Google Scholar] [CrossRef]
- Sterkenburg, E.; Bahr, A.; Brandström Durling, M.; Clemmensen, K.E.; Lindahl, B.D. Changes in fungal commu-nities along a boreal forest soil fertility gradient. New Phytol. 2015, 207, 1145–1158. [Google Scholar] [CrossRef]
- Shade, A. Diversity is the question, not the answer. ISME J. 2017, 11, 1–6. [Google Scholar] [CrossRef]
- Bittleston, L. Connecting microbial community assembly and function. COMIRC 2024, 80, 10212. [Google Scholar] [CrossRef] [PubMed]
- Tomao, A.; Bonet, J.A.; Castano, C.; de-Miguel, S. How does forest management affect fungal diversity and community composition? Current knowledge and future perspectives for the conservation of forest fungi. For. Ecol. Manag. 2020, 457, 117678. [Google Scholar] [CrossRef]
- Wang, T.; Xu, Q.; Gao, D.; Zhang, B.; Zuo, H.; Jiang, J. Effects of thinning and understory removal on the soil water-holding capacity in Pinus massoniana plantations. Sci. Rep. 2021, 11, 13029. [Google Scholar] [CrossRef]
- Kara, Ö.; Bolat, İ. Influence of soil compaction on microfungal community structure in two soil types in Bartin Province, Turkey. J. Basic Microbiol. 2007, 47, 394–399. [Google Scholar] [CrossRef]
- Suz, L.M.; Barsoum, N.; Benham, S.; Cheffings, C.; Cox, F.; Hackett, L.; Jones, A.G.; Mueller, G.M.; Orme, D.; Seidling, W.; et al. Monitoring ectomycorrhizal fungi at large scales for science, forest management, fungal conservation and environmental policy. Ann. For. Sci. 2015, 72, 877–885. [Google Scholar] [CrossRef]
- Bååth, E. Soil fungal biomass after clear-cutting of a pine forest in central Sweden. Soil. Biol. Biochem. 1980, 12, 495–500. [Google Scholar] [CrossRef]
- Parladé, J.; Queralt, M.; Pera, J.; Bonet, J.A.; Castaño, C.; Martínez-Peña, F.; Piñol, J.; Senar, M.A.; De Miguel, A.M. Temporal dynamics of soil fungal communities after partial and total clear-cutting in a managed Pinus sylvestris stand. For. Ecol. Manag. 2019, 449, 117456. [Google Scholar] [CrossRef]
- Vannette, R.L.; Leopold, D.R.; Fukami, T. Forest area and connectivity influence root-associated fungal communities in a fragmented landscape. Ecology 2016, 97, 2374–2383. [Google Scholar] [CrossRef] [PubMed]
- Grilli, G.; Longo, S.; Huais, P.Y.; Pereyra, M.; Verga, E.G.; Urcelay, C.; Galetto, L. Fungal diversity at fragmented landscapes: Synthesis and future perspectives. Curr. Opin. Microbiol. 2017, 37, 161–165. [Google Scholar] [CrossRef]
- Boeraeve, M.; Honnay, O.; Jacquemyn, H. Effects of host species, environmental filtering and forest age on community assembly of ectomycorrhizal fungi in fragmented forests. Fungal Ecol. 2018, 36, 89–98. [Google Scholar] [CrossRef]
- Boeraeve, C.E.; Pierroz, G.; Guzman, A.; Anderegg, L.D.; Gao, C.; Coleman-Derr, D.; Taylor, J.W.; Bruns, T.D.; Dawson, T.E. Keep your friends close: Host compartmentalisation of microbial communities facilitates decoupling from effects of habitat fragmentation. Ecol. Lett. 2021, 24, 2674–2686. [Google Scholar] [CrossRef]
- Nordén, J.; Åström, J.; Josefsson, T.; Blumentrath, S.; Ovaskainen, O.; Sverdrup-Thygeson, A.; Nordén, B. At which spatial and temporal scales can fungi indicate habitat connectivity? Ecol. Indic. 2018, 91, 138–148. [Google Scholar] [CrossRef]
- Gundersen, P.; Bezemer, Т.M.; Rojas, S.K.; Tedersoo, L.; Vesterdal, L.; Schmidt, I.K. Silva Nova—Restoring soil biology and soil functions to gain multiple benefits in new forests. RIO 2023, 9, e101455. [Google Scholar] [CrossRef]
- Gomes, S.I.F.; Gundersen, P.; Bezemer, T.M.; Barsotti, D.; D’Imperio, L.; Georgopoulos, K.; Justesen, M.J.; Rheault, K.; Rosas, Y.M.; Schmidt, I.K.; et al. Soil Microbiome Inoculation for Resilient and Multifunctional New Forests in Post-Agricultural Landscapes. Glob. Change Biol. 2025, 31, e70031. [Google Scholar] [CrossRef] [PubMed]
- Baldrian, P.; López-Mondéjar, R.; Kohout, P. Forest microbiome and global change. Nat. Rev. Microbiol. 2023, 21, 487–501. [Google Scholar] [CrossRef] [PubMed]
- Eisenhauer, N.; Bender, S.F.; Calderón-Sanou, I.; de Vries, F.T.; Lembrechts, J.J.; Thuiller, W.; Wall, D.H.; Zeiss, R.; Bahram, M.; Beugnon, R.; et al. Frontiers in Soil Ecology—Insights from the World Biodiversity Forum 2022. J. Sustain. Agric. Environ. 2022, 1, 245–261. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Stand Age | Status | Location |
|---|---|---|---|
| Site #1 (50) | 50 | Managed | 57°28′15.2″ N, 22°28′26.6″ E |
| Site #2 (85) | 85 | Managed | 57°28′01.9″ N, 22°27′43.4″ E |
| Site #3 (CC) | 3 | Managed, clear-cut | 57°28′09.7″ N, 22°28′33.1″ E |
| Site #4 (UM) | 58 | Unmanaged | 57°38′26.7″ N, 22°18′28.6″ E |
| Groups | Measure | F | R2 | p-Value | p-Adjusted |
|---|---|---|---|---|---|
| 50 vs. 85 | Bray–Curtis | 1.21 | 0.17 | 0.280 | 0.2800 |
| 50 vs. CC | Bray–Curtis | 1.47 | 0.20 | 0.098 | 0.1176 |
| 50 vs. UM | Bray–Curtis | 4.64 | 0.44 | 0.031 | 0.0720 |
| 85 vs. CC | Bray–Curtis | 1.62 | 0.21 | 0.087 | 0.1176 |
| 85 vs. UM | Bray–Curtis | 4.06 | 0.40 | 0.036 | 0.0720 |
| CC vs. UM | Bray–Curtis | 3.92 | 0.40 | 0.025 | 0.0720 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katrevičs, J.; Bitenieks, K.; Jansons, Ā.; Jansone, B.; Ruņģis, D.E. Forest Soil Fungal Diversity in Stands of Norway Spruce (Picea abies (L.) Karst.) of Different Ages. Forests 2025, 16, 500. https://doi.org/10.3390/f16030500
Katrevičs J, Bitenieks K, Jansons Ā, Jansone B, Ruņģis DE. Forest Soil Fungal Diversity in Stands of Norway Spruce (Picea abies (L.) Karst.) of Different Ages. Forests. 2025; 16(3):500. https://doi.org/10.3390/f16030500
Chicago/Turabian StyleKatrevičs, Juris, Krišs Bitenieks, Āris Jansons, Baiba Jansone, and Dainis Edgars Ruņģis. 2025. "Forest Soil Fungal Diversity in Stands of Norway Spruce (Picea abies (L.) Karst.) of Different Ages" Forests 16, no. 3: 500. https://doi.org/10.3390/f16030500
APA StyleKatrevičs, J., Bitenieks, K., Jansons, Ā., Jansone, B., & Ruņģis, D. E. (2025). Forest Soil Fungal Diversity in Stands of Norway Spruce (Picea abies (L.) Karst.) of Different Ages. Forests, 16(3), 500. https://doi.org/10.3390/f16030500